- Видообразование
-
Видообразова́ние — процесс возникновения новых видов[1] Видообразование — это процесс изменения старых видов и появления новых в результате накопления новых признаков. При этом генетическая несовместимость новообразованных видов, то есть их неспособность производить плодотворное потомство или вообще потомство, при скрещивании называется межвидовым барьером, или барьером межвидовой совместимости.
Существуют разнообразные теории, объясняющие механизмы видообразования, ни одна из которых не считается общепризнанной и полностью доказанной. Одна из причин этого — сложность эмпирической проверки из-за долговременности изучаемого процесса.
Согласно синтетической теории эволюции (СТЭ), основой для видообразования является наследственная изменчивость организмов, ведущий фактор — естественный отбор. В СТЭ выделяют два способа видообразования: географическое, или аллопатрическое, и экологическое, или симпатрическое.
Содержание
Симпатрическое (экологическое) видообразование
Связано с расхождением групп особей одного вида и обитающих на одном ареале по экологическим признакам. При этом особи с промежуточными характеристиками оказываются менее приспособленными. Расходящиеся группы формируют новые виды.
Симпатрическое видообразование может протекать несколькими способами. Один из них — возникновение новых видов при быстром изменении кариотипа путём полиплоидизации. Известны группы близких видов, обычно растений, с кратным числом хромосом. Другой способ симпатрического видообразования — гибридизация с последующим удвоением числа хромосом. Сейчас известно немало видов, гибридогенное происхождение и характер генома которых может считаться экспериментально доказанным. Третий способ симпатрического видообразования — возникновение репродуктивной изоляции особей внутри первоначально единой популяции в результате фрагментации или слияния хромосом и других хромосомных перестроек. Этот способ распространён как у растений, так и у животных. Особенностью симпатрического пути видообразования является то, что он приводит к возникновению новых видов, всегда морфологически близких к исходному виду. Лишь в случае гибридогенного возникновения видов появляется новая видовая форма, отличная от каждой из родительских.
Аллопатрическое (географическое) видообразование
Вызывается разделением ареала вида на несколько изолированных частей.Возникновение географических преград (горных хребтов, морских проливов и пр.) приводит к возникновению изолятов- географически изолированных популяций. При этом на каждую такую часть отбор может действовать по-разному, а эффекты дрейфа генов и мутационного процесса будут явно отличаться. Тогда со временем в изолированных частях будут накапливаться новые генотипы и фенотипы. Особи в разных частях ранее единого ареала могут изменить свою экологическую нишу. При таких исторических процессах степень расхождения групп может достигнуть видового уровня.
«Мгновенное» видообразование на основе полиплоидии
Не предполагает деление ареала на части и формально является симпатрическим. При этом за несколько поколений в результате резких изменений в геноме формируется новый вид.
Сальтационно происходит видообразование на основе полиплоидии у растений.
Гибридогенное видообразование
При скрещивании различных видов потомство обычно бывает стерильным. Это связано с тем, что число хромосом у разных видов различно. Несходные хромосомы не могут нормально сходиться в пары в процессе мейоза, и образующиеся половые клетки не получают нормального набора хромосом. Однако, если у такого гибрида происходит геномная мутация, вызывающая удвоение числа хромосом, то мейоз протекает нормально и дает нормальные половые клетки. При этом гибридная форма приобретает способность к размножению и утрачивает возможность скрещивания с родительскими формами. Кроме того, межвидовые гибриды растений могут размножатся вегетативным путем.
Существующие в природе естественные ряды гибридных видов растений возникли, вероятно, именно таким путем. Так, известны виды пшеницы с 14, 28 и 42 хромосомами, виды роз с 14, 28, 42 и 56 хромосомами и виды фиалок с числом хромосом, кратным 6 в интервале от 12 до 54. По некоторым данным, гибридогенное происхождение имеют не менее трети всех видов цветковых растений[2].
Гибридогенное происхождение доказано и для некоторых видов животных, в частности, скальных ящериц, земноводных и рыб[3]. Некоторые виды кавказских ящериц, имеющих гибридогенное происхождение, триплоидны и размножаются с помощью партеногенеза.
Наблюдение видообразования
Судя по палеонтологической летописи и по измерениям скорости мутаций, полная несовместимость геномов, делающая невозможным скрещивание, достигается в природе в среднем за 3 млн лет[4]. А значит, наблюдение образования нового вида в естественных условиях в принципе возможно, но это редкое событие. В то же время, в лабораторных условиях скорость эволюционных изменений может быть увеличена, поэтому есть основания надеяться увидеть видообразование у лабораторных животных[5][6].
Известны многие случаи видообразования посредством гибридизации и полиплоидизации у таких растений как конопля, крапива, первоцвет, редька, капуста, а также у различных видов папоротников. В ряде случаев видообразование у растений происходило без гибридизации и полиплоидизации (кукуруза[7], стефаномерия (англ.)русск. Stephanomeria malheurensis из семейства астровых[8]).
Дрозофилы, также известные как плодовые мухи, входят в число наиболее изученных организмов. С 1970-х годов зафиксированы многие случаи видообразования у дрозофил. Видообразование происходило, в частности, за счёт пространственного разделения, разделения по экологическим нишам в одном ареале, изменения поведения при спаривании, дизруптивного отбора, а также за счет сочетания эффекта основателя с эффектом бутылочного горлышка (в ходе экспериментов founder-flush).
Видообразование наблюдалось в лабораторных популяциях комнатных мух, мух Eurosta solidaginis, яблонных мух-пестрокрылок, мучных жуков, комаров и других насекомых.
Известны случаи, когда в результате давления отбора (в присутствии хищников) одноклеточные зелёные водоросли из рода хлорелла образовывали многоклеточные колониальные организмы, а у бактерий в аналогичных условиях менялось строение и увеличивались размеры (c 1,5 до 20 микрометров за 8—10 недель). Являются ли эти случаи примерами видообразования, зависит от того, какое используется определение вида (при бесполом размножении нельзя использовать критерий репродуктивной изоляции)[9][10].
Видообразование также наблюдалось и у млекопитающих. Шесть случаев видообразования у домовых мышей на острове Мадейра за последние 500 лет были следствием исключительно географической изоляции, генетического дрейфа и слияния хромосом. Слияние двух хромосом — это наиболее заметное различие геномов человека и шимпанзе, а у некоторых популяций мышей на Мадейре за 500 лет было девять подобных слияний[11][12].
Rhagoletis pomonella
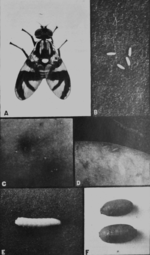
Яблонная муха (Rhagoletis pomonella) является примером ранней стадии симпатрического видообразования[13]. Первоначально вид обитал в восточной части США. До появления европейцев личинки этих мух развивались только в плодах боярышника. Однако с завозом в Америку яблонь (первое упоминание яблонь в Америке — 1647 год), открылась новая экологическая ниша. В 1864 году личинки Rhagoletis pomonella были обнаружены в яблоках, тем самым зафиксирована яблонная раса этого вида[14]. За полтора века наблюдений расы очень сильно разошлись. Они почти не скрещиваются друг с другом (уровень гибридизации не превышает 4—6 %). Яблоневая раса спаривается почти исключительно на яблонях, а боярышниковая — на боярышнике, что, учитывая разное время созревания плодов, приводит к репродуктивной изоляции. В скором времени возможно выделение из этих рас самостоятельных видов.
Кроме того вслед за Rhagoletis pomonella подобное видообразование произошло у наездника Diachasma alloeum, личинки которого паразитируют на личинках мух. После появление новой расы мух появилась и новая раса наездников, предпочитающая паразитировать на яблонных мухах[15].
Опыты Г. Шапошникова по искусственному видообразованию
Литература
- Дарвин Ч., Происхождение видов путём естественного отбора…, Соч., т. 3, М.—Л., 1939
- 3авадский К. М., Вид и видообразование, Л., 1968
- Майр Э., Зоологический вид и эволюция, пер. с англ., М., 1968
- Симпсон Д., Темпы и формы эволюции, пер. с англ., М., 1948
- Тимофеев-Ресовский Н. Видообразование, Воронцов Н. Н., Яблоков А. Видообразование, Краткий очерк теории эволюции, М., 1969
См. также
- Макроэволюция
- Симпатрическое видообразование
- Перипатрическое видообразование
- Аллопатрическое видообразование
Примечания
- ↑ Значение «Видообразование» в Большой советской энциклопедии
- ↑ http://sbio.info/page.php?id=57
- ↑ http://vivovoco.rsl.ru/VV/JOURNAL/NATURE/09_00/PARTENO.HTM
- ↑ Douglas J. Futuyma. Evolutionary Biology. — 3-е изд. — Sunderland, Massachusetts: Sinauer Associates, 1998. — ISBN 0-87893-189-9 (англ.)
- ↑ Gingerich, P. D. Rates of evolution: Effects of time and temporal scaling (англ.) // Science. — 1983. — Т. 222. — № 4620. — P. 159—161. DOI:10.1126/science.222.4620.159
- ↑ Speciations (англ.). Архив TalkOrigins.org. Архивировано из первоисточника 21 августа 2011.
- ↑ Pasterniani, E. Selection for reproductive isolation between two populations of maize, Zea mays L // Evolution. — 1969. — Т. 23. — P. 534—547.
- ↑ Gottlieb, L. D. Genetic differentiation, sympatric speciation, and the origin of a diploid species of Stephanomeria // American Journal of Botany. — 1973. — Т. 60. — P. 545—553.
- ↑ Boraas, M. E. 1983. Predator induced evolution in chemostat culture. EOS. Transactions of the American Geophysical Union. 64:1102. (англ.)
- ↑ Shikano, S., L. S. Luckinbill, Y. Kurihara. Changes of traits in a bacterial population associated with protozoal predation (англ.) // Microbial Ecology. — 1990. — Т. 20. — № 1. — P. 75—84. DOI:10.1007/BF02543868
- ↑ Observed Instances of Speciation (англ.). Архив TalkOrigins.org. Архивировано из первоисточника 21 августа 2011.
- ↑ Britton-Davidian, J., J. Catalan, et al. Rapid chromosomal evolution in island mice (англ.) // Nature. — 2000. — Т. 403. — P. 158. DOI:10.1038/35003116
- ↑ As the Worm Turns: Speciation and the Apple Maggot Fly — Case Study Collection — National Center for Case Study Teaching in Science
- ↑ Элементы.ру: Цепная реакция видообразования.
- ↑ Знание-сила 9/2009
Ссылки
- Проблемы эволюции
- Видообразование
- Начальные этапы видообразования воспроизведены в эксперименте на дрожжах
- Опыты по «искусственному видообразованию»
- Энтомологическое обозрение
- Морфологическая дивергенция и конвегенция в эксперименте с тлями
- Возникновение и утрата репродуктивной изоляции и критерий вида
- Динамика клонов, популяций и видов и эволюция
- Формы видообразования


 Эволюция • Доказательства эволюции
Эволюция • Доказательства эволюцииЭволюционные процессы Адаптация • Преадаптация • Экзаптация • Абаптация • Видообразование • Микроэволюция • Макроэволюция Генетика популяций Дрейф генов • Естественный отбор • Изоляция • Поток генов Происхождение жизни Возникновение жизни • Химическая эволюция • Гипотеза мира РНК Исторические концепции Дарвинизм • Ламаркизм • Пангенезис • Ортогенез • Номогенез • Сальтационизм • Катастрофизм Современные теории Синтетическая теория эволюции • Теория прерывистого равновесия • Нейтральная теория молекулярной эволюции • Эволюционная биология развития • Эпигенетическая теория эволюции Эволюция таксонов Растения • Земноводные • Рептилии • Птицы • Млекопитающие • Китообразные • Человек История эволюционного учения • Хронология эволюции • История жизни на Земле Категория:- Микроэволюция

Wikimedia Foundation. 2010.