- Первичный суп
-
Химическая эволюция или пребиотическая эволюция — первый этап эволюции жизни, в ходе которого органические, пребиотические вещества возникли из неорганических молекул под влиянием внешних энергетических и селекционных факторов и в силу развертывания процессов самоорганизации, свойственных всем относительно сложным системам, которыми бесспорно являются все углерод-содержащие молекулы.
Также этими терминами обозначается теория возникновения и развития тех молекул, которые имеют принципиальное значение для возникновения и развития живого вещества.
Всё, что известно о химизме вещества, позволяет ограничить проблему Х.Э. рамками так наз. "водно-углеродного шовинизма", постулирующего, что жизнь в нашей Вселенной представлена в единственно возможном варианте: в качестве "способа существования белковых тел", осуществимого благодаря уникальному сочетанию полимеризационных свойств углерода и деполяризующих свойств жидко-фазной водной среды, как совместно необходимых и|или достаточных(?) условий для возникновения и развития всех известных нам форм жизни. При этом подразумевается, что, по крайней мере, в пределах одной сформировавшейся биосферы может существовать только один, общий для всех живых существ данной биоты код наследственности, но пока остается открытым вопрос, существуют ли иные биосферы вне Земли и возможны ли иные варианты генетического аппарата.
Также неизвестно, когда и где началась химическая эволюция. Возможны любые сроки по окончанию второго цикла звёздообразования, наступившего после конденсации продуктов взрывов первичных сверхновых звезд, поставляющих в межзвездное пространство тяжелые элементы (с атомной массой более 26). Второе поколение звёзд , уже с планетными системами, обогащенными тяжёлыми элементами, которые необходимы для реализации Х.Э. появилось через 0,5-1,2 млрд. лет после Большого взрыва. При выполнении некоторых вполне вероятных условий, для запуска Х.Э. может быть пригодна практически любая среда: глубины океанов, недра планет, их поверхности, протопланетные образования и даже облака межзвёздного газа, что подтверждается повсеместным обнаружением в космосе методами астрофизики многих видов органических веществ - альдегидов, спиртов, сахаров и даже аминокислоты глицина, которые вместе могут служить исходным материалом для Х.Э., имеющей своим конечным результатом возникновение жизни.
Содержание
Методология исследования химической эволюции
Исследование химической эволюции осложняется тем, что в настоящее время знания о геохимических условиях древней Земли не являются достаточно полными.
Поэтому, кроме геологических, привлекаются также астрономические данные. Так, условия на Венере и Марсе рассматривают как близкие к тем, что были на Земле на различных этапах ее эволюции.
Основные данные о химической эволюции получены в результате модельных экспериментов, в ходе которых удалось получить сложные органические молекулы при различных химических составах атмосферы, гидросферы и литосферы и климатических условиях.
На основе имеющихся данных был выдвинут ряд гипотез о конкретных механизмах и непосредственных движущих силах химической эволюции.
Абиогенез
Абиогене́з — образование органических соединений, распространённых в живой природе, вне организма без участия ферментов.
В широком смысле абиогенез — возникновение живого из неживого, то есть исходная гипотеза современной теории происхождения жизни. В 20-х годах XX века академик Опарин предположил, что в растворах высокомолекулярных соединений могут самопроизвольно образовываться зоны повышенной концентрации, которые относительно отделены от внешней среды и могут поддерживать обмен с ней. Он назвал их Коацерватные капли, или просто коацерваты. В 1953 году Стэнли Миллером экспериментально осуществлён абиогенный синтез аминокислот и других органических веществ в условиях, воспроизводящих условия первобытной Земли.
Существует также теория гиперциклов; согласно которой первые проявления жизни были соответственно в виде гиперциклов — комплекса сложных каталитических реакций, продукты выхода которых являются катализаторами для последующих реакций.
В 2008 году американские биологи сделали важный шаг к пониманию начальных этапов зарождения жизни. Им удалось создать «протоклетку» с оболочкой из простых липидов и жирных кислот, способную втягивать из окружающей среды активированные нуклеотиды — «кирпичики», необходимые для синтеза ДНК [1].
Обзор темы
Гипотезы химической эволюции должны объяснять различные аспекты:
- Небиологическое начало биомолекул, то есть их развитие из неживущих и, соответственно, неорганических предшественников.
- Появление способных к само-репликации и само-изменению химических информационных систем, то есть возникновение клетки.
- Появление взаимной зависимости функции (ферментов) и информации (РНК, ДНК).
- Условия среды Земли в период от 4,5 до 3,5 млрд лет назад.
Большой вклад в прояснение этих вопросов, среди прочих, сделали следующие ученые:
- Александр Опарин: Коацерваты.
- Гарольд Юри и Стэнли Миллер в 1953: Возникновение простых биомолекул в симулируемой древней атмосфере.
- Сидней Фокс: Микросферы из протеноидов.
- Томас Чек (университет Колорадо) и Сидней Алтман (университет Yale New Haven Connecticut) в 1981: Автокаталитическое РНК-деление: «Рибозимы» объединяют катализ и информацию в молекуле. Они в состоянии вырезать себя из более длинной цепи РНК и соединять остающиеся концы снова.
- Уолтер Гилберт (Гарвард, университет Кембридж) разрабатывает в 1986 идею мира РНК.
- Гюнтер фон Кидровски (Рур-университет Бохум) представляет в 1986 году первую само-реплицирующуюся систему на основе ДНК, важный вклад в понимание функций роста само-реплицирующихся систем
- Манфред Эйген (институт Макса Планка факультет биофизической химии, Геттинген): Эволюция ансамблей молекул РНК. Гиперцикл.
- Юлий Ребек (Кембридж) создаёт искусственную молекулу (Aminoadenosintriazidester), которая само-реплицируется в растворе хлороформа. Копии все же идентичны образцу, так что эволюция для этих молекул невозможна.
- Джон Корлис (Goddard центр космических полётов — НАСА): Термальные источники морей поставляют энергию и химикалии, которые делают возможными абсолютно независимую от столкновений с метеоритами химическую эволюцию. Ещё сегодня они являются средой жизни для первоначальных по многих признаках археобактерий (Archaea).
- Гюнтер Вэхтерсхойзер (Мюнхен): Первые само-реплицирующиеся структуры с обменом веществ возникли на поверхности пирита. Пирит (сульфид железа) поставил для этого необходимую энергию. На растущих и снова распадающихся кристаллах пирита эти системы могли расти и размножаться, и различные популяции конфронтировали с разным условиям среды (условия отбора).
- А. Г. Cairns-Smith (университет Глазго) и Дэвид К. Мауерцалл (Rockefeller-Universität New York, Нью-Йорк) видят в глиняных минералах систему, которая сначала сама подчинена химической эволюции, из-за чего возникает много различных, самореплицирующихся кристаллов. Эти кристаллы притягивают своим электрическим зарядом органические молекулы и катализируют синтез комплексных биомолекул, при чём объём информации кристаллических структур служит сначала матрицей. Эти органические соединения становятся все более сложными, до тех пор пока они не могут размножаться без помощи глиняных минералов.
- Вольфганг Вайганд, Марк Дерр и др. (Институт Макса Планка факультет биогеохимии, Йена) показали в 2003, что сульфид железа может катализировать синтез аммиака из молекулярного азота.
Унифицированная модель химической эволюции ещё не разработана, возможно потому, что основные принципы ещё не открыты.
Предварительные рассуждения
Биомолекулы
Пребиотический синтез сложных соединений молекул может делится на три последовательных этапа:
- Возникновение простых органических соединений (спиртов, кислот, гетероциклических соединений: пуринов, пиримидинов и пирролов) из неорганических материалов.
- Синтез более сложных органических соединений-"биомолекул" — представителей наиболее распостранённых классов метаболитов, в том числе и мономеров — структурных единиц биополимеров (моносахаридов, аминокислот, жирных кислот, нуклеотиды) из простых органических соединений.
- Возникновение сложных биополимеров (полисахариды, белки, нуклеиновые кислоты) из основных структурных единиц — мономеров.
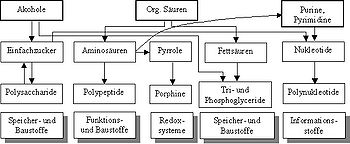 Биомолекулы — возникновение и функция
Биомолекулы — возникновение и функцияОдним из вопросов является химический состав среды в которой осуществлялся пребиологический синтез, в том числе какие неорганические компоненты являлись источниками различных элементов, входящих в состав различных органических соединений.
Элементный состав биомолекул: C H O N S P Углеводы X X X Жиры X X X X Фосфолипиды X X X X X Белки X X X X X Нуклеотиды X X X X X Порфирины X X X X Возможные неорганические источники элементов: Восстановленная форма Окислённая форма Углерод (C) Метан (CH4), Монооксид углерода (угарный газ)]] (CO) Углекислый газ (CO2) Водород (H) Водород (H2) Вода (H2O) Кислород (O) H2O Кислород (O2) Азот (N) Аммиак (NH3) Нитраты (NO3-) Сера (S) Сероводород (H2S) Сульфаты (SO42-) Фосфор (P) Фосфаты (PO43-) Все гипотезы исходят из того, что помимо воды и фосфатов в начале в достаточной массе находились в достаточном количестве только упрощённые формы привычных сегодня химических соединений, так как древняя атмосфера не содержала молекулярный кислород.
В качестве источника энергии, инициирующей синтез, в это время могли выступать Ультрафиолетовое излучение Солнца, тепло вулканических процессов, ионизирующие излучения радиоактивного распада и электрические разряды. Существуют также теории, в рамках которых источником необходимой для возникновения биомолекул энергии могут служить окислительно-восстановительные процессы между вулканическими газами (восстановитель) и частично окисляющими сульфидными минералами, например пиритом (FeS2)
Развитие древней атмосферы
Развитие земной атмосферы является частью химической эволюции и к тому же важным элементом истории климата. Сегодня её разделяют на четыре важные ступени развития.
В начале происходило образование химических элементов в космосе и появление из них земли — приблизительно 4,56 миллиардов лет назад. Предположительно наша планета уже довольно рано имела атмосферу из водорода (H2) и гелия (He), которая была однако снова потеряна в космическое пространство. Астрономы исходят так же из того, что из-за относительно высоких температур и эффектов солнечного ветра на земле и других близлежащих солнцу планетах могло остаться только мало легких химических элементов (включая углерод, азот и кислород). Все эти элементы, составляющие сегодня основную часть биосферы, были занесены, по этой теории, ударами комет из более внешних участков солнечной системы лишь через большой промежуток времени, когда протопланеты немного остыли. В течение первых нескольких миллионов лет после возникновения солнечной системы постоянно повторялись столкновения с небесными телами, вызванные ими коллизии уничтожали глобальными стерилизациями образованные в это время живые системы. Поэтому появление жизни смогло начаться только после накопления воды на длительное время хотя бы в самых глубоких местах моря.
 Следы вулканической активности: отложения серы на краях Halema’uma’u-кратера вулкана Мауна Лоа на Гаваях
Следы вулканической активности: отложения серы на краях Halema’uma’u-кратера вулкана Мауна Лоа на Гаваях Извержение вулкана — самая захватывающая форма вулканической деятельности
Извержение вулкана — самая захватывающая форма вулканической деятельностиС медленным остыванием земли, вулканической деятельностью (выделение газов из недр земли) и глобальным распределением материалов упавших комет возникла вторая атмосфера земли. Скорее всего она состояла из водяного пара (H2O до 80 %), углекислого газа (CO2; до 20 %), сероводорода (до 7 %), аммиака и метана. Высокий процент водяного пара объясняется тем, что поверхность земли была на тот момент ещё слишком горяча для образования морей. Прежде всего из воды, метана и аммиака в условиях молодой земли могли образоваться небольшие органические молекулы (кислоты, спирты, аминокислоты), позднее так же органические полимеры (полисахариды, жиры, полипептиды), которые были нестабильны в кислотной атмосфере.
После охлаждения атмосферы ниже температуры кипения воды наступил очень длительный дождь, который и образовал океаны. Насыщенность других газов атмосферы относительно водяного пара повысилась. Высокое ультрафиолетовое облучение обусловило фотохимический распад воды метана и аммиака, в результате чего накопились углекислый газ и азот. Лёгкие газы — водород и гелий — уносились в космос, углекислый газ растворялся в больших количествах в океане, окисляя воду. Значение pH упало до 4. Инертный и малорастворимый азот N2 накапливался со временем и образовывал около 3,4 миллиардов лет назад основную составляющую атмосферы.
Выпадение в осадок, прореагировавшего с ионами металлов, растворенного углекислого газа (карбонаты) и дальнейшее развитие живых существ, которые ассимилировали углекислый газ, привело к уменьшению CO2-концентрации и поднятию значения pH в водоёмах.
Кислород O2 играет важнейшую роль в дальнейшем развитии атмосферы. Он образовался с появлением способных к фотосинтезу живых существ, предположительно цианобактерий (сине-зелёных водорослей) или им подобных прокариотов. Ассимиляция ими углекислого газа привела к дальнейшему понижению кислотности, насыщенность атмосферы кислородом оставалась все-таки довольно низкой. Причина этого — незамедлительное использование растворенного в океане кислорода для окисления двухвалентных ионов железа и других окисляемых соединений. Около двух миллиардов лет назад этот процесс завершился, и кислород стал постепенно накапливаться в атмосфере.
Очень реактивный кислород легко окисляет восприимчивые органические биомолекулы и становится таким образом фактором отбора окружающей среды для ранних организмов. Только немногие анаэробные организмы смогли переместиться в свободные от кислорода жизненные пространства, другая часть выработала ферменты (например, каталазы), которые делают кислород не опасным. В некоторых микроорганизмах из подобных энзимов развились комплексные мембранные энзимы — конечные оксидазы, которые метаболически использовали присутствующий кислород для накопления энергии необходимой для роста собственной клетки — конечная стадия окисления в аэробной цепи дыхания. В зависимости от организма имеются различные формы конечных оксидаз, например хинол-оксидаза или цитохром C — оксидаза, которые различаются активными центрами, содержащими ионы меди и гемины. Это дает основание полагать, что они произошли различными параллельными путями развития. Во многих случаях в одном организме встречаются различные типы конечных оксидаз. Эти энзимы являются последними в цепи последовательно задействованных комплексов энзимов, которые энергию окислительно-восстановительных процессов сохраняют переносом протонов или ионов натрия в форме трансмембранного электрического потенциала. Последний другим комплексом энзимов преобразовывается снова в химическую энергию в форме АТФ. Синтез АТФ и прочих компонент цепи дыхания в эволюционном свете значительно старше конечных оксидаз, так как они играли важную роль уже во многих аэробных процессах обмена веществ (аэробное дыхание, многие процессы брожения, метаногенез) а так же при аноксигенном и оксигенном фотосинтезе.
Миллиард лет назад содержание кислорода в атмосфере перешагнуло планку одного процента и спустя несколько миллионов лет был образован озоновый слой. Сегодняшнее содержание кислорода в 21 % было достигнуто лишь 350 миллионов лет назад и сохраняется с тех пор стабильным.
Значение воды для возникновения жизни
 H2O — химическое соединение, присутствующие при обычных условиях во всех трёх агрегатных состояниях.
H2O — химическое соединение, присутствующие при обычных условиях во всех трёх агрегатных состояниях.Для жизни, как мы её знаем (или определяем), нужна вода как универсальный растворитель. Вода имеет комллекс качеств, которые делают жизнь возможной. Нет данных, что жизнь может возникать и существовать независимо от воды и принято считать, что только наличие воды в жидкой фазе (в определённой области или на определённой планете), делает вероятным возникновение там жизни.
Для синтеза сложных молекул и соблюдения граничных условий автокаталитических реакций между ними (что обеспечивает принципиальную возможность перехода от химической эволюции к эволюции живого вещества), важны следующие характеристики воды:
- Вода находится в жидком состоянии при температуре, в которой стабильны органические молекулы и их синтез возможен только в водных растворах.
- Вода необходима как деполяризующий растворитель для химических реакций, так как она делает возможным гомогенное перемешивание, а имея высокую теплоёмкость, принимает выделяющуюся при реакциях теплоту и предоставляет в распоряжение протоны для катализов.
- Высокая удельная теплоемкость воды и особенности характеристики спектрального поглощения водяных паров (при их зна́чимых долях в объёме планетных оболочек) обеспечивают стабилизацию глобальных колебаний температуры и осмотических процессов (локально могут возникать большие различия), что создает предпосылки к планетарно-уравновешенному климату, неблагоприятные изменения которого, вызванные как энтропийными процессами, так и внешними неблагоприятными факторами, могут быть в определенных пределах скомпенсированы жизнедеятельностью самой биоты, изменяющей интенсивность испарения воды и скорость захоронения углерода (см.гипотеза Геи[2], а также биотическая регуляция[3]).
- Вода и водяной пар поглощают вредное для макромолекул ультрафиолетовое облучение. Однако оно проникает через замёрзшую воду (лед) до определённой глубины.
- Вода, в которой растворены вещества, например, морская вода, образует при вымораживании области разных концентраций веществ, которые окружены мембранами льда (см. гипотеза морского льда). Ограничение от внешней среды и повышенная концентрация веществ считаются необходимыми для образования биологически активных молекул.
- Особенность термодинамики воды (при нагревании от 0 до 3,98°С вода сжимается) предотвращает замерзание водоёмов до дна — холодная вода, как менее плотная, остаётся на поверхности и замерзает, а подо льдом сохраняется положительная температура.
Эксперименты
Эксперимент Миллера — Юри
Одна из самых известных гипотез эволюции была опубликована в двадцатые годы XX столетия русским исследователем А. И. Опариным и британским исследователем Дж. Холдэном. Теория утверждала, что условия на земле того времени благоприятствовали химическим реакциям. Из неорганических соединений в атмосфере и море должны были синтезироваться сложные органические соединения. Необходимая энергия поставлялась очень интенсивным ультрафиолетовым облучением, которое могло беспрепятственно проникать в атмосферу в связи с малым содержанием в ней O2 и O3.
В 1953 году эта теория была обоснована химиками Стэнли Миллером и Гарольдом К. Юри очень хорошими результатами эксперимента с первичным бульоном. Опытным путём ими было доказано, что в среде, похожей на среду с предположительными пребиотическими условиями, посредством притока энергии извне (молнии), из неорганических соединений (вода, метан, аммиак и водород) могут возникнуть аминокислоты и более простые карбоновые и жирные кислоты — одни из важнейших строительных элементов биомолекул (причём современные исследования сохранившегося содержимого колб Миллера показало, что там содержалось большее количество аминокислот, чем смог выявить Миллер[4]).
В более поздних, в большинстве случаев, более сложно построенных опытах с первичным бульоном экспериментаторы смогли получить как все важнейшие строительные элементы живых существ — аминокислоты, жиры, сахара, нуклеотиды, — так и более сложные органические соединения — порфины и изопреноиды.
Хотя этим была показана возможность естественного образования органических молекул, эти результаты сегодня иногда подвергаются критическим оценкам. В эксперименте с первичным бульоном исходили из того, что атмосфера на тот период времени имела щелочной характер, что соответствовало научным представлениям того времени. Сегодня же исходят из слабощелочного или даже нейтрального характера атмосферы, хотя вопрос ещё не окончательно решён и обсуждаются также локальные химические отклонения атмосферных условий, например в окрестностях вулканов. Позднейшими экспериментами была доказана возможность появления органических молекул и в этих условиях, даже таких, которые не получились при первых опытах, но в значительно меньших количествах. Этим часто аргументируется, что происхождение органических молекул другим путём, играло как минимум дополнительную роль. Приводятся также теории происхождения органики в окрестностях Гидротермальных источников срединно-океанических хребтов.
В качестве аргумента против происхождения органических молекул из первичного бульона иногда приводят тот факт, что во время опыта получается рацемат, то есть равная смесь из L и D-форм аминокислот. Соответственно, должен был существовать естественный процесс, в котором отдавалось предпочтение определённому вращению хиральных молекул. Некоторые космобиологи утверждают, что легче доказать происхождение органических соединений в космосе, так как, по их мнению, фотохимические процессы с циркулярно-поляризированным излучением, например от пульсаров, в состоянии уничтожить молекулы только определённого вращения. И действительно у найденных в метеоритах хиральных органических молекул преобладали на 9 % левовращающие. Однако в 2001 году Alan Saghatelian et al.[5] показал, что самореплицирующиеся пептидные системы тоже в состоянии эффективно усиливать молекулы определённого вращения в рацематной смеси, что делает возможным и земное происхождение оптически активных биомолекул.
Дальнейшие реакции
Из появляющихся при эксперименте Миллера-Юри промежуточных продуктов альдегидов и синильной кислоты HCN можно получить при симулируемых условиях земли 4,5 млрд лет назад дальнейшие биомолекулы. Таким образом Хуан Оро удалось в 1961 году осуществить синтез аденина:
Эдукты Продукты 2 Формальдегид HCN H2O Серин 5 Формальдегид Рибоза 5 HCN Аденин Из рибозы, аденина и трифосфата возникает аденозинтрифосфат (АТФ), который используется в организмах в качестве универсального энергоносителя и строительного элемента (как монофосфат) рибонуклеиновых кислот (РНК).
Участие минералов и горных пород
- Кристаллические поверхности могут служить матрицей для растущих макромолекул. При этом различные кристаллические поверхности могут связывать определенные энантиомеры молекул. L-и D аминокислоты присоединяются к разным местам кристалла кальцита.
- Аарон Качальсский (Вайцманн-институт, Израиль) показал, что в водном растворе, содержащем монтмориллонит (один из минералов глин), возможен синтез белков с длиной цепи в больше чем 50 аминокислот с почти 100%-ным выходом.
- Ионы металлов могут выступать в роли катализаторов, доноров электронов или же включаться в биомолекулы.
- Минералы глин в водных растворах часто несут поверхностный электрический заряд и могут таким образом притягивать и удерживать противоположно заряженные органические молекулы.
- В микрополостях горных пород молекулы органических соединений защищены от ультрафиолетового облучения.
Теория Вехтерхойзера
Особенно интенсивная форма содействия минералов и горных пород пребиотическому синтезу органических молекул должна протекать на поверхности минералов сульфида железа. Теория Миллера-Юри имеет существенные ограничения, особенно учитывая ошибочное объяснение полимеризации мономерных составляющих биомолекулы.
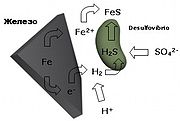 Анаэробные бактерии, обмен веществ которых происходит с участием железа и серы, существуют и сегодня.
Анаэробные бактерии, обмен веществ которых происходит с участием железа и серы, существуют и сегодня. Слиток сросшихся кристаллов сульфида железа FeS2
Слиток сросшихся кристаллов сульфида железа FeS2Альтернативный сценарий был с начала 1980-х годов разработан Гюнтером Вехтерхойзером. По этой теории жизнь на земле возникла на поверхности железно-серных минералов, то есть сульфидов, которые и сегодня образуются посредством геологических процессов, а на молодой земле должны были встречаться гораздо чаще. Большое преимущество этого концепта перед предшественниками в том, что впервые образование комплексных биомолекул связано с постоянным надежным источником энергии. Энергия поступает из восстановления частично окисленных железно-серных минералов, например пирита (FeS2), водородом (уравнение реакции: FeS2 + H2
 FeS + H2S) и поставляет достаточно энергии для эндотермического синтеза мономерных структурных элементов биомолекул и их полимеризации:
FeS + H2S) и поставляет достаточно энергии для эндотермического синтеза мономерных структурных элементов биомолекул и их полимеризации:Fe2+ + FeS2 + H2
2 FeS + 2 H+ ΔG°' = — 44.2 kJ/molДругие тяжелые металлы так же как и ионы железа тоже образуют нерастворимые сульфиды. В дополнение к этому пирит и другие железно-серные минералы имеют положительно заряженную поверхность, на которой могут располагаться, концентрироваться и реагировать между собой преимущественно отрицательно заряженные биомолекулы (органические кислоты, фосфорные эфиры, тиолы). Необходимые для этого вещества (сероводород, моноксид углерода и соли двухвалентного железа) попадают из раствора на поверхность этого «железо-серного мира». Вехтерхойзер привлекает для своей теории и сегодня существующие основопологающие механизмы обмена веществ и выводит из них замкнутый в себе сценарий синтеза комплексных органических молекул (органические кислоты, аминокислоты, сахар, азотистые основания, жиры) из простых неорганических соединений, находящихся в вулканических газах (NH3, H2, CO, CO2, CH4, H2S).
В противоположность к эксперименту Миллер-Юри не привлекаются источники энергии из вне, в форме молнии или ультрафиолетового излучения, кроме того первые ступени синтеза при высоких температурах протекают гораздо быстрее (например катализированные энзимами химические реакции). При температурах подводных вулканов до 350 °C возникновение жизни является вполне представимым. Только позднее при возникновении чувствительных к высоким температурах катализаторам (витамины, белки) эволюция должна была происходить при более низких температурах.
Сценарий Вехтерхойзера хорошо подходит к условиям глубоководных гидротермальных источников, так как имеющийся там переход температур из глубины к поверхности, позволяет подобное распределение реакций. Древнейшие из сегодня живущих микроорганизмов самые жароустойчивые, предельный известный температурный максимум для их роста составляет +113 °C. Кроме того железо-серные активные центры и сегодня задействованы в биохимических процессах, что может указывать на первичное участие Fe-S-минералов в развитии жизни.
Образование макромолекул
Биомакромолекулы — это белки и нуклеиновые кислоты. Увеличение молекулярных цепей (полимеризация) нуждается в энергии и происходит с выделением воды (конденсация). При расщеплении макромолекул (гидролиз) энергия выделяется. Так как химическое равновесие настолько сильно смещено в сторону мономеров, что реакция протекает термодинамически необратимо в сторону гидролиза полимеров, синтез полимеров невозможен без постоянного поступления энергии. Даже с помощью теоретической поддержки испарением воды, добавлением солей (связывают воду) или распадом продуктов, равновесие смещается лишь незначительно. В итоге возникновение жизни очень вероятно связано с надежным источником энергии, который использовался бы для полимеризации.
[Мономеры]n + (n-1) H2O
 n Мономеры + Тепло,
n Мономеры + Тепло,но
Энергия + n Мономеры
[Мономеры]n + (n-1) H2O.В качестве источника энергии в биохимии используется чаще всего АТФ, для образования которого необходимы уже существующие энзимы. В условиях молодой земли возможно снабжение энергией синтеза полимеров гидролитическим расщеплением полифосфатов, что используется некоторыми энзимами вместо ращепления АТФ и сегодня. Но маловероятно, что полифосфаты были в необходимом количестве, так как они могли спонтанно образовываться, при испарении фосфатосодержащих растворах, но также относительно быстро гидролизироваться, растворяясь в воде. Подобные процессы могли происходить на побережье при регулярном приливе и отливе. Но в этом случае все зависимые от воды процессы постоянно прерывались бы, что слишком замедлило бы синтез комплексных соединений. Поэтому обратимся к совсем другой системе, в которой происходит как синтез мономерных состовляющих так и зависимое от постоянного источника энергии образование полимеров, — к анаэробным окислительно-восстановительным реакциям с сульфидами металлов.
Равновесие синтеза полимеров смещается в сторону образования полимеров повышением концетрации мономеров и обезвоживанием продуктов реакции. Условием для этого является ограничение среды реакции, которая имеет лишь ограниченный обмен веществ с внешней средой. Традиционно считалось, что подобные процессы протекают в маленьких прудах с высоким испарением, что основной идеей опирается ещё на труды Ч. Дарвина. Сегодня рассматриваются вулканические регионы океанов с осевшими от гидротермальных источников сульфидами металлов как вполне подходящее место для развития подобного сценария.
Другие варианты решения проблемы имеют сильные ограничения и сложносопоставимы с условиями ранней земли. Преимущественно требуется для одного или нескольких этапов исключение воды, что очень легко осуществить в лаборатории, но не в рассматриваемое время на земле. Одна из подобных систем — это полимеризация карбомидов (R-N=C=N-R) или дициана (N≡C-C≡N) в безводной среде. При этом конденсация исходных составляющих протекает параллельно реакции карбомида, при которой выделяется необходимая энергия:
Энергия + [H-X-OH] + [H-X-OH]
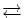 [X-X] + H2O (H-X-OH = мономер, например аминокислота или рибоза)
[X-X] + H2O (H-X-OH = мономер, например аминокислота или рибоза)[R-N ≡ C ≡ N-R] + H2O
[R-NH-CO-NH-R] + Энергия (если R = H возникает мочевина)При ультрафиолетовом излучении из синильной кислоты и образуется дициан, однако, в высыхающей трясине летучая молекула быстро испарилась бы. Если сухая смесь аминокислот нагревается до 130 °C несколько часов, то образуются похожие на белок макромолекулы. При наличии полифосфатов достаточно 60 °C. Эти условия могут образоваться, если вода с растворенными аминокислотами вступает в контакт с горячим вулканическим пеплом.
Если нагреть смесь нуклеотида в присутствии полифосфатов до 55 °C, то хотя и возникают полинуклеотиды, но все же соединение происходит вероятнее за счет 5’- и 2’-C-атомов рибозы, так как оно протекает легче, чем во всех организмах имеющиеся 5’-3’-связи. Из обоих типов полинуклеотидов образуются двойные цепи (сравни со стороением ДНК). Разумеется 5’-3’-двойные цепи стабильнее, чем 5’-2’.
Если на 2’ атоме углерода рибозы отсутствует гидроксильная группа, получается дезоксорибоза. Теперь могут образовываться типичные для ДНК 5’-3’ связи.
Образование пребиотических структур (предшественников клеток)
Клетки поддерживают свои функции тем, что они образуют ограниченные среды для реакций, чтобы разделять друг от друга процессы обмена веществ и исключать нежелательные реакции. Одновременно могут создаваться различия концентраций.
Коацерваты
Известно, что с повышением концентрации многие органические соединения, молекулы которых содержат как гидрофильные, так и гидрофобные участки, способны в водных растворах к мицеллообразованию, то есть выделению микрокапелек органической фазы. Мицеллообразование наблюдается также при высаливании, то есть при увеличении концентрации солей в коллоидных растворах биополимеров-полиэлектролитов, при этом выделяются микрокапли диаметром 1—500 мкм, содержащие биополимеры в высокой концентрации.
Александр Иванович Опарин (1894—1980) главным образом изучал возможности обмена веществ в коацерватах (от латинского coacervatio — сгусток или куча) — модельных мицеллах. Им было показано, что ограниченные среды с простым обменом веществ могут возникнуть исключительно самоорганизацией, при условии присутствия катализаторов со специфическими свойствами. Так как использованные субстанции входят в состав живущих сегодня организмов, Опаринские коацерваты нужно видеть не как предшественники клеток, а как модель-аналог для возникновения предшественников клеток.
Опарин исследовал смеси из белков (гистон и альбумин), белков и углеводов (гистон и гуммиарабик) и белков и полинуклеотидов (гистон или клупеин и ДНК или РНК).
Если капельки из гистона и гуммиарабика содержат фермент фосфорилазу, то они могут поглотить из окружающей среды глюкозу-1-фосфат, создать из неё крахмал и сохранить. Освобожденный фосфат диффундирует наружу. Капелька увеличивается с накоплением крахмала, пока не распадается на мелкие капельки, которые тоже могут содержать фосфорилазу, разумеется, меньше чем исходная капелька. Вместе с тем замедляется обмен веществ. Таким образом становится очевидным, что для сохраниния качеств клетки необходима регенерация ферментов после её деления.
Если к взвеси добавить разлагающий крахмал, фермент амилазу, получаются коацерваты с двухступенчатым обменом веществ: 1) поглощение глюкозы-1-фосфата → синтез крахмала; 2) вывод фосфата → расщепление крахмала и вывод мальтозы.
Микросферы
В 1970 году Сидней Фокс смог подтвердить, что из подобных белку продуктов, которые возникают при нагревании сухих смесей аминокислоты, а также самосоединением могут возникнуть растущие капельки, так называемые микросферы. Они отграничиваются от окружающей среды мембраной обратного осмоса и поглощают другой протеиноподобный материал из среды. Вследствие чего они растут и разделяются снова на более мелкие капельки. На основании этого Фокс пришёл к выводу, что эти системы имеют ферментные качества, разлагают глюкозу или ведут себя как эстераза или пероксидаза, без добавления ферментов извне.
Мир РНК
Впервые гипотеза мира-РНК была выдвинута в 1986 году Уолтером Гильбертом и гласила, что молекулы РНК были предшественниками организмов.
Гипотеза отталкивается от способности РНК к хранению, передаче, и размножению генетической информации, а также от её способности катализировать реакции в качестве рибозимов. В эволюционной среде молекулы РНК, которые множат преимущественно себя, встречались бы чаще других.
Исходным пунктом являются простые самовоспроизводящиеся молекулы РНК. Некоторые из них имеют способность катализировать синтез белков, которые, в свою очередь, сами катализируют синтез РНК и свой собственный синтез (развитие трансляции). Некоторые молекулы РНК соединяются в двойную РНК-спираль, они развиваются в молекулы ДНК и носители наследственной информации (развитие транскрипции).
Основой служат определенные РНК-молекулы, которые могут копировать любые РНК-образцы включая себя самих. Дженнифер А. Doudna и Джек В. Szostak использовали как образец для развития этого типа РНК производящий разрезание и сплайсинг самой себя интрон прокариотного одноклеточного организма Tetrahymena thermophila. Это подтверждает то, что в рибосомах собственно rРНК являются каталитическими молекулами и таким образом РНК катализирует синтез белка. Однако ограничения состоят в том, чтобы при само-реплицирующихся РНК не моно-, а олигонуклеотиды являются составляющими звеньями и необходимы вспомогательные вещества.
В 2001 году было открыто, что важными каталитическими центрами рибосом являются РНК, а не, как раньше было принято, белки. Это показывает, что каталитическая функция РНК, как она предполагалась в гипотезе мира-РНК, используется сегодня живыми существами.
Так как рибосомы считаются очень первоначальными клеточными органоидами, это открытие считается важным взносом в обоснование гипотезы мира-РНК. Уже можно уверенно сказать, что молекулы РНК — по меньшей мере принципиально — являются в состоянии из аминокислот синтезировать белки.
В этой связи «ПНК» (Пептид — нуклеиновые кислоты) также представляют интерес как возможные предшественники РНК.
Альтернативные возможности появления жизни на земле
Биомолекулы из космоса
Земля все время своего существования подвергается бомбардировке кометами и метеоритами. Особенно интенсивными они были сразу после образования планеты. В некоторых метеоритах были обнаружены простые органические соединения, среди прочих аминокислоты. Одним из возможных доказательств происхождения аминокислот в космосе могла бы быть одинаковая хиральность органических соединений (L-аминокислот и D-сахаров) — у метеоритных аминокислот доказано преобладание L-типа до 9 %. [6]. Но это распределение может быть объяснено и действием неорганических жировых катализаторов. С помощью опытов, при которых симулировались условия космоса, было доказано, что основные органические соединения, в особенности аминокислоты, могут образовываться и в таких условиях (Бернштейн, М. П., et al. 2002. «Racemic amino acids from the ultraviolet photolysis of interstellar ice analogues» Nature 416, 401 и Munoz Caro, G., et al. 2002. «Amino acids from ultraviolet irradiation of interstellar ice analogues» Nature 416, 403).
«Черные курильщики»
Условия среды в этих гидротермальных источниках, возможно, способствовали хемоавтотрофному возниковению жизни. Эта теория отстаивается исследователями Вильямом Мартином и Михаелем Русселем (M. Martin, M. J. Russell, (2003). On the origins of cells : a hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleared cells. Philos. Trans. R. Soc. London Ser. B 358, 59-85).
Ссылки
- ↑ Искусственные протоклетки синтезируют ДНК без помощи ферментов
- ↑ http://elementy.ru/trefil/21138
- ↑ http://www.bioticregulation.ru/ques/ques1_r.php
- ↑ «Получены новые результаты старого эксперимента Стэнли Миллера»
- ↑ A. Saghatelian et al. , 2001, A chiroselective peptide replicator, Nature, 409, 797—801
- ↑ J.R. Cronin, S. Pizzarello, (2002) Enantiomeric Excesses in Meteoritic Amino Acids Science, 275, pp. 951—955 [1]
- Статья об искусственной химической эволюции
- [2]
- [3]
- Заметка «Искусственные протоклетки синтезируют ДНК без помощи ферментов»
См. также
Эволюционная биология Механизмы и процессы Адаптация • Видообразование • Генетический дрейф • Естественный отбор • Наследственность • Изменчивость • Микроэволюция • Макроэволюция • Мутации • Перенос генов Возникновение жизни Химическая эволюция • Гипотеза мира РНК • Биологическая эволюция История эволюционного учения Жоффруизм • Броккизм • Дарвинизм • Ламаркизм • Пангенезис • Ортогенез • Автогенез • Номогенез • Сальтационизм • Катастрофизм • Неокатастрофизм Современные концепциии Синтетическая теория эволюции • Теория прерывистого равновесия • Нейтральная теория молекулярной эволюции • Эпигенетическая теория эволюции • Экосистемная теория эволюции Человек Антропогенез
Wikimedia Foundation. 2010.