- Эволюция
-
Биологи́ческая эволю́ция (от лат. evolutio — «развёртывание») — естественный процесс развития живой природы, сопровождающийся изменением генетического состава популяций, формированием адаптаций, видообразованием и вымиранием видов, преобразованием экосистем и биосферы в целом.
Существует несколько эволюционных теорий, объясняющих механизмы, лежащие в основе эволюционных процессов. В данный момент общепринятой является синтетическая теория эволюции (СТЭ), являющаяся синтезом классического дарвинизма и популяционной генетики[1]. СТЭ позволяет объяснить связь материала эволюции (генетические мутации) и механизма эволюции (естественный отбор). В рамках СТЭ эволюция определяется как процесс изменения частот аллелей генов в популяциях организмов в течение времени, превышающего продолжительность жизни одного поколения[2].
Чарльз Дарвин первым сформулировал теорию эволюции путём естественного отбора. Эволюция путём естественного отбора — это процесс, который следует из трёх фактов о популяциях: 1) рождается больше потомства, чем может выжить; 2) у разных организмов разные черты, что приводит к различиям в выживаемости и вероятности оставить потомство; 3) эти черты — наследуемые. Эти условия приводят к появлению внутривидовой конкуренции и избирательной элиминации наименее приспособленных к среде особей, что ведёт к увеличению в следующем поколении доли таких особей, черты которых способствуют выживанию и размножению в этой среде. Естественный отбор — единственная известная причина адаптаций, но не единственная причина эволюции. К числу неадаптивных причин относятся генетический дрейф, поток генов и мутации.
Несмотря на неоднозначное восприятие в обществе, эволюция как естественный процесс является твёрдо установленным научным фактом, имеет огромное количество доказательств и не вызывает сомнений в научном сообществе[3]. В то же время отдельные аспекты теорий, объясняющих механизмы эволюции, являются предметом научных дискуссий. Открытия в эволюционной биологии оказали огромное влияние не только на традиционные области биологии, но и на другие академические дисциплины, например, антропологию, психологию. Представления об эволюции стали основой современных концепций сельского хозяйства, охраны окружающей среды, широко используются в медицине, биотехнологии и многих других социально значимых областях человеческой деятельности[4].
Содержание
История развития представлений об эволюции
Предположение о том, что живые организмы могут изменяться, впервые встречается у греческих философов-досократиков. Представитель милетской школы Анаксимандр считал, что все животные произошли из воды, после чего вышли на сушу. Человек, по его представлениям, зародился в теле рыбы[5]. У Эмпедокла можно найти идеи гомологии и выживания наиболее приспособленных[6]. Демокрит считал, что наземные животные произошли от земноводных, а те в свою очередь самозародились в илу[7]. В отличие от этих материалистических взглядов, Аристотель считал все природные вещи несовершенными проявлениями различных постоянных естественных возможностей, известных как «формы», «идеи» или (в латинской транскрипции) «виды» (лат. species)[8][9]. Это было частью его телеологического понимания природы, в рамках которого у каждой вещи есть своё предназначение в божественном космическом порядке. Вариации этой идеи стали основой средневекового миропонимания и были объединены с христианским учением. Однако Аристотель не постулировал того, что реальные типы животных являются точными копиями метафизических форм, и приводил примеры, как могут образовываться новые формы живых существ[10].
В XVII веке появился новый метод, который отклонял аристотелевский подход и искал объяснения естественных явлений в законах природы, единых для всех видимых вещей и не нуждающихся в неизменяемых естественных типах или божественном космическом порядке. Но этот новый подход с трудом проникал в биологические науки, которые стали последним оплотом понятия неизменного естественного типа. Джон Рей использовал для животных и растений более общий термин для определения неизменных природных типов — «вид» (лат. species), но в отличие от Аристотеля он строго определил каждый тип живого существа как вид и считал, что каждый вид может быть определён по чертам, которые воспроизводятся от поколения к поколению. По Рею эти виды созданы Богом, но могут быть изменчивы в зависимости от местных условий[11][12]. Биологическая классификация Линнея также рассматривала виды неизменными и созданными по божественному плану[13][14].
Однако в то время были и натуралисты, которые размышляли об эволюционном изменении организмов, происходящем в течение длительного времени. Мопертюи писал в 1751 году об естественных модификациях, происходящих во время воспроизводства, накапливающихся в течение многих поколений и приводящих к формированию новых видов. Бюффон предположил, что виды могут дегенерировать и превращаться в другие организмы[15]. Эразм Дарвин считал, что все теплокровные организмы возможно происходят от одного микроорганизма (или «филамента»)[16]. Первая полноценная эволюционная концепция была предложена Жаном Батистом Ламарком в 1809 году в труде «Философия зоологии». Ламарк считал, что простые организмы (инфузории и черви) постоянно самозарождаются. Затем эти формы изменяются и усложняют своё строение, приспосабливаясь к окружающей среде. Эти приспособления происходят за счёт прямого влияния окружающей среды путём упражнения или неупражнения органов и последующей передачи этих приобретённых признаков потомкам[17][18] (позже эта теория получила название ламаркизм). Эти идеи были отвергнуты натуралистами, поскольку не имели экспериментальных доказательств. Кроме того, всё ещё были сильны позиции учёных, считавших, что виды неизменны, а их сходство свидетельствует о божественном замысле. Одним из самых известных среди них был Жорж Кювье[19].
Концом доминирования в биологии представлений о неизменности видов стала теория эволюции посредством естественного отбора, сформулированная Чарльзом Дарвином. Частично под воздействием «Опыта закона о народонаселении» Томаса Мальтуса, Дарвин заметил, что прирост населения ведёт к «борьбе за существование», в ходе которой начинают преобладать организмы с благоприятными признаками, поскольку те, у кого их нет, погибают. Этот процесс начинается, если каждое поколение производит больше потомства, чем может выжить, что приводит к борьбе за ограниченные ресурсы. Это могло объяснить происхождение живых существ от общего предка за счёт законов природы[20][21]. Дарвин развивал свою теорию начиная с 1838 года, пока Альфред Уоллес в 1858 году не прислал ему свою работу с такими же идеями. Статья Уоллеса была опубликована в том же году в одном томе трудов Линневского общества вместе с краткой выдержкой из работ Дарвина[22]. Публикация в конце 1859 года книги Дарвина «Происхождение видов», в которой детально объясняется концепция естественного отбора, привела к более широкому распространению дарвиновской концепции эволюции.
Точные механизмы наследственности и появления новых черт оставались неизвестными. С целью объяснения этих механизмов Дарвин развивал «временную теорию пангенезиса»[23]. В 1865 году Грегор Мендель открыл законы наследственности, однако его работы оставались практически неизвестными до 1900 года[24]. Август Вейсманн отметил важное различие между зародышевыми (половыми) и соматическими клетками, а также то, что наследственность обусловлена только зародышевой линией клеток. Хуго де Фриз соединил дарвиновскую теорию пангенезиса с вейсманновскими представлениями о половых и соматических клетках и предположил, что пангены расположены в ядре клетки и могут перемещаться в цитоплазму и изменять структуру клетки. Де Фриз был также одним из учёных, которые сделали работу Менделя известной. Он полагал, что менделевские наследственные черты соответствуют передаче наследственных изменений по зародышевому пути. Чтобы объяснить возникновение новых черт, де Фриз развивал теорию мутаций, которая стала одной из причин временного разногласия между зарождающейся генетикой и дарвинизмом[25]. Работы пионеров популяционной генетики, таких как Дж. Б. С. Холдейн, Сьюэл Райт, Рональд Фишер, ставят исследования эволюции на статистическую основу и, таким образом, устраняют это ложное противопоставление генетики и эволюции путём естественного отбора[26].
В 1920-х — 30-х годах XX века современный эволюционный синтез соединил естественный отбор, теорию мутаций и менделевское наследование в единую теорию, применимую к любому разделу биологии. Открытая в 1953 году Уотсоном и Криком структура ДНК продемонстрировала материальную основу наследственности. Молекулярная биология улучшила наше понимание взаимосвязи генотипа и фенотипа. Достижения произошли и в филогенетической систематике. Благодаря публикации и использованию филогенетических деревьев появилась возможность изучать и сравнивать изменения признаков в разных филогенетических группах. В 1973 году эволюционный биолог Феодосий Добжанский писал: «Ничто в биологии не имеет смысл кроме как в свете эволюции», потому что эволюция объединила то, что сначала казалось бессвязными фактами, в непротиворечивую систему знаний, объясняющую и предсказывающую различные факты о жизни на Земле[27].
С тех пор современный синтез был расширен для объяснения биологических явлений на всех уровнях организации живого[1][28]. Так, в 1960-х годах Мотоо Кимура показал, что подавляющее число мутаций на молекулярном уровне носит нейтральный по отношению к естественному отбору характер[29], а в 1972 году палеонтологи Нильс Элдридж и Стивен Гулд возродили дискуссию о прерывистом характере эволюционного процесса[30]. В конце XX века эволюционная биология получила импульс от исследований в области индивидуального развития. Открытие hox-генов и более полное понимание генетического регулирования эмбриогенеза помогли установить роль онтогенеза в филогенетическом развитии и сформировали представление об эволюции новых форм на основе прежнего набора структурных генов и сохранения схожих программы развития у филогенетически далёких организмов[31][32].
Введение
Эволюция происходит в течение периода времени, превышающего срок жизни одного поколения, и заключается в изменении наследуемых черт организма. Первым этапом этого процесса является изменение частот аллелей генов в популяции. В идеальной популяции, в которой отсутствуют мутации, естественный отбор, изоляция (избирательность скрещивания), дрейф и поток генов, частота аллелей, согласно закону Харди — Вайнберга, будет неизменна от поколения к поколению[33]. Таким образом, эволюция в данной популяции не будет происходить. Однако в реальных популяциях существуют факторы, которые приводят к изменениям частот аллелей генов. Это мутации, поток генов, естественный отбор и дрейф генов. Эти факторы и являются факторами эволюции[34][35]. Мутации увеличивают изменчивость в популяции за счёт появления новых аллельных вариантов генов — мутационной изменчивости. Кроме мутационной есть также комбинативная изменчивость, обусловленная рекомбинацией. Обычно она приводит не к изменениям частот аллелей, а к их новым сочетаниям. Однако генная конверсия[36] может приводить как к появлению новых аллелей, так и к изменениям частот аллелей. Ещё одним фактором, приводящим к изменению частот аллелей, является поток генов.
Два других фактора эволюции — естественный отбор и дрейф генов — «сортируют» созданную мутациями и потоком генов изменчивость, приводя к установлению новой частоты аллелей в популяции. Дрейф генов — вероятностный процесс изменения частот генов, и наиболее ярко он проявляется в популяциях относительно небольшого размера. Дрейф может приводить к полному исчезновению определённых аллелей из популяции. Естественный отбор является главным творческим фактором эволюции. Под его действием особи с определённым фенотипом (и определённым набором наследственных черт) в условиях конкуренции будут более успешны, чем другие, то есть будут иметь более высокую вероятность выжить и оставить потомство. Таким образом, в популяции будет увеличиваться доля таких организмов, у которых есть наследственные черты, обладающие селективным преимуществом. Взаимное влияние дрейфа и естественного отбора однозначно оценить сложно, но в целом, вероятно, оно зависит от размера популяции и интенсивности отбора. Кроме выше названных факторов, важное значение может иметь и горизонтальный перенос генов, который может приводить к появлению совершенно новых для данного организма генов.
Естественный отбор приводит к формированию адаптаций и повышению приспособленности. Протекающие в течение длительного времени эволюционные процессы могут привести как к образованию новых видов и их дальнейшей дивергенции, так и к вымиранию целых видов.
Наследственность
Наследственность — это свойство организмов повторять в ряду поколений сходные типы обмена веществ и индивидуального развития в целом. Эволюция организмов происходит посредством изменения наследственных признаков организма. Примером наследственного признака у человека может служить коричневый цвет глаз, унаследованный от одного из родителей[37]. Наследственные признаки контролируются генами. Совокупность всех генов организма образует его генотип[38].
Полный набор поведенческих и структурных черт организма называют фенотипом. Фенотип организма образуется за счёт взаимодействия генотипа с окружающей средой. Многие черты фенотипа являются ненаследственными. Так, например, загар не наследуется, поскольку его появление обусловлено воздействием солнечного света. Однако у некоторых людей загар появляется легче, чем у других. Это является наследственной чертой.
Передача наследственных признаков от одного поколения к другому обеспечивается ДНК[38]. ДНК — это биополимер, состоящий из четырёх типов нуклеотидов. Во время деления клетки ДНК копируется — в результате каждая из дочерних клеток получает точную копию материнских молекул ДНК.
Участки молекулы ДНК, определяющие функциональную единицу наследственности, называются генами. Внутри клеток ДНК находится в составе хроматина, который в свою очередь образует хромосомы. Положение гена на хромосоме называется локусом. Различные формы гена, расположенные в одинаковых локусах гомологичных хромосом и определяющие различные проявления признаков, называются аллелями. Последовательность ДНК может изменяться (мутировать), создавая новые аллели. Если мутация происходит внутри гена, то новая аллель может затронуть признак, управляемый геном, и изменить фенотип организма.
Однако большинство признаков определяется не одним геном, а взаимодействием нескольких генов (примером подобных явлений могут служить эпистаз и полимерия)[39][40]. Исследование подобных взаимодействий генов — одна из главных задач современной генетики.
Исследования последних лет подтвердили существование изменений в наследственности, которые не могут быть объяснены изменениями в последовательности нуклеотидов ДНК. Этот феномен известен как эпигенетические системы наследования[41]. К таким системам относятся профиль метилирования ДНК, самоподдерживающиеся метаболические петли, сайленсинг генов за счёт РНК-интерференции и другие механизмы[42][43].
Наследуемость также может встречаться и в более крупных масштабах. Например, экологическое наследование посредством строительства ниши[44]. Таким образом, потомки наследуют не только гены, но и экологические особенности местообитания, созданного активностью предков. Другими примерами наследования, не находящегося под контролем генов, могут служить наследование культурных черт и симбиогенез[45][46].
Изменчивость
Изменчивость — это свойство организма отличаться от родителей, а также свойство особей одного вида отличаться друг от друга. Дарвин полагал, что движущими силами эволюции являются: определённая изменчивость (которая повышает приспособленность организмов, например, листопад у лиственных деревьев), неопределённая изменчивость (которая не повышает приспособленности, например, простуда может вызывает кашель, насморк и ревматизм), а также борьба за существование и естественный отбор[47]. Особое значение Дарвин придавал наследственной изменчивости, которая даёт сырой материал для естественного отбора. Также следствием наследственной изменчивости является генетическое разнообразие, которое возникает либо в результате сочетания разных признаков родителей (комбинативная изменчивость), либо в ходе мутационного процесса (мутационная изменчивость).
Фенотип организма обусловлен его генотипом и влиянием окружающей среды. Существенная часть вариаций фенотипов в популяциях вызвана различиями их генотипов[40]. В СТЭ эволюция определяется как изменение с течением времени генетической структуры популяций. Частота одной из аллелей изменяется, становясь более или менее распространённой среди других форм этого гена. Действующие силы эволюции ведут к изменениям в частоте аллели в одну или в другую стороны. Изменение исчезает, когда новая аллель достигает точки фиксации — целиком заменяет предковую аллель или исчезает из популяции[48].
Изменчивость складывается из мутаций, потока генов и рекомбинации генетического материала. Изменчивость также увеличивается за счёт обменов генами между разными видами, таких как горизонтальный перенос генов у бактерий[49], гибридизация у растений[50]. Несмотря на постоянное увеличение изменчивости за счёт этих процессов, большая часть генома идентична у всех представителей данного вида[51]. Однако даже сравнительно небольшие изменения в генотипе могут вызвать огромные различия в фенотипе, например, геномы шимпанзе и людей различаются всего на 5 %[52].
Мутации
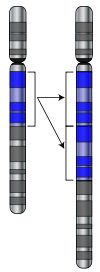 Дупликация участка хромосомы
Дупликация участка хромосомы
Случайные мутации постоянно происходят в геномах всех организмов. Эти мутации создают генетическую изменчивость. Мутации — изменения в последовательности ДНК. Они могут быть вызваны радиацией, вирусами, транспозонами, мутагенными веществами, а также ошибками, происходящими во время репликации ДНК или мейоза[53][54][55]. Мутации могут не иметь никакого эффекта, могут изменять продукт гена или препятствовать его функционированию. Исследования, проведённые на дрозофиле, показали, что если мутация изменяет белок, производимый геном, то примерно в 70% случаев это будет иметь вредные воздействия, а в остальных случаях нейтральные или слабоположительные[56]. Для уменьшения негативного эффекта мутаций в клетках существуют механизмы репарации ДНК[53]. Оптимальный уровень мутаций — это баланс между высоким уровнем вредных мутаций и затратами на поддержание системы репарации[57]. У РНК-вирусов уровень мутабильности высокий[58], что, видимо, является преимуществом, помогая избегать защитных ответов иммунной системы[59].
Мутации могут затрагивать большие участки хромосом. Например, при дупликации, которая вызывает появление дополнительных копий гена в геноме[60]. Эти копии становятся основным материалом для возникновения новых генов. Это важный процесс, поскольку новые гены развиваются в пределах семейства генов от общего предка[61]. Например, в образовании светочувствительных структур в глазу человека участвуют четыре гена: три для цветного зрения и один для ночного зрения. Все эти гены произошли от одного предкового гена[62]. Новые гены возникают из предкового гена в результате дупликации, после того как копия гена мутирует и приобретёт новую функцию. Этот процесс идёт легче после дупликации, поскольку это увеличивает избыточность системы. Один ген из пары может приобрести новую функцию, в то время как другой продолжает выполнять основную функцию[63][64]. Другие типы мутаций могут создать новые гены из некодирующей ДНК[65][66]. Новые гены также могут возникнуть за счёт рекомбинации небольших участков дублированных генов. При этом возникает новая структура с новыми функциями[67][68]. Когда новые гены собраны перетасовкой ранее существовавших частей (выполняющих простые независимые функции), то их комбинация может выполнять новые более сложные функции. Примером подобного комплекса являются поликетидсинтазы — ферменты, синтезирующие вторичные метаболиты, например антибиотики. Они могут содержать до 100 частей, катализирующих один этап в процессе полного синтеза[69].
Изменения на хромосомном уровне могут приводить к ещё большим мутациям. Среди хромосомных мутаций выделяют делеции и инверсии крупных участков хромосом, транслокацию участков одной хромосомы на другую. Также возможно слияние хромосом (робертсоновская транслокация (англ.)русск.). Например, в ходе эволюции рода Homo произошло слияние двух хромосом с образованием второй хромосомы человека[70]. У других обезьян этого слияния не произошло, и хромосомы сохраняются по отдельности. В эволюции хромосомные перестройки играют большую роль. Благодаря им может ускориться расхождение популяций с образованием новых видов, поскольку уменьшается вероятность скрещивания и таким образом увеличиваются генетические различия между популяциями[71].
Мобильные элементы генома, такие как транспозоны, составляют значительную долю в геномах растений и животных и, возможно, важны для эволюции[72]. Так, в человеческом геноме около 1 миллиона копий Alu-повтора, они выполняют некоторые функции, такие как регулирование экспрессии генов[73]. Другое влияние мобильных генетических элементов на геном заключается в том, что при их перемещении внутри генома может произойти видоизменение или удаление существующих генов[54].
Рекомбинация
У бесполых организмов гены во время размножения не могут смешиваться с генами других особей. В отличие от них у организмов с половым размножением потомство получает случайные смеси хромосом своих родителей. Кроме этого, в ходе мейотического деления половых клеток происходит кроссинговер, приводящий к гомологичной рекомбинации, которая заключается в обмене участками двух гомологичных хромосом[74]. При рекомбинации не происходит изменения частоты аллелей, но происходит образование их новых комбинаций[75]. Таким образом, половое размножение обычно увеличивает наследственную изменчивость и может ускорять темп эволюции организмов[76][77]. Однако бесполое размножение зачастую выгодно и может развиваться у животных с половым размножением[78].
Гомологичная рекомбинация позволяет наследоваться независимо даже аллелям, которые находятся близко друг от друга в ДНК. Однако уровень кроссинговера низок — примерно два на хромосому за поколение. В результате гены, находящиеся рядом на хромосоме, имеют тенденцию наследоваться сцеплено. Это тенденция измеряется тем, как часто две аллели находятся вместе на одной хромосоме, и носит название неравновесного сцепления генов[79]. Несколько аллелей, которые наследуются вместе, обычно называют гаплотипом. Если одна аллель в гаплотипе даёт значительное преимущество, то в результате естественного отбора частота в популяции других аллелей этого гаплотипа также может повыситься. Это явление называется genetic hitchhiking (англ.)русск. («передвижение автостопом»)[80].
Когда аллели не могут быть разделены рекомбинацией, как в Y-хромосоме млекопитающих, которая передаётся без изменений от отца к сыну, происходит накопление вредных мутаций[81]. Половое размножение за счёт изменения комбинаций аллелей позволяет удалять вредные мутации и сохранять полезные[82]. Подобные положительные эффекты уравновешиваются негативными явлениями полового размножения, такими как уменьшение репродуктивного уровня и возможные нарушения выгодных комбинаций аллелей при рекомбинации. Поэтому причины возникновения полового размножения до сих пор не ясны и активно исследуются в эволюционной биологии[83][84], порождая такие гипотезы, как гипотеза Чёрной королевы[85].
Поток генов
Потоком генов называют перенос аллелей генов между популяциями. Поток генов может осуществляться за счёт миграций особей между популяциями в случае подвижных организмов либо, например, с помощью переноса пыльцы или семян в случае растений. Скорость потока генов сильно зависит от подвижности организмов[86].
Степень влияния потока генов на изменчивость в популяциях до конца не ясна. Существуют две точки зрения: одна из них заключается в том, что поток генов может иметь значительное влияние на крупных популяционных системах, гомогенизируя их и, соответственно, действуя против процессов видообразования; вторая, что скорости потока генов достаточно только для воздействия на локальные популяции[86][87].
Механизмы эволюции
Естественный отбор
 Мутации и естественный отбор создают популяцию с более тёмной окраской
Мутации и естественный отбор создают популяцию с более тёмной окраскойЭволюция посредством естественного отбора — это процесс, при котором закрепляются мутации, увеличивающие приспособленность организмов. Естественный отбор часто называют «самоочевидным» механизмом, поскольку он следует из таких простых фактов, как:
- Организмы производят потомков больше, чем может выжить;
- В популяции этих организмов существует наследственная изменчивость;
- Организмы, имеющие разные генетические черты, имеют различную выживаемость и способность размножаться.
Такие условия создают конкуренцию между организмами в выживании и размножении и являются минимально необходимыми условиями для эволюции посредством естественного отбора[88]. Таким образом, организмы с наследственными чертами, которые дают им конкурентное преимущество, имеют большую вероятность передать их своим потомкам, чем организмы с наследственными чертами, не имеющими подобного преимущества[89].
Центральное понятие концепции естественного отбора — приспособленность организмов[90]. Приспособленность определяется как способность организма к выживанию и размножению, которая определяет размер его генетического вклада в следующее поколение[90]. Однако главным в определении приспособленности является не общее число потомков, а число потомков с данным генотипом (относительная приспособленность)[91]. Например, если потомки успешного и быстро размножающегося организма слабые и плохо размножаются, то генетический вклад и, соответственно, приспособленность этого организма будут низкими[90].
Если какая-либо аллель увеличивает приспособленность организма больше, чем другие аллели этого гена, то с каждым поколением доля этой аллели в популяции будет расти. То есть, отбор происходит в пользу этой аллели. И наоборот, для менее выгодных или вредных аллелей — их доля в популяциях будет снижаться, то есть отбор будет действовать против этих аллелей[92]. Важно отметить, что влияние определённых аллелей на приспособленность организма не является постоянным — при изменении условий окружающей среды вредные или нейтральные аллели могут стать полезными, а полезные вредными[93].
Естественный отбор для черт, которые могут изменяться в некотором диапазоне значений (например, размер организма), можно разделить на три типа[94]:
- Направленный отбор — изменения среднего значения признака в течение долгого времени, например увеличение размеров тела;
- Дизруптивный отбор — отбор на крайние значения признака и против средних значений, например, большие и маленькие размеры тела;
- Стабилизирующий отбор — отбор против крайних значений признака, что приводит к уменьшению дисперсии признака и уменьшению разнообразия.
Частным случаем естественного отбора является половой отбор, субстратом которого является любой признак, который увеличивает успешность спаривания за счёт увеличения привлекательности особи для потенциальных партнёров[95]. Черты, которые эволюционировали за счёт полового отбора, особенно хорошо заметны у самцов некоторых видов животных. Такие признаки, как крупные рога, яркая окраска, с одной стороны могут привлекать хищников и понижать выживаемость самцов[96], а с другой это уравновешивается репродуктивным успехом самцов с подобными ярко выраженными признаками[97].
Отбор может действовать на различных уровнях организации, таких как гены, клетки, отдельные организмы, группы организмов и виды[98][99][100]. Причём отбор может одновременно действовать на разных уровнях[101]. Отбор на уровнях выше индивидуального, например, групповой отбор, может приводить к кооперации (см. Эволюция#Кооперация)[102].
Дрейф генов
 Моделирование дрейфа генов для 20 несцепленных аллелей в популяции из 10 (сверху) и 100 (снизу) особей. Фиксация аллелей происходит быстрее в популяции с меньшим размером
Моделирование дрейфа генов для 20 несцепленных аллелей в популяции из 10 (сверху) и 100 (снизу) особей. Фиксация аллелей происходит быстрее в популяции с меньшим размеромДрейф генов — изменение частоты аллелей генов от поколения к поколению, обусловленное случайными процессами при выживании и размножении организмов[103]. В математических терминах аллели подвергаются ошибке выборки. Если действие отбора относительно слабое или отсутствует, то частота аллелей имеет тенденцию случайно «дрейфовать» вверх или вниз (случайное блуждание). Дрейф останавливается, когда аллель фиксируется в популяции — исчезая или полностью заменяя другие аллели этого гена в популяции. Таким образом, за счёт только случайных процессов из популяции могут быть элиминированы некоторые аллели[104].
Время, которое требуется для фиксации аллелей, зависит от размера популяции[105]. При этом главным фактором оказывается не число особей, а эффективный размер популяции[106]. Эффективный размер популяции обычно меньше, чем общее число особей в популяции. При его расчёте принимаются во внимание уровень инбридинга, стадия жизненного цикла, при которой численность популяции наименьшая, и тот факт, что некоторые селективно-нейтральные гены сцеплены с генами, подверженными действию отбора[106].
Обычно трудно определить относительную важность естественного отбора и дрейфа[107], сравнительная важность этих двух факторов в эволюционных процессах является предметом современных исследований[108].
Нейтральная теория молекулярной эволюции предполагает, что большинство эволюционных изменений является результатом фиксации нейтральных мутаций в геноме, которые не оказывают никакого влияния на приспособленность организмов[29]. Следовательно, в этой модели большинство генетических изменений в популяции — эффект давления мутаций и дрейфа генов[109][110]. Эта трактовка теории нейтральности в настоящее время не принимается учёными, поскольку не соответствует характеру генетической изменчивости в природе[111][112]. Однако есть более современная и поддерживаемая учёными версия этой теории — nочти нейтральная теория (англ.)русск., в рамках которой мутации нейтральные в небольших популяциях не обязательно будут нейтральными в крупных[113]. Альтернативные теории предполагают, что генетический дрейф маскируется другими стохастическими влияниями, такими как genetic hitchhiking (англ.)русск.[103][114][115].
Горизонтальный перенос генов
Горизонтальный перенос генов представляет собой передачу генетического материала от одного организма другому организму, который не является его потомком. Наиболее распространён горизонтальный перенос генов у бактерий[116]. В частности, это способствует распространению устойчивости к антибиотикам, посредством того, что после возникновения у одной бактерии генов устойчивости они могут быстро передаться другим видам[117]. Известна также возможность горизонтального переноса генов от бактерий к эукариотам, таким как дрожжи Saccharomyces cerevisiae и обитающий на адзуки жук Callosobruchus chinensis[118][119]. Примером крупномасштабного переноса генов среди эукариот могут служить бделлоидные коловратки, которые получили ряд генов от бактерий, грибов и растений[120]. Переносчиками ДНК между представителями отдельных доменов могут быть вирусы[121] .
Масштабный перенос генов также произошёл между предками клеток эукариот и бактерий, во время приобретения первыми хлоропластов и митохондрий. Вполне возможно, что эукариоты произошли в результате горизонтальных переносов генов между бактериями и археями[122].
Результаты эволюции
Эволюция влияет на все аспекты жизнедеятельности организмов. Один из самых заметных для наблюдения результатов эволюции — адаптация (поведенческая, морфологическая или физиологическая), которая является результатом естественного отбора. Адаптация увеличивает приспособленность организмов. В долгосрочной перспективе эволюционные процессы могут привести к появлению новых видов, например, после разделения передковой популяции организмов на новые популяции, особи из которых не могут скрещиваться.
Эволюцию иногда разделяют на макроэволюцию, то есть эволюцию, происходящую на уровне вида и выше, к ней относятся такие процессы, как видообразование и вымирание, и микроэволюцию, то есть эволюцию, происходящую ниже видового уровня, например, адаптация в популяции[123]. В целом макроэволюция рассматривается как результат длительной микроэволюции[124]. То есть между макроэволюцией и микроэволюцией нет фундаментальных различий — отличается только необходимое время[125]. Однако, в случае макроэволюции, важными могут быть черты целых видов. Например, большое количество различий между особями позволяет виду быстро приспосабливаться к новым средам обитания, уменьшая шанс вымирания, а большой ареал вида увеличивает шанс видообразования.
Распространённым заблуждением относительно эволюции является утверждение о том, что у неё есть некая цель или долгосрочный план. В действительности у эволюции нет ни целей, ни планов[~ 1], а также в ходе эволюции не обязательно происходит возрастание сложности организмов[126][127][128][129]. Хотя в ходе эволюции и возникают сложно организованные организмы, они являются «побочным продуктом» эволюции, а наиболее распространёнными в биосфере являются более «простые» организмы[130]. Так, микроскопические прокариоты представлены огромным числом видов и составляют более половины всей биомассы[131] и значительную часть биоразнообразия[132].
Адаптация
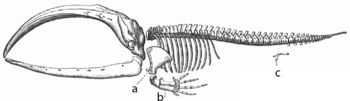 Скелет усатого кита: a, b — кости плавников, произошедшие от костей передних конечностей; с — рудиментарные кости задних конечностей[133]
Скелет усатого кита: a, b — кости плавников, произошедшие от костей передних конечностей; с — рудиментарные кости задних конечностей[133]Адаптация — процесс, результатом которого является улучшение приспособленности организма к местообитанию. Также термин «адаптация» может относиться к черте, важной для выживания организма. Например, адаптация зубов лошади к пережёвыванию травы. Поэтому существует два различных значения для понятия «адаптация» — адаптация как эволюционный процесс и адаптивная черта как его продукт. Адаптация является следствием естественного отбора[134]. Следующие определения адаптации даны Феодосием Добжанским:
- Адаптация — эволюционный процесс, благодаря которому организм становится лучше приспособлен к месту или местам обитания.
- Адаптивность — возможность адаптации: степень, до которой организм способен жить и размножаться в данном наборе мест обитания.
- Адаптивная черта — черта организма, позволяющая или увеличивающая вероятность того, что организм выживет и размножится[135].
Адаптация может привести как к появлению новой черты, так и к потере предковой черты. Примером, демонстрирующим оба типа изменений, является устойчивость бактерий к антибиотикам, или антибиотикорезистентность. Генетической основой для этой устойчивости может быть либо модификация молекул бактериальных клеток, на которые воздействуют антибиотики, либо усиление деятельности транспортёров, выводящих антибиотики из клетки[136]. Другими примерами приобретения адаптивных черт являются бактерии Escherichia coli, у которых в ходе долговременного эксперимента появилась способность к использованию в качестве источника углерода лимонной кислоты[137]; Flavobacterium, у которых появился новый фермент, позволяющий им использовать побочные продукты производства нейлона[138][139], и почвенная бактерия Sphingobium, у которой развился полностью новый метаболический путь разрушения синтетического пестицида пентахлорфенола[140][141]. Интересная, но все ещё спорная идея состоит в том, что некоторые адаптации могут повышать способность организмов к генетической изменяемости, повышая генетическое разнообразие, что в дальнейшем увеличивает вероятность адаптироваться в результате естественного отбора (повышение эволюционной способности организмов)[142][143][144][145].
Адаптация происходит посредством постепенного изменения структур. Следовательно, у структур со схожим строением у родственных организмов могут быть разные функции. Это является результатом того, что предковые структуры адаптируются для выполнения разных функций. Например, кости в крыльях летучих мышей гомологичны костям передних конечностей других млекопитающих, поскольку происходят от одних и тех же костей общего предка всех млекопитающих[146]. Так как все живые организмы родственны в той или иной степени[147], то даже те органы разных групп организмов, у которых мало или вообще нет схожести, такие как глаза членистоногих, кальмаров и позвоночных или конечности членистоногих и позвоночных, могут зависеть от схожего набора гомологичных генов, контролирующих их формирование и функции. Это называется глубокой гомологией (англ.)русск.[148][149].
В ходе эволюции некоторые структуры могут терять свою основную функцию, становясь рудиментами[150]. У таких структур могут оставаться вторичные функции, либо они полностью утрачивают функциональность. Примерами таких структур являются псевдогены[151], не функционирующие остатки глаз у пещерных рыб[152], крылья у нелетающих птиц[153], тазовые кости у китообразных и змей[133]. Примером рудиментов у человека являются зубы мудрости[154], копчик[150], аппендикс[150], а также физиологические реакции, такие как гусиная кожа[155] и врождённые рефлексы[156].
Однако многие черты, кажущиеся простыми адаптациями, в действительности являются результатом экзаптации, процесса, посредством которого формы или структуры, развившиеся в ходе эволюции, чтобы выполнять одну функцию, кооптируются, чтобы обслуживать другие функции[157]. Примером экзаптации могут быть молекулярные машины, такие как бактериальный жгутик[158] и митохондриальные транслоказные комплексы[159], которые произошли сборкой нескольких белков, до этого выполнявших другие функции[123].
Один из основополагающих экологических принципов — принцип конкурентного исключения, он заключается в том, что два вида не могут занимать одну и ту же экологическую нишу в течение длительного времени в той же самой среде[160]. Следовательно, под действием естественного отбора виды будут иметь тенденцию адаптироваться к разным экологическим нишам.
Коэволюция
Взаимодействие между организмами может вызвать как конфликт, так и кооперацию. Если взаимодействуют пары организмов, такие как патоген и его хозяин или хищник и жертва, то у них могут развиваться связанные адаптации. Эволюция одного вида в таких парах вызывает эволюцию у другого вида, а это, в свою очередь, вызывает эволюцию первого вида и т. д. Этот цикл носит название коэволюции[161]. Примером коэволюции может служить пара: жертва — Желтобрюхий тритон, хищник — Подвязочная змея (англ. Common Garter Snake). Эволюционная гонка вооружений привела к тому, что у тритона вырабатывается большое количество яда тетродотоксина, а у змеи высокий уровень устойчивости к этому яду[162].
Кооперация
Не все случаи совместной эволюции основаны на конфликте между разными видами[163][164]. Известно много случаев взаимовыгодных отношений. Например, очень тесная связь существует между растениями и мицелиями грибов, которые растут на их корнях и помогают растениям получать питательные вещества из почвы[165]. Это взаимовыгодные отношения, поскольку грибы получают от растений сахара. Для обмена питательными веществами грибы фактически прорастают внутрь клеток растений, подавляя их защитные системы[166].
Также существует и кооперация между особями одного вида. Крайний случай — эусоциальные насекомые, такие как пчёлы, термиты и муравьи, у которых бесплодные насекомые кормят и охраняют небольшое количество способных к размножению особей.
Такое сотрудничество в рамках одного вида, возможно, появилось благодаря кин-отбору, который направлен на отбор признаков, способствующих выживанию близких родичей данной особи[167]. Отбор действует в пользу аллелей, способствующих помощи близким родичам, потому что, если у особи есть эти аллели, то высока вероятность, что и у её близких родичей также есть эти аллели[168]. Другие процессы, способствующие кооперации между особями, включают групповой отбор, при котором взаимодействия между особями даёт преимущества для их групп, по сравнению с другими группами, у которых нет такого взаимодействия[169].
Видообразование
Видообразование — процесс образования новых видов. Видообразование наблюдалось многократно и в контролируемых лабораторных условиях, и в природе[170][171][172].
Есть несколько подходов к определению понятия «вид», различающихся тем, какие черты организмов для этого используются[173]. Например, некоторые концепции вида больше подходят для организмов, размножающихся половым способом, другие — для организмов, размножающихся бесполым. Несмотря на различие подходов, они могут быть объединены в три подхода: гибридологический, экологический и филогенетический[174]. Биологический критерий вида, предложенный в 1942 году Эрнстом Майером, — яркий пример гибридологического подхода. По Майеру, вид — «группы фактически или потенциально скрещивающихся естественных популяций, которые физиологически изолированы от других таких групп»[175]. Несмотря на широкое и длительное использование этого критерия, он имеет свои недостатки, особенно в случае прокариот[176].
 Географическая изоляция на Галапагосских островах привела к образованию более десяти новых видов вьюрков
Географическая изоляция на Галапагосских островах привела к образованию более десяти новых видов вьюрковНаличие репродуктивных барьеров между двумя дивергирующими популяциями необходимо для образования новых видов. Поток генов может замедлить процесс образования этого барьера, распространяя новые генетические черты между обеими дивергирующими популяциями. В зависимости от того, как далеко от общего предка дивергировали два новых вида, у них может сохраниться способность скрещиваться и давать потомство. Например, как в случае лошадей и ослов, которые могут скрещиваться и давать потомство — мулов. Однако обычно такое потомство бесплодно. В этом случае два близких вида могут регулярно скрещиваться, но отбор действует против гибридов. Однако в некоторых случаях гибриды могут быть плодовитыми, и эти новые виды могут иметь либо промежуточные черты между двумя родительскими видами, а могут обладать и совсем новым фенотипом[177]. Полностью значение гибридизации для видообразования неясно, однако примеры есть во многих типах животных[178]. Наиболее изученным является случай Серой квакши (Hyla chrysoscelis[en])[179].
 Четыре способа видообразования: аллопатрическое, перипатрическое, парапатрическое и симпатрическое
Четыре способа видообразования: аллопатрическое, перипатрическое, парапатрическое и симпатрическоеСуществует четыре способа видообразования. Наиболее распространённым у животных является аллопатрическое видообразование (англ.)русск., происходящее в популяциях, разделённых географически, например из-за фрагментации мест обитания или за счёт миграций. Естественный отбор в этих условиях может очень быстро привести к изменениям в строении и поведении организмов[180][181]. Поскольку отбор и дрейф генов действуют независимо в популяции, изолированной от остальной части вида, то постепенно это разделение может привести к появлению репродуктивной изоляции[182].
Второй способ видообразования — это перипатрическое видообразование, которое происходит в случае изоляции небольшой популяции в новой окружающей среде. Основное отличие от аллопатрического видообразования состоит в том, что изолированная популяция значительно меньше предковой популяции. В новой небольшой популяции эффект основателя вызывает быстрое видообразование за счёт близкородственного скрещивания и отбора на гомозиготы, что приводит к быстрым генетическим изменениям[183].
Третий способ видообразования — парапатрическое видообразование (англ.)русск.. Оно похоже на перипатрическое видообразование, когда небольшая популяция оказывается изолированной в новой среде обитания, но в случае парапатрического видообразования нет никакого физического разделения между двумя популяциями. Вместо этого видообразование является следствием процессов, уменьшающих поток генов между популяциями[184]. Обычно подобный тип видообразования встречается при радикальном изменении условий окружающей среды. Один из примеров: Душистый колосок обыкновенный (Anthoxanthum odoratum[en]) может подвергаться парапатрическому видообразованию в ответ на загрязнение почв металлами из-за рудников[185]. В местах загрязнения растения эволюционируют в сторону устойчивости к высокому уровню металлов в почве. Отбор против скрещивания между растениями, устойчивыми к металлам в почве, и растениями предковой популяции вызвал постепенное изменение времени цветения устойчивых к металлам растений, что в конечном счёте привело к появлению репродуктивной изоляции[186].
Четвёртый способ — симпатрическое видообразование — происходит без изоляции или изменений в месте обитания. Этот способ видообразования редок, поскольку даже небольшой уровень потока генов может сгладить генетические различия между частями популяции[187][188]. Для формирования репродуктивной изоляции симпатрическое образование требует как генетических различий, так и неслучайного спаривания[189].
Один тип симпатрического видообразования заключается в гибридизации двух близких видов. Этот тип обычно не встречается у животных, так как хромосомы разных родительских видов не могут конъюгировать во время мейоза, что приводит к бесплодности их гибридов. Однако у растений подобная проблема решается полиплоидизацией[190], которая позволяет из хромосом одного родительского вида формировать биваленты при мейозе[191]. Пример такого видообразования — скрещивание Arabidopsis thaliana и Arabidopsis arenosa (нем.)русск.) с образованием нового вида Arabidopsis suecica[192]. Это произошло около 20 тысяч лет назад[193] и было повторено в лабораторных условиях[194].
Вымирание

Вымирание — исчезновение всех представителей определённого таксона. Вымирания и видообразование происходят постоянно в течение всей истории жизни[195]. Бо́льшая часть видов, когда-либо обитавших на Земле, вымерла[196], и, по всей видимости, вымирание — судьба любого вида[197]. Вымирание происходит постоянно, но некоторые глобальные события приводят к массовому вымиранию видов и других таксонов[198]. Мел-палеогеновое вымирание, частью которого являлось вымирание динозавров, самое известное массовое вымирание, а пермское вымирание — самое крупное — оно привело к вымиранию 96% видов[198]. Голоценовое вымирание (англ.)русск. — массовое вымирание, связанное с человеческой деятельностью за последние несколько тысяч лет. Современные темпы вымирания в 100—1000 раз выше, чем фоновые темпы вымирания, и к середине XXI века может исчезнуть до 30% видов[199]. Деятельность человека является первоочередной причиной нынешнего вымирания видов[200][201], а глобальные изменения климата могут в дальнейшем значительно усугубить ситуацию[202].
Последствия вымирания в эволюции выживших видов не до конца понятны и могут отличаться в зависимости от того, какой тип вымирания рассматривать. Причиной постоянного немассового вымирания может быть конкуренция за ограниченные ресурсы (принцип конкурентного исключения)[1]. Здесь вступает в действие видовой отбор, в результате чего более приспособленный вид выживет, а менее приспособленный может вымереть[98]. Массовые вымирания неспецифическим образом уменьшают биоразнообразие, что может привести к всплескам быстрой эволюции и видообразования у выживших[203].
Эволюция жизни на Земле
Происхождение жизни
Происхождение жизни — необходимый элемент для начала эволюции как таковой, но понимание и исследование эволюционных процессов, начавшихся сразу после возникновения первого организма, не зависят от того, что нам известно о возникновении жизни[204]. На данный момент общепринятой в науке является концепция, согласно которой сложные органические вещества и биохимические реакции произошли от простых химических реакций, однако детали этого процесса до конца не ясны[205]. Также нет точных сведений о ранних этапах развития жизни, строении и особенностях первых организмов и последнего универсального общего предка[206]. Одной из ключевых концепций, описывающих возникновение жизни, является гипотеза мира РНК[207].
Доказательства происхождения от общего предка
 Гоминиды произошли от общего предка
Гоминиды произошли от общего предкаВсе организмы на Земле происходят от общего предка или от предкового пула генов[147]. Ныне живущие организмы являются этапом в эволюции, которому предшествовали процессы видообразования и вымирания[208]. Родство всех организмов было показано за счёт четырёх очевидных фактов. Во-первых, географическое распределение организмов не может быть объяснено только адаптациями к конкретным условиям среды. Во-вторых, разнообразие жизни — это ряд не абсолютно уникальных организмов, а организмов, у которых есть общие черты в строении. В-третьих, наличие рудиментарных органов, напоминающие функционирующие органы у предков. И, в-четвёртых, все организмы могут быть классифицированы на основе общих черт в иерархические вложенные группы[209]. Однако современные исследования показывают, что «дерево жизни» может быть сложнее простого ветвящегося дерева из-за горизонтального переноса генов[210][211].
Вымершие организмы также оставляют «записи» о своей эволюционной истории в виде окаменелостей. Палеонтологи исследуют анатомию и морфологию вымерших видов и, сравнивая их с современными, могут определить пути их эволюции[212]. Однако этот метод подходит в основном для организмов, у которых есть твёрдые части, например, раковины, кости или зубы. Этот палеонтологический метод не подходит для прокариот, поскольку все они обладают схожей морфологией.
Доказательства происхождения от общего предка были также подтверждены общностью биохимических процессов в клетках организмов. Так, практически все организмы используют одни и те же нуклеотиды и аминокислоты[213]. С развитием молекулярной генетики было показано, что процессы эволюции оставляют следы в геномах в виде мутаций. На основе гипотезы молекулярных часов стало возможным определение времени дивергенции видов[214]. Например, геномы шимпанзе и человека одинаковы на 96%, а те немногие области, которые различаются, позволяют определить время существования их общего предка[215].
Краткая хронология эволюции
Первые организмы появились на Земле в результате химической эволюции не позднее 3,5 млрд лет назад[216]. Около 2,5 млрд лет назад у цианобактерий появился кислородный фотосинтез, что привело к оксигенации атмосферы Земли.
Около 2 миллиардов лет назад появились первые эукариотические организмы. Точные детали того, как прокариотические клетки эволюционировали в эукариотические клетки, неизвестны, и на этот счёт существует значительное количество гипотез. Достоверно известно, что некоторые органеллы — митохондрии и хлоропласты — произошли в результате симбиогенеза протеобактерий и цианобактерий соответственно[217][218].
Следующим важным шагом в эволюции жизни на Земле стало появление многоклеточных организмов. Это произошло около 610 млн лет назад в океанах в эдиакарском периоде[219][220].
После появления первых многоклеточных организмов в течение последующих примерно 10 миллионов лет произошло значительное повышение их биоразнообразия, получившее название Кембрийского взрыва. В палеонтологической летописи появляются представители почти всех современных типов живых организмов, а также представители многих вымерших линий[221][222]. Для объяснения причин Кембрийского взрыва были предложены различные гипотезы, в том числе накопление в атмосфере кислорода в результате деятельности фотосинтезирующих организмов[223][224].
Около 500 миллионов лет назад растения и грибы вышли на сушу, вскоре за ними последовали членистоногие и другие животные[225]. Насекомые были одними из самых успешных и в настоящее время составляют большинство видов животных[226]. Около 360 млн лет назад появились земноводные, от них 330 млн лет назад произошли первые амниотические организмы. Чуть позже произошло разделение амниот на две эволюционные линии — зауропсидную (давшую начало рептилиям и птицам) и синапсидную (давшую начало млекопитающим)[227].
Антропогенез
Происхождение и эволюция человека является неотъемлемой частью эволюции живого. Человек разумный (неоантроп) относится к семейству гоминид, отряда приматов. Приматы появились, по разным оценкам, от 65 до 85 млн лет назад[228]. Около 30 млн лет назад надсемейство человекообразных обезьян отделяется от других сухоносых обезьян[228], а около 19 миллионов лет назад гоминиды дивергируют от предков гиббонов[229]. Около 7 млн лет назад появляется сахелантроп, последний общий предок шимпанзе и рода Homo[230]. Дальнейшая эволюция приводит к появлению австралопитеков, от которых, по всей видимости, 2 млн лет назад происходят первые Homo[230]. Около 200 тыс. лет назад появляются анатомически современные люди (англ.)русск.[231]. По современным оценкам, около 190 тыс. лет назад жил последний общий предок ныне живущих людей по материнской линии — митохондриальная Ева[232].
Применение
Концепции и модели, используемые в эволюционной биологии, в частности естественный отбор, имеют большое число применений[233].
Искусственный отбор использовался в течение тысяч лет для выведения новых сортов растений и пород одомашненных животных[234]. В последнее время селекция стала играть значительную роль в генетической инженерии, например, при отборе нужных клонов с помощью селектируемых маркеров.
Понимание того, какие изменения произошли в ходе эволюции организма, может помочь выявить гены, которые участвуют в построении той или иной части тела, или гены, участвующие в развитии наследственных заболеваний[235]. Например, пещерная рыба-альбинос мексиканская тетра в ходе эволюции стала слепой[236]. Скрещивание особей из разных популяций этой рыбы привело к появлению потомков с функционирующими глазами, поскольку в разных популяциях, обитающих в разных пещерах, произошли разные мутации. Это помогло идентифицировать гены, необходимые для зрения[237].
В компьютерных науках использование эволюционных алгоритмов и искусственной жизни было начато с 1960-х годов и впоследствии было расширено симуляцией искусственного отбора[238]. Эволюционные алгоритмы стали признанным методом оптимизации после работ Инго Рехенберга (англ.)русск.. Он использовал эволюционные алгоритмы для решения сложных инженерных задач. После работ Джона Холланда стали популярными генетические алгоритмы. Практические приложения также включают автоматическую эволюцию компьютерных программ. Эволюционные алгоритмы ныне используются для решения многомерных проблем, а также для оптимизации проектирования систем[239].
Отражение в общественной жизни
 Карикатура на Чарльза Дарвина. 1871 год
Карикатура на Чарльза Дарвина. 1871 годВ XIX веке, особенно после публикации в 1859 году «Происхождения видов», идея того, что жизнь развивалась, а не была создана в «готовом» виде, была предметом научных и общественных дискуссий. В основном данные дискуссии фокусировались на философских, социальных аспектах и взаимоотношении эволюционной теории с религией. В настоящее время факт эволюции не вызывает сомнений у подавляющего числа учёных, а наиболее общепринятой теорией, объясняющей механизмы эволюционного процесса, является синтетическая теория эволюции[1]. Несмотря на это, факт эволюции не признаётся некоторыми верующими.
В настоящее время различные религиозные конфессии признали возможность сочетания религиозных верований с эволюцией через такие концепции, как теистический эволюционизм. Однако есть и креационисты, которые считают, что эволюция противоречит религиозным представлениям о происхождении мира и живых существ, которые есть в их религиях, и в связи с этим ставят под сомнение реальность эволюции[123][240][241]. Самое большое количество споров между религиозными и научными взглядами вызывает такой аспект эволюционной биологии, как происхождение человека и его родственные отношения с обезьянами, а также то, что возникновение умственных способностей и моральных качеств у людей имело естественные причины. В некоторых странах, особенно в США, эти противоречия между наукой и религией подпитывали эволюционно-креационные дебаты (англ.)русск., конфликт, фокусирующийся на политике и общественном образовании[242]. Хотя данные многих научных областей, таких как космология и геология, противоречат буквальному толкованию религиозных текстов, именно эволюционная биология больше всего входит в противоречие со взглядами сторонников религиозного буквализма.
Часть из креационистских антиэволюционных концепций, такие как «научный креационизм» и появившаяся в середине 1990-х годов нео-креационистская концепция «Разумного замысла» (англ. Intelligent design), утверждают, что имеют научное основание. Однако научным сообществом они признаны псевдонаучными концепциями, поскольку не соответствуют критериям верифицируемости, фальсифицируемости и принципу Оккама, а также противоречат научным данным[243].
См. также
- Эволюционная биология
- Доказательства эволюции
- Критика эволюционизма
- Антропогенез
- Социальная эволюция
Комментарии
- ↑ Однако следует иметь в виду, что отсутствие долгосрочных целей или планов не означает, что у процесса эволюции нет закономерностей, а также не исключает возможность канализированности эволюции, например, за счёт особенностей морфофункциональной организации или условий окружающей среды. См., например, Северцов А. С. Теория эволюции. — М.: Владос, 2005. — С. 354—358. — 380 с. — 10 000 экз.
Примечания
- ↑ 1 2 3 4 Kutschera U, Niklas KJ The modern theory of biological evolution: an expanded synthesis (англ.) // Naturwissenschaften. — 2004. — Т. 91. — № 6. — С. 255-276.
- ↑ Futuyma, 2005, с. 2
- ↑ National Academy of Science Institute of Medicine. Science, Evolution, and Creationism. — National Academy Press, 2008. — ISBN 0309105862
- ↑ Futuyma, Douglas. Evolution, Science, and Society: Evolutionary Biology and the National Research Agenda. — Office of University Publications, Rutgers, The State University of New Jersey, 1999. — P. 46.
- ↑ Воронцов, 1999, с. 118-119
- ↑ Воронцов, 1999, с. 124-125
- ↑ Воронцов, 1999, с. 127
- ↑ Torrey H. B., Felin F. Was Aristotle an evolutionist? // The Quarterly Review of Biology. — 1937. — Т. 12. — № 1. — С. 1-18.
- ↑ Hull D. L. The metaphysics of evolution // The British Journal for the History of Science. — 1967. — Т. 3. — № 4. — С. 309-337.
- ↑ Stephen F. Mason. A history of the sciences. — Collier Books, 1968. — С. 44-45. — 638 с.
- ↑ Воронцов, 1999, с. 171-172
- ↑ Ernst Mayr The growth of biological thought: diversity, evolution, and inheritance. — Harvard University Press, 1982. — С. 256-257. — ISBN 0674364465
- ↑ Carl Linnaeus (1707-1778) (англ.). University of California Museum of Paleontology. Архивировано из первоисточника 21 июня 2012. Проверено 29 февраля 2012.
- ↑ Воронцов, 1999, с. 181-183
- ↑ Peter J. Bowler. Evolution: The History of an Idea. — University of California Press, 1989. — С. 71-72. — 432 с. — ISBN 0520236939
- ↑ Erasmus Darwin (1731-1802) (англ.). University of California Museum of Paleontology. Архивировано из первоисточника 21 июня 2012. Проверено 29 февраля 2012.
- ↑ Воронцов, 1999, с. 201-209
- ↑ Gould, 2002, с. 170-189
- ↑ Воронцов, 1999, с. 210-217
- ↑ Peter J. Bowler. Evolution: The History of an Idea. — University of California Press, 1989. — С. 145-146. — 432 с. — ISBN 0520236939
- ↑ Mayr, 2001, с. 165
- ↑ Воронцов, 1999, с. 278-279
- ↑ Воронцов, 1999, с. 282-283
- ↑ Воронцов, 1999, с. 283
- ↑ Stamhuis IH, Meijer OG, Zevenhuizen EJ. Hugo de Vries on heredity, 1889-1903. Statistics, Mendelian laws, pangenes, mutations // Isis. — 1999. — Т. 90. — № 2. — С. 238-267.
- ↑ Воронцов, 1999, с. 405-407
- ↑ Dobzhansky, T. Nothing in biology makes sense except in the light of evolution // The American Biology Teacher. — 1973. — Т. 35. — № 3. — С. 125–129.
- ↑ Avise, J. C.; Ayala, F. J. In the Light of Evolution IV. The Human Condition (introduction) // Proceedings of the National Academy of Sciences USA. — 2010. — Т. 107. — С. 8897–8901.
- ↑ 1 2 Кимура М. Молекулярная эволюция: теория нейтральности: Пер. с англ. — М.: Мир, 1985. — 394 с.
- ↑ Eldredge N., Gould S. J. Punctuated equilibria: an alternative to phyletic gradualism // Models in paleobiology / Schopf T. J. M., ed. — San Francisco: Freeman Cooper, 1972. — P. 82—115.
- ↑ West-Eberhard M.-J. Developmental plasticity and evolution. — New York: Oxford University Press, 2003. — ISBN 978-0-19-512235-0
- ↑ Palmer R. A. Symmetry breaking and the evolution of development // Science. — 2004. — Т. 306. — С. 828–833. — DOI:10.1126/science.1103707 — Bibcode: 2004Sci...306..828P — PMID 15514148.
- ↑ Солдбриг, 1982, с. 78-80
- ↑ Солдбриг, 1982, с. 125-126
- ↑ Грант, 1991
- ↑ Глазер В. М. Конверсия гена
- ↑ Sturm RA, Frudakis TN Eye colour: portals into pigmentation genes and ancestry // Trends Genet. — 2004. — Т. 20(8). — С. 327-332.
- ↑ 1 2 Pearson H Genetics: what is a gene? // Nature. — 2006. — Т. 441 (7092). — С. 398—401.
- ↑ Patrick C. Phillips Epistasis—the essential role of gene interactions in the structure and evolution of genetic systems // Nat Rev Genet. — 2008. — № 9(11). — С. 855—867.
- ↑ 1 2 Wu R, Lin M Functional mapping — how to map and study the genetic architecture of dynamic complex traits // Nat. Rev. Genet. — 2006. — Т. 7 (3). — С. 229-237.
- ↑ Jablonka E., Raz G. Transgenerational epigenetic inheritance: Prevalence, mechanisms and implications for the study of heredity and evolution // The Quarterly Review of Biology. — 2009. — Т. 84. — № 2. — С. 131–176.
- ↑ Bossdorf O., Arcuri D., Richards C. L., Pigliucci M. Experimental alteration of DNA methylation affects the phenotypic plasticity of ecologically relevant traits in Arabidopsis thaliana // Evolutionary Ecology. — 2010. — Т. 24. — № 3. — С. 541–553.
- ↑ Jablonka E., Lamb M. Evolution in four dimensions: Genetic, epigenetic, behavioural and symbolic. — MIT Press, 2005.
- ↑ Laland K. N., Sterelny K. Perspective: Seven reasons (not) to neglect niche construction // Evolution. — 2006. — Т. 60. — № 8. — С. 1751–1762.
- ↑ Chapman M. J., Margulis L. Morphogenesis by symbiogenesis // International Microbiology. — 1998. — Т. 1. — № 4. — С. 319–326.
- ↑ Wilson D. S., Wilson E. O. Rethinking the theoretical foundation of sociobiology // The Quarterly Review of Biology. — 2007. — Т. 82. — № 4.
- ↑ Северцов, 2005
- ↑ AmosW, Harwood J Factors affecting levels of genetic diversity in natural populations // Philos Trans R Soc Lond B Biol Sci. — 1998. — № 353(1366). — С. 177—186.
- ↑ Draghi J, Turner P DNA secretion and gene-level selection in bacteria // Microbiology (Reading, Engl.). — 2008. — Т. 152. — С. 2683-26888.
- ↑ Mallet J Hybrid speciation // Nature. — 2007. — Т. 446 (7133). — С. 279-283.
- ↑ Butlin RK, Tregenza T Levels of genetic polymorphism: marker loci versus quantitative traits // Philos. Trans. R. Soc. Lond., B, Biol. Sci. — 1998. — Т. 353 (1366). — С. 187-198.
- ↑ Wetterbom A, Sevov M, Cavelier L, Bergström TF Comparative genomic analysis of human and chimpanzee indicates a key role for indels in primate evolution // J. Mol. Evol. — 2006. — Т. 63 (5). — С. 682-690.
- ↑ 1 2 Bertram JS The molecular biology of cancer // Mol Aspects Med. — 2000. — Т. 21(6). — С. 167-223.
- ↑ 1 2 Aminetzach YT, Macpherson JM, Petrov DA Pesticide resistance via transposition-mediated adaptive gene truncation in Drosophila // Science. — 2005. — Т. 309(5735). — С. 764-767.
- ↑ Burrus V, Waldor MK Shaping bacterial genomes with integrative and conjugative elements // Res Microbiol. — 2004. — Т. 155(5). — С. 376-386.
- ↑ Sawyer SA, Parsch J, Zhang Z, Hartl DL Prevalence of positive selection among nearly neutral amino acid replacements in Drosophila // Proc Natl Acad Sci U S A. — 2007. — Т. 104(16). — С. 6504-6510.
- ↑ Sniegowski PD, Gerrish PJ, Johnson T, Shaver A The evolution of mutation rates: separating causes from consequences // Bioessays. — 2000. — Т. 22(12). — С. 1057-1066.
- ↑ Drake JW, Holland JJ Mutation rates among RNA viruses // Proc Natl Acad Sci U S A. — 1999. — Т. 96(24). — С. 13910-13913.
- ↑ Holland J, Spindler K, Horodyski F, Grabau E, Nichol S, VandePol S Rapid evolution of RNA genomes // Science. — 1982. — Т. 215(4540). — С. 1577-1585.
- ↑ Hastings PJ, Lupski JR, Rosenberg SM, Ira G Mechanisms of change in gene copy number // Nat Rev Genet. — 2009. — Т. 10(8). — С. 551-564.
- ↑ Harrison PM, Gerstein M Studying genomes through the aeons: protein families, pseudogenes and proteome evolution // J Mol Biol. — 2002. — Т. 318(5). — С. 1155-1174.
- ↑ Bowmaker JK Evolution of colour vision in vertebrates // Eye (Lond). — 1998. — Т. 12. — С. 541-547.
- ↑ Gregory TR, Hebert PD The modulation of DNA content: proximate causes and ultimate consequences // Genome Res. — 1999. — Т. 9(4). — С. 317-324.
- ↑ Hurles M Gene duplication: the genomic trade in spare parts // PLoS Biol. — 2004. — Т. 2(7). — С. E206.
- ↑ Adam Siepel Darwinian alchemy: Human genes from noncoding DNA // Genome Res. — 2009. — Т. 19. — С. 1693—1695.
- ↑ Liu N, Okamura K, Tyler DM, Phillips MD, Chung WJ, Lai EC The evolution and functional diversification of animal microRNA genes // Cell Res. — 2008. — Т. 18(10). — С. 985-996.
- ↑ Orengo CA, Thornton JM Protein families and their evolution-a structural perspective // Annu Rev Biochem. — 2005. — Т. 74. — С. 867-900.
- ↑ Long M, Betrán E, Thornton K, Wang W The origin of new genes: glimpses from the young and old // Nat Rev Genet. — 2003. — Т. 4(11). — С. 865-875.
- ↑ Weissman KJ, Müller R Protein-protein interactions in multienzyme megasynthetases // Chembiochem. — 2008. — Т. 9(6). — С. 826-848.
- ↑ Zhang J, Wang X, Podlaha O Testing the chromosomal speciation hypothesis for humans and chimpanzees // Genome Res. — 2004. — Т. 14(5). — С. 845-851.
- ↑ Ayala FJ, Coluzzi M Chromosome speciation: humans, Drosophila, and mosquitoes // Proc Natl Acad Sci U S A. — Т. 102. — С. 6535-6542.
- ↑ Hurst GD, Werren JH The role of selfish genetic elements in eukaryotic evolution // Nat Rev Genet. — 2001. — Т. 2(8). — С. 597-606.
- ↑ Häsler J, Strub K Alu elements as regulators of gene expression // Nucleic Acids Res. — 2006. — Т. 34(19). — С. 5491-5497.
- ↑ Radding, C. M. Homologous pairing and strand exchange in genetic recombination (англ.) // Annu Rev Genet. — 1982. — Т. 16. — P. 405-437.
- ↑ Agrawal AF Evolution of sex: why do organisms shuffle their genotypes? (англ.) // Curr Biol. — 2006. — В. 17. — Т. 16. — P. 696-704.
- ↑ Goddard MR, Godfray HC, Burt A Sex increases the efficacy of natural selection in experimental yeast populations (англ.) // Nature. — 2005. — Т. 434. — P. 636-640.
- ↑ Peters AD, Otto SP Liberating genetic variance through sex (англ.) // Bioessays. — 2003. — В. 6. — Т. 25. — P. 533-537.
- ↑ Fontaneto D, Herniou EA, Boschetti C, Caprioli M, Melone G, Ricci C, Barraclough TG Independently evolving species in asexual bdelloid rotifers (англ.) // PLoS Biol. — 2007. — В. 4. — Т. 5. — P. 87.
- ↑ Lien S, Szyda J, Schechinger B, Rappold G, Arnheim N Evidence for heterogeneity in recombination in the human pseudoautosomal region: high resolution analysis by sperm typing and radiation-hybrid mapping (англ.) // Am J Hum Genet. — 2000. — В. 2. — Т. 66. — P. 557-566.
- ↑ Barton NH Genetic hitchhiking (англ.) // Philos Trans R Soc Lond B Biol Sci. — 2000. — Т. 355. — P. 1553-1562.
- ↑ Charlesworth B, Charlesworth D The degeneration of Y chromosomes (англ.) // Philos Trans R Soc Lond B Biol Sci. — 200. — Т. 355. — P. 1563-1572.
- ↑ Otto SP The advantages of segregation and the evolution of sex (англ.) // Genetics. — 2003. — В. 3. — Т. 164. — P. 1099-118.
- ↑ Doncaster CP, Pound GE, Cox SJ The ecological cost of sex (англ.) // Nature. — 2000. — Т. 404. — P. 281-285.
- ↑ Butlin R Evolution of sex: The costs and benefits of sex: new insights from old asexual lineages (англ.) // Nat Rev Genet. — 2002. — В. 4. — Т. 3. — P. 311-317.
- ↑ Salathé M, Kouyos RD, Bonhoeffer S The state of affairs in the kingdom of the Red Queen (англ.) // Trends Ecol Evol. — 2008. — В. 8. — Т. 23. — P. 439-445.
- ↑ 1 2 Грант, 1991, Глава 17
- ↑ Грант, 1991, Глава 23
- ↑ Солдбриг, 1982, с. 43-47
- ↑ Futuyma, 2005, p. 251-252
- ↑ 1 2 3 Orr HA Fitness and its role in evolutionary genetics // Nat Rev Genet. — 2009. — Vol. 10(8). — P. 531-539.
- ↑ Haldane J The theory of natural selection today // Nature. — 1959. — Vol. 183. — P. 710-713.
- ↑ Lande R, Arnold SJ The measurement of selection on correlated characters // Evolution. — 1983. — Vol. 37. — P. 1210–26}. — DOI:10.2307/2408842
- ↑ Futuyma, 2005
- ↑ Грант, 1991, Глава 14
- ↑ Andersson M, Simmons L Sexual selection and mate choice // Trends Ecol Evol. — 2001. — Vol. 21(6). — P. 296-302.
- ↑ Kokko H, Brooks R, McNamara J, Houston A The sexual selection continuum // Proc Biol Sci. — 2002. — Vol. 269. — P. 1331-1340.
- ↑ Hunt J, Brooks R, Jennions MD, Smith MJ, Bentsen CL, Bussière LF High-quality male field crickets invest heavily in sexual display but die young // Nature. — 2004. — Vol. 432. — P. 1024-1027.
- ↑ 1 2 Gould SJ Gulliver's further travels: the necessity and difficulty of a hierarchical theory of selection // Philos. Trans. R. Soc. Lond., B, Biol. Sci. — 1998. — Т. 353. — С. 307–314.
- ↑ Okasha, S. Evolution and the Levels of Selection. — Oxford University Press, 2007. — 263 с. — ISBN 0-19-926797-9
- ↑ Mayr E The objects of selection // Philos. Trans. R. Soc. Lond., B, Biol. Sci. — 1998. — Т. 353. — С. 307–14.
- ↑ Maynard Smith J The units of selection // Novartis Found. Symp. — 1998. — Т. 213. — С. 211–217.
- ↑ Gould SJ, Lloyd EA Individuality and adaptation across levels of selection: how shall we name and generalise the unit of Darwinism? // Proc. Natl. Acad. Sci. U.S.A. — 1999. — Т. 96. — № 21. — С. 11904–11909.
- ↑ 1 2 Masel J Genetic drift // Current Biology. — 2011. — Т. 21. — № 20. — С. R837-R838.
- ↑ Lande R Fisherian and Wrightian theories of speciation // Genome. — 1989. — Т. 31. — № 1. — С. 221-227.
- ↑ Otto SP, Whitlock MC The probability of fixation in populations of changing size // Genetics. — 1997. — Т. 146. — № 2. — С. 723-733.
- ↑ 1 2 Charlesworth B Fundamental concepts in genetics: effective population size and patterns of molecular evolution and variation // Nat Rev Genet. — 2009. — Т. 10. — № 3. — С. 195-205.
- ↑ Mitchell-Olds T, Willis JH, Goldstein DB Which evolutionary processes influence natural genetic variation for phenotypic traits? // Nat Rev Genet. — 2007. — Т. 8. — № 11. — С. 845-856.
- ↑ Nei M Selectionism and neutralism in molecular evolution // Mol Biol Evol. — 2005. — Т. 22. — № 12. — С. 2318-2342.
- ↑ Kimura M The neutral theory of molecular evolution: a review of recent evidence // Jpn J Genet. — 1991. — Т. 66. — № 4. — С. 367-386.
- ↑ Kimura M The neutral theory of molecular evolution and the world view of the neutralists // Genome. — 1989. — Т. 31. — № 1. — С. 24-31.
- ↑ Kreitman M The neutral theory is dead. Long live the neutral theory // Bioessays. — 1996. — Т. 18. — № 8. — С. 678–683.
- ↑ Leigh E.G. (Jr) Neutral theory: a historical perspective // Journal of Evolutionary Biology. — 2007. — Т. 20. — № 6. — С. 2075–2091.
- ↑ Hurst LD Fundamental concepts in genetics: genetics and the understanding of selection // Nat. Rev. Genet. — 2009. — Т. 10. — № 2. — С. 83–93.
- ↑ Gillespie, John H. Is the population size of a species relevant to its evolution? // Evolution. — 2001. — Т. 55. — № 11. — С. 2161–2169.
- ↑ R. A. Neher and B. I. Shraiman Genetic Draft and Quasi-Neutrality in Large Facultatively Sexual Populations // Genetics. — 2011. — Т. 188. — С. 975–996.
- ↑ Boucher Y, Douady CJ, Papke RT, Walsh DA, Boudreau ME, Nesbo CL, Case RJ, Doolittle WF Lateral gene transfer and the origins of prokaryotic groups // Annu Rev Genet. — 2003. — Т. 37. — № 1. — С. 283–328.
- ↑ Walsh T Combinatorial genetic evolution of multiresistance // Curr. Opin. Microbiol. — 2006. — Т. 9. — № 5. — С. 476–82.
- ↑ Kondo N, Nikoh N, Ijichi N, Shimada M, Fukatsu T Genome fragment of Wolbachia endosymbiont transferred to X chromosome of host insect // Proc. Natl. Acad. Sci. U.S.A. — 2002. — Т. 99. — № 22. — С. 14280–5.
- ↑ Sprague G Genetic exchange between kingdoms // Curr. Opin. Genet. Dev. — 1991. — Т. 1. — № 4. — С. 530–3.
- ↑ Gladyshev EA, Meselson M, Arkhipova IR Massive horizontal gene transfer in bdelloid rotifers // Science. — 2008. — Т. 320. — № 5880. — С. 1210–3.
- ↑ Baldo A, McClure M Evolution and horizontal transfer of dUTPase-encoding genes in viruses and their hosts // J. Virol. — Т. 73. — № 9. — С. 7710–21.
- ↑ River, M. C. and Lake, J. A. The ring of life provides evidence for a genome fusion origin of eukaryotes // Nature. — 2004. — Т. 431. — № 9. — С. 152–5.
- ↑ 1 2 3 Scott EC, Matzke NJ Biological design in science classrooms // Proc Natl Acad Sci U S A. — 207. — Т. 104. — С. 8669-8676.
- ↑ Hendry AP, Kinnison MT An introduction to microevolution: rate, pattern, process // Genetica. — 2001. — Т. 112–113. — С. 1–8.
- ↑ Leroi AM The scale independence of evolution // Evol. Dev. — 2000. — Т. 2. — С. 67–77.
- ↑ Michael J. Dougherty Is the human race evolving or devolving?. Scientific American (1998). Архивировано из первоисточника 21 июня 2012. Проверено 30 января 2012.
- ↑ response to Creationist claims – Claim CB932: Evolution of degenerate forms. TalkOrigins Archive. Архивировано из первоисточника 21 июня 2012. Проверено 30 января 2012.
- ↑ Futuyma, 2005, с. 250-251
- ↑ Mayr, 2001, с. 133-134, 235-238
- ↑ Carroll SB Chance and necessity: the evolution of morphological complexity and diversity // Nature. — 2001. — Т. 409. — С. 1102–1109.
- ↑ Whitman W, Coleman D, Wiebe W Prokaryotes: the unseen majority // Proc Natl Acad Sci U S A. — 1999. — Т. 95. — № 12. — С. 6578–6583.
- ↑ Schloss P, Handelsman J Status of the microbial census // Microbiol Mol Biol Rev. — 2004. — Т. 68. — № 4. — С. 686–691.
- ↑ 1 2 Bejder L, Hall BK Limbs in whales and limblessness in other vertebrates: mechanisms of evolutionary and developmental transformation and loss // Evol. Dev. — 2002. — Т. 4. — № 6. — С. 445–58.
- ↑ Orr H The genetic theory of adaptation: a brief history // Nat. Rev. Genet. — 2005. — Т. 6. — № 2. — С. 119–27.
- ↑ Dobzhansky T. Genetics of natural populations XXV. Genetic changes in populations of Drosophila pseudoobscura and Drosphila persimilis in some locations in California // Evolution. — 1956. — Т. 10. — № 1. — С. 82–92.
- ↑ Nakajima A, Sugimoto Y, Yoneyama H, Nakae T High-level fluoroquinolone resistance in Pseudomonas aeruginosa due to interplay of the MexAB-OprM efflux pump and the DNA gyrase mutation // Microbiol. Immunol. — 2002. — Т. 46. — № 6. — С. 391–5.
- ↑ Blount ZD, Borland CZ, Lenski RE Inaugural Article: Historical contingency and the evolution of a key innovation in an experimental population of Escherichia coli // Proc. Natl. Acad. Sci. U.S.A. — 2008. — Т. 105. — № 23. — С. 7899–906.
- ↑ Okada H, Negoro S, Kimura H, Nakamura S Evolutionary adaptation of plasmid-encoded enzymes for degrading nylon oligomers // Nature. — 1983. — Т. 306. — № 5939. — С. 203–6.
- ↑ Ohno S Birth of a unique enzyme from an alternative reading frame of the preexisted, internally repetitious coding sequence // Proc. Natl. Acad. Sci. U.S.A. — 1984. — Т. 81. — № 8. — С. 2421–5.
- ↑ Copley SD Evolution of a metabolic pathway for degradation of a toxic xenobiotic: the patchwork approach // Trends Biochem. Sci. — 2000. — Т. 25. — № 6. — С. 261–5.
- ↑ Crawford RL, Jung CM, Strap JL The recent evolution of pentachlorophenol (PCP)-4-monooxygenase (PcpB) and associated pathways for bacterial degradation of PCP // Biodegradation. — 2007. — Т. 18. — № 5. — С. 525–39.
- ↑ Eshel I Clone-selection and optimal rates of mutation // Journal of Applied Probability. — 1973. — Т. 10. — № 4. — С. 728–738. — DOI:10.2307/3212376
- ↑ Masel J, Bergman A, The evolution of the evolvability properties of the yeast prion [PSI+] // Evolution. — 2003. — Т. 57. — № 7. — С. 1498–1512.
- ↑ Lancaster AK, Bardill JP, True HL, Masel J The Spontaneous Appearance Rate of the Yeast Prion [PSI+ and Its Implications for the Evolution of the Evolvability Properties of the [PSI+] System] // Genetics. — 2010. — Т. 184. — № 2. — С. 393–400.
- ↑ Draghi J, Wagner G Evolution of evolvability in a developmental model // Theoretical Population Biology. — 2008. — Т. 62. — С. 301–315.
- ↑ Young NM., Hallgrímsson B. Serial homology and the evolution of mammalian limb covariation structure // Evolution. — 2005. — Т. 59. — № 12. — С. 2691–704.
- ↑ 1 2 Penny D, Poole A. The nature of the last universal common ancestor (англ.) // Curr Opin Genet Dev. — 1999. — В. 6. — Т. 9. — P. 672-677.
- ↑ Hall BK Descent with modification: the unity underlying homology and homoplasy as seen through an analysis of development and evolution // Biological Reviews of the Cambridge Philosophical Society. — 2003. — Т. 78. — № 3. — С. 409–433.
- ↑ Shubin N, Tabin С, Carroll C Deep homology and the origins of evolutionary novelty // Nature. — 2009. — Т. 457. — № 7231. — С. 818–823.
- ↑ 1 2 3 Fong D, Kane T, Culver D Vestigialization and Loss of Nonfunctional Characters // Ann. Rev. Ecol. Syst. — 1995. — Т. 26. — № 4. — С. 249–68.
- ↑ Zhang Z, Gerstein M Large-scale analysis of pseudogenes in the human genome // Curr. Opin. Genet. Dev. — 2004. — Т. 14. — № 4. — С. 328–35.
- ↑ Jeffery WR Adaptive evolution of eye degeneration in the Mexican blind cavefish // J. Hered. — 2005. — Т. 96. — № 3. — С. 185–96.
- ↑ Maxwell EE, Larsson HC Osteology and myology of the wing of the Emu (Dromaius novaehollandiae) and its bearing on the evolution of vestigial structures // J. Morphol. — 2007. — Т. 268. — № 5. — С. 423–41.
- ↑ Silvestri AR, Singh I The unresolved problem of the third molar: would people be better off without it? // Journal of the American Dental Association (1939). — 2003. — Т. 134. — № 4. — С. 450–5.
- ↑ Coyne, Jerry A. Why Evolution is True. — Oxford University Press, 2009. — С. 66. — 309 с. — ISBN 9780670020539
- ↑ Coyne, Jerry A. Why Evolution is True. — Oxford University Press, 2009. — С. 85-86. — 309 с. — ISBN 9780670020539
- ↑ Gould, 2002
- ↑ Pallen MJ., Matzke NJ From The Origin of Species to the origin of bacterial flagella // Nat Rev Micro. — Т. 4. — № 10. — С. 784–790. — PMID http://www.ncbi.nlm.nih.gov/pubmed/16953248.
- ↑ Clements A, Bursac D., Gatsos X., Perry AJ., Civciristov S., Celik N., Likic VA., Poggio S., Jacobs-Wagner S. The reducible complexity of a mitochondrial molecular machine // Proceedings of the National Academy of Sciences. — 2009. — Т. 106. — № 37. — С. 15791–15795.
- ↑ Hardin G The competitive exclusion principle // Science. — 1960. — Т. 131. — № 3409. — С. 1292–7.
- ↑ Wade MJ The co-evolutionary genetics of ecological communities // Nat Rev Genet. — 2007. — Т. 8. — № 3. — С. 185-195.
- ↑ Brodie ED Jr, Ridenhour BJ, Brodie ED 3rd. The evolutionary response of predators to dangerous prey: hotspots and coldspots in the geographic mosaic of coevolution between garter snakes and newts // Evolution. — 2002. — Т. 56. — № 10. — С. 2067-2082.
- ↑ Sachs JL Cooperation within and among species // J Evol Biol. — 2006. — Т. 19. — № 5. — С. 1426-1436.
- ↑ Nowak MA Five rules for the evolution of cooperation // Science. — 2006. — Т. 314. — С. 1560-1563.
- ↑ Paszkowski U Mutualism and parasitism: the yin and yang of plant symbioses // Curr Opin Plant Biol. — 2006. — Т. 9. — № 4. — С. 364-370.
- ↑ Hause B, Fester T Molecular and cell biology of arbuscular mycorrhizal symbiosis // Planta. — 2005. — Т. 221. — № 2. — С. 184-196.
- ↑ Reeve HK, Hölldobler B The emergence of a superorganism through intergroup competition // Proc Natl Acad Sci U S A. — 2007. — Т. 104. — С. 9736-9740.
- ↑ Axelrod R, Hamilton WD The evolution of cooperation // Science. — 1981. — Т. 211. — С. 1390-1396.
- ↑ Wilson EO, Hölldobler B Eusociality: origin and consequences // Proc Natl Acad Sci U S A. — 2005. — Т. 102. — С. 13367-13371.
- ↑ Joseph Boxhorn Observed Instances of Speciation (англ.). TalkOrigins Archive. Архивировано из первоисточника 21 июня 2012. Проверено 23 сентября 2011.
- ↑ Jiggins CD, Bridle JR Speciation in the apple maggot fly: a blend of vintages? // Trends Ecol Evol. — 2004. — Т. 19. — № 3. — С. 11-114.
- ↑ Weinberg JR, Starczak VR, Jorg, D Evidence for Rapid Speciation Following a Founder Event in the Laboratory // Evolution. — 1992. — Т. 46. — № 4. — С. 1214-1220.
- ↑ de Queiroz K Ernst Mayr and the modern concept of species // Proc Natl Acad Sci U S A. — 2005. — Т. 102. — С. Suppl 1:6600-6607.
- ↑ Ereshefsky, M Eliminative pluralism // Philosophy of Science. — 1992. — Т. 59. — № 4. — С. 671–690.
- ↑ Майр, Эрнест. Систематика и происхождение видов с точки зрения зоолога. — М.: государственное из-во иностранной литературы, 1942. — С. 194. — 505 с.
- ↑ Fraser C, Alm EJ, Polz MF, Spratt BG, Hanage WP The bacterial species challenge: making sense of genetic and ecological diversity // Science. — 2009. — Т. 323. — С. 741-746.
- ↑ Gross BL, Rieseberg LH The ecological genetics of homoploid hybrid speciation // J Hered. — 2005. — Т. 96. — № 3. — С. 241-252.
- ↑ Burke JM, Arnold ML Genetics and the fitness of hybrids // Annu Rev Genet. — 2001. — Т. 35. — С. 31-52.
- ↑ Vrijenhoek RC Polyploid hybrids: multiple origins of a treefrog species // Curr Biol. — 2006. — Т. 16. — № 7. — С. R245-247.
- ↑ Herrel A, Huyghe K, Vanhooydonck B, Backeljau T, Breugelmans K, Grbac I, Van Damme R, Irschick DJ Rapid large-scale evolutionary divergence in morphology and performance associated with exploitation of a different dietary resource // Proc Natl Acad Sci U S A. — 2008. — Т. 105. — № 12. — С. 4792-4795.
- ↑ Losos, J. B. Warhelt, K. I. Schoener, T. W Adaptive differentiation following experimental island colonization in Anolis lizards // Nature. — 1997. — Т. 387. — С. 70-73.
- ↑ Hoskin CJ, Higgie M, McDonald KR, Moritz C Reinforcement drives rapid allopatric speciation // Nature. — 2005. — Т. 437. — С. 1353-1356.
- ↑ Templeton AR The theory of speciation via the founder principle // Genetics. — 1980. — Т. 94. — № 4. — С. 1011-1038.
- ↑ Gavrilets S Perspective: models of speciation: what have we learned in 40 years? // Evolution. — 2003. — Т. 57. — № 10. — С. 2197-2215.
- ↑ Antonovics J Evolution in closely adjacent plant populations X: long-term persistence of prereproductive isolation at a mine boundary // Heredity. — 2006. — Т. 97. — № 1. — С. 33-37.
- ↑ Nosil P, Crespi BJ, Gries R, Gries G Natural selection and divergence in mate preference during speciation // Genetica. — 2007. — Т. 129. — № 3. — С. 309-327.
- ↑ Savolainen V, Anstett MC, Lexer C, Hutton I, Clarkson JJ, Norup MV, Powell MP, Springate D, Salamin N, Baker WJ Sympatric speciation in palms on an oceanic island // Nature. — 2006. — Т. 441. — С. 210-213.
- ↑ Barluenga M, Stölting KN, Salzburger W, Muschick M, Meyer A Sympatric speciation in Nicaraguan crater lake cichlid fish // Nature. — 2006. — Т. 439. — С. 719-723.
- ↑ Gavrilets S The Maynard Smith model of sympatric speciation // J Theor Biol. — 2006. — Т. 239. — № 2. — С. 172-182.
- ↑ Wood TE, Takebayashi N, Barker MS, Mayrose I, Greenspoon PB, Rieseberg LH The frequency of polyploid speciation in vascular plants // Proc Natl Acad Sci U S A. — 2009. — Т. 106. — С. 13875-13879.
- ↑ Hegarty MJ, Hiscock SJ Genomic clues to the evolutionary success of polyploid plants // Curr Biol. — 2008. — Т. 18. — № 10. — С. R435-444.
- ↑ Jakobsson M, Hagenblad J, Tavaré S, Säll T, Halldén C, Lind-Halldén C, Nordborg M A unique recent origin of the allotetraploid species Arabidopsis suecica: Evidence from nuclear DNA markers // Mol Biol Evol. — 2006. — Т. 23. — № 6. — С. 1217-1231.
- ↑ Säll T, Jakobsson M, Lind-Halldén C, Halldén C Chloroplast DNA indicates a single origin of the allotetraploid Arabidopsis suecica // J Evol Biol. — 2003. — Т. 16. — № 5. — С. 1019-1029.
- ↑ Bomblies K, Weigel D Arabidopsis: a model genus for speciation // Curr Opin Genet Dev. — 2007. — Т. 17. — № 6. — С. 500-504.
- ↑ Benton MJ Diversification and extinction in the history of life // Science. — 1995. — Т. 268. — С. 52-58.
- ↑ Raup DM Biological extinction in Earth history // Science. — 1986. — Т. 231. — С. 1528–1533.
- ↑ Avise JC, Hubbell SP, Ayala FJ In the light of evolution II: Biodiversity and extinction // Proc. Natl. Acad. Sci. U.S.A. — 2008. — Т. 105. — С. 11453–11457.
- ↑ 1 2 Raup DM The role of extinction in evolution // Proc. Natl. Acad. Sci. U.S.A. — 1994. — Т. 91. — С. 6758–6763.
- ↑ Novacek MJ, Cleland EE The current biodiversity extinction event: scenarios for mitigation and recovery // Proc. Natl. Acad. Sci. U.S.A. — 2001. — Т. 98. — № 10. — С. 5466–5470.
- ↑ Pimm S, Raven P, Peterson A, Sekercioglu CH, Ehrlich PR Human impacts on the rates of recent, present, and future bird extinctions. — 2006. — Т. 103. — № 29. — С. 10941-10946.
- ↑ Barnosky AD, Koch PL, Feranec RS, Wing SL, Shabel AB Assessing the causes of late Pleistocene extinctions on the continents // Science. — 2004. — Т. 306. — С. 70-75.
- ↑ Lewis OT Climate change, species-area curves and the extinction crisis // hilos. Trans. R. Soc. Lond., B, Biol. Sci. — 2006. — Т. 361. — С. 163-171.
- ↑ Jablonski D Lessons from the past: evolutionary impacts of mass extinctions // Proc. Natl. Acad. Sci. U.S.A. — 2001. — Т. 98. — С. 5393–5398.
- ↑ Mark Isaak. Index to Creationist Claims.
- ↑ Peretó J. Controversies on the origin of life. — Int Microbiol., 2005. — Т. 8(1). — С. 23-31.
- ↑ Luisi PL, Ferri F, Stano P. Approaches to semi-synthetic minimal cells: a review. — Naturwissenschaften, 2006. — Т. 93(1). — С. 1-13.
- ↑ Joyce GF. The antiquity of RNA-based evolution. — Nature., 2002. — Т. 418. — С. 214-221.
- ↑ Bapteste E, Walsh DA. Does the 'Ring of Life' ring true? (англ.) // Trends Microbiol. — 2005. — Т. 13. — P. 256-261.
- ↑ Darwin, Charles (1859). On the Origin of Species
- ↑ Doolittle WF, Bapteste E. Pattern pluralism and the Tree of Life hypothesis (англ.) // Proc Natl Acad Sci U S A. — 2007. — Т. 104. — P. 2043-2049.
- ↑ Kunin V, Goldovsky L, Darzentas N, Ouzounis CA. The net of life: reconstructing the microbial phylogenetic network (англ.) // Genome Res. — 2005. — Т. 15. — P. 954-959.
- ↑ Jablonski D. The future of the fossil record (англ.) // Science. — 1999. — Т. 284. — P. 2114-2116.
- ↑ Mason SF. Origins of biomolecular handedness (англ.) // Nature. — 1984. — Т. 311. — P. 19-23.
- ↑ Wolf YI, Rogozin IB, Grishin NV, Koonin EV. Genome trees and the tree of life (англ.) // Trends Genet. — 2002. — Т. 18. — P. 472-479.
- ↑ Varki A, Altheide TK. Comparing the human and chimpanzee genomes: searching for needles in a haystack (англ.) // Genome Res. — 2005. — Т. 15. — P. 1746-1758.
- ↑ Futuyma, 2005, p. 94
- ↑ Futuyma, 2005, p. 96
- ↑ Марков, 2010, p. 135-169
- ↑ Cavalier-Smith T Cell evolution and Earth history: stasis and revolution // Philos Trans R Soc Lond B Biol Sci. — 2006. — Т. 361(1470). — P. 969-1006.
- ↑ Schopf JW Fossil evidence of Archaean life // Philos Trans R Soc Lond B Biol Sci. — 2006. — Т. 361(1470). — P. 869-85.
- ↑ Valentine JW, Jablonski D, Erwin DH Fossils, molecules and embryos: new perspectives on the Cambrian explosion // Development. — 1999. — Т. 126. — P. 851-859.
- ↑ Futuyma, 2005, p. 97-99
- ↑ Ohno S The reason for as well as the consequence of the Cambrian explosion in animal evolution // J Mol Evol. — 1999. — Т. 44. — P. S23-27.
- ↑ Valentine JW, Jablonski D Morphological and developmental macroevolution: a paleontological perspective // Int J Dev Biol. — 2003. — Т. 47(7-8). — P. 517-522.
- ↑ Waters ER Molecular adaptation and the origin of land plants // Mol Phylogenet Evol. — 2003. — Т. 29(3). — P. 456-463.
- ↑ Mayhew PJ Why are there so many insect species? Perspectives from fossils and phylogenies // Biol Rev Camb Philos Soc. — 2007. — Т. 82(3). — P. 425-454.
- ↑ Benton, M. J. Vertebrate Paleontology. — Blackwell Science Ltd, 2005. — 472 с. — P. 111-115. — ISBN 978-0-632-05637-8
- ↑ 1 2 Roger Lewin. Human Evolution: An Illustrated Introduction. — John Wiley & Sons. — ISBN 1405103787
- ↑ Michael Steiper, Nathan Young. Primates // The Timetree of Life / S. Blair Hedges,Sudhir Kumar. — Oxford University Press, 2007. — P. 483. — ISBN 0199535035
- ↑ 1 2 Michael Ruse,Joseph Travis. volution: The First Four Billion Years. — Belknap Press of Harvard University Press, 2009. — P. 256-289.
- ↑ Александр Марков. Эволюция человека. — М.: Астрель, Corpus, 2011. — Т. 1. — 216-224 с. — ISBN 978-5-271-36293-4
- ↑ Soares P, Ermini L, Thomson N, Mormina M, Rito T, Rohl A, Salas A, Oppenheimer S, Macaulay V, Richards MB Correcting for purifying selection: an improved human mitochondrial molecular clock // Am J Hum Genet. — 2009. — Т. 84. — № 6. — С. 740-759.
- ↑ J. J. Bull, H. A. Wichman Applied evolution // Annual Review of Ecology and Systematics. — 2001. — Т. 32. — С. 183-217.
- ↑ Doebley JF, Gaut BS, Smith BD The molecular genetics of crop domestication // Cell. — 2006. — Т. 127. — № 7. — С. 1309-1321.
- ↑ Maher B Evolution: Biology's next top model? // Nature. — 2009. — Т. 458. — С. 695-698.
- ↑ Borowsky R Restoring sight in blind cavefish // Curr Biol. — 2008. — Т. 18. — № 1. — С. R23-24.
- ↑ Gross JB, Borowsky R, Tabin CJ A novel role for Mc1r in the parallel evolution of depigmentation in independent populations of the cavefish Astyanax mexicanus // PLoS Genet. — 2009. — Т. 5. — № 1.
- ↑ FRASER AS Monte Carlo analyses of genetic models // Nature. — 1958. — Т. 181. — С. 208-2090.
- ↑ Jamshidi M Tools for intelligent control: fuzzy controllers, neural networks and genetic algorithms // Philos Transact A Math Phys Eng Sci. — 2003. — Т. 361. — С. 1781-1808.
- ↑ Ross, M. R [Who Believes What? Clearing up Confusion over Intelligent Design and Young-Earth Creationism Who Believes What? Clearing up Confusion over Intelligent Design and Young-Earth Creationism] // Journal of Geoscience Education. — 2008. — Т. 53. — № 3. — С. 319.
- ↑ Hameed, Salman Science and Religion: Bracing for Islamic Creationism // Science. — 2009. — Т. 322. — С. 1637–1638.
- ↑ Spergel D. N.; Scott, EC; Okamoto, S Science communication. Public acceptance of evolution // Science. — 2006. — Т. 313. — С. 765-766.
- ↑ См.например:
- National Science Teachers Association Disappointed About Intelligent Design Comments Made by President Bush (англ.). nsta.org (2005). Архивировано из первоисточника 21 июня 2012. Проверено 28 января 2012.
- Royal Society issues statement on evolution (англ.). ncse.com (2006). Архивировано из первоисточника 21 июня 2012. Проверено 28 января 2012.
- Science and Creationism: A View from the National Academy of Sciences. — 1999.
- Mark Perakh, Matt Young. Is Intelligent Design Science? / Ed. Matt Young, Taner Edis. — Why Intellegent Design Fails. — Rutgers University Press, 2004. — P. 185-196. — 238 p. — ISBN 0-8135-3433-X
- Gregory Neil Derry. What Science Is and How It Works. — Princeton University Press, 2002. — С. 170-174. — ISBN 9780691095509
Литература
- Научно-популярная
- Еськов К. Ю. История Земли и жизни на ней: От хаоса до человека. — М.: НЦ ЭНАС, 2004. — ISBN 5-93196-477-0
- Марков А. В. Рождение сложности. — М.: Астрель, 2010. — 527 с. — 4000 экз. — ISBN 978-5-271-24663-0
- Учебная и научная
- Воронцов Н. Н. История эволюционного учения. — М.: Изд.отдел УНЦ ДО МГУ, 1999. — 640 с.
- Грант В. Эволюционный процесс. Критический обзор эволюционной теории. — М.: Мир, 1991. — 488 с.
- Иорданский Н. Н. Эволюция жизни. — М.: Академия, 2001. — 425 с.
- Северцов А. С. Теория эволюции. — М.: Владос, 2005. — 380 с. — 10 000 экз.
- Солбриг О., Солбриг Д. Популяционная биология и эволюция. — М.: Мир, 1982. — 244 с.
- На английском языке
- Futuyma D. J. Evolution. — Sunderland: Sinauer Associates, 2005. — ISBN 0-878-93187-2
- Gould S. J. The Structure of Evolutionary Theory. — Cambridge: Belknap Press of Harvard University Press, 2002. — 1443 экз. — ISBN 978-0-674-00613-3
- Mayr E. What Evolution Is. — New York: Basic Books, 2001. — 336 p.
- Ridley M. Evolution. — 3rd ed. — Wiley-Blackwell, 2004. — 751 p. — ISBN 978-1-4051-0345-9
- Shapiro J. Evolution: A View from the 21st Century. — FT Press Science, 2011. — 253 p. — ISBN 978-0-13-278093-3
Ссылки

Портал «Эволюционная биология» 
Эволюция в Викисловаре? 
Эволюция в Викицитатнике? - «Проблемы эволюции» — сайт российских учёных. Архивировано из первоисточника 19 мая 2012. Проверено 11 марта 2012.
- Новости проекта «Элементы.ру» по теме «Эволюция». Архивировано из первоисточника 19 мая 2012. Проверено 11 марта 2012.
- Новости раздела «Эволюция» проекта «Вся Биология». Архивировано из первоисточника 19 мая 2012. Проверено 11 марта 2012.
- Проект «Антропогенез.ру». Архивировано из первоисточника 19 мая 2012. Проверено 11 марта 2012.
- Сайт «Теория эволюции как она есть». Архивировано из первоисточника 19 мая 2012. Проверено 11 марта 2012.
- Статьи по теме «Эволюция» проекта «New scientist»accessdate=2012-03-11 (англ.). Архивировано из первоисточника 19 мая 2012. Проверено 11 марта 2012.
 Эволюция • Доказательства эволюции
Эволюция • Доказательства эволюцииЭволюционные процессы Адаптация • Преадаптация • Экзаптация • Абаптация • Видообразование • Микроэволюция • Макроэволюция Генетика популяций Дрейф генов • Естественный отбор • Изоляция • Поток генов Происхождение жизни Возникновение жизни • Химическая эволюция • Гипотеза мира РНК Исторические концепции Дарвинизм • Ламаркизм • Пангенезис • Ортогенез • Номогенез • Сальтационизм • Катастрофизм Современные теории Синтетическая теория эволюции • Теория прерывистого равновесия • Нейтральная теория молекулярной эволюции • Эволюционная биология развития • Эпигенетическая теория эволюции Эволюция таксонов Растения • Земноводные • Рептилии • Птицы • Млекопитающие • Китообразные • Человек История эволюционного учения • Хронология эволюции • История жизни на Земле 
Эта статья входит в число избранных статей русскоязычного раздела Википедии. 
Эта статья участвует в конкурсе Статьи года — 2012
Правила конкурса • Жюри конкурса • Страница обсуждения статей-кандидатов • Страница голосования
Категории:- Эволюция
- Эволюционная биология





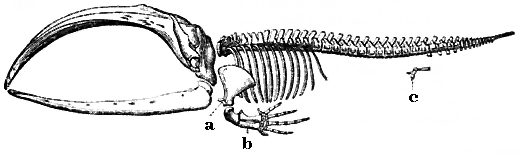





Wikimedia Foundation. 2010.