- Saccharomycetes
-
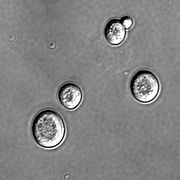 Клетки дрожжей Saccharomyces cerevisiae под микроскопом.
Клетки дрожжей Saccharomyces cerevisiae под микроскопом.Дрожжи — внетаксономическая группа одноклеточных грибов, утративших мицелиальное строение в связи с переходом к обитанию в жидких и полужидких, богатых органическими веществами субстратах. Объединяет около 1500 видов, относящихся к аскомицетам и базидиомицетам.
Общие сведения
Границы группы очерчены нечётко: многие грибы, способные вегетативно размножаться в одноклеточной форме и идентифицируемые поэтому как дрожжи, на других стадиях жизненного цикла образуют развитый мицелий, а в ряде случаев и макроскопические плодовые тела. Раньше такие грибы выделяли в особую группу дрожжеподобных, но сейчас их все обычно рассматривают вместе с дрожжами. Исследования 18S рРНК показали близкое родство с типичными дрожжами видов, способных к росту только в виде мицелия.
Размеры дрожжевых клеток обычно составляют 3—7 мкм в диаметре. Есть данные, что некоторые виды способны вырастать до 40 мкм[1].
Дрожжи имеют большое практическое значение, особенно пекарские или пивные дрожжи (Saccharomyces cerevisiae). Некоторые виды являются факультативными и условными патогенами. К настоящему времени полностью расшифрован геном дрожжей Saccharomyces cerevisiae (они стали первыми эукариотами, чей геном был полностью секвенирован) и Schizosaccharomyces pombe.[2]
История
Русское слово «дрожжи» имеет общий корень со словами «дрожь», «дрожать», которые применялись при описании вспенивания жидкости, зачастую сопровождающего брожение, осуществляемое дрожжами. Английское слово «yeast» (дрожжи) происходит от староанглийского «gist», «gyst», что означает «пена, кипеть, выделять газ»[3].
Дрожжи, вероятно, одни из наиболее древних «домашних организмов». Тысячи лет люди использовали их для ферментации и выпечки. Археологи нашли среди руин древнеегипетских городов жернова и пекарни, а также изображение пекарей и пивоваров. Предполагается, что пиво египтяне начали варить за 6000 лет до н. э., а к 1200 году до н. э. овладели технологией выпечки дрожжевого хлеба наряду с выпечкой пресного[4]. Для начала сбраживания нового субстрата люди использовали остатки старого. В результате в различных хозяйствах столетиями происходила селекция дрожжей и сформировались новые физиологические расы, не встречающиеся в природе, многие из которых даже изначально были описаны как отдельные виды. Они являются такими же продуктами человеческой деятельности, как сорта культурных растений.[5]
 Луи Пастер — учёный, установивший роль дрожжей в спиртовом брожении
Луи Пастер — учёный, установивший роль дрожжей в спиртовом броженииВ 1680 году голландский натуралист Антони ван Левенгук впервые увидел дрожжи в оптический микроскоп, однако не распознал в них из-за отсутствия движения живых организмов [6]. И лишь в 1857 году французский микробиолог Луи Пастер в работе «Mémoire sur la fermentation alcoholique» доказал, что спиртовое брожение — не просто химическая реакция, как считалось ранее, а биологический процесс, производимый дрожжами[7][8].
В 1881 году Эмиль Христиан Хансен, работник лаборатории датской компании 1883 году впервые использовал её для получения пива вместо нестабильных заквасок[4]. В конце XIX века при его участии создаётся первая классификация дрожжей, в начале XX века появляются определители и коллекции дрожжевых культур. Во второй половине века наука о дрожжах (зимология) помимо практических вопросов начинает уделять внимание экологии дрожжей в природе, цитологии, генетике.
До середины XX века учёные наблюдали только половой цикл аскомицетных дрожжей и рассматривали их всех как обособленную таксономическую группу сумчатых грибов. Японскому микологу Исао Банно в 1969 году удалось индуцировать половой цикл размножения у Rhodotorula glutinis, которая является базидиомицетом. Современные молекулярно-биологические исследования показали, что дрожжи сформировались независимо среди аскомицетных и базидиомицетных грибов и представляют собой не единый таксон, а скорее жизненную форму. [9]
24 апреля 1996 года года было объявлено, что Saccharomyces cerevisiae стал первым эукариотическим организмом, чей геном (12 млн пар оснований) был полностью секвенирован[10]. Секвенирование заняло 7 лет, и в нём принимали участие более 100 лабораторий[11]. Следующим дрожжевым организмом и шестым эукариотом с полностью расшифрованным геномом в 2002 году стал Schizosaccharomyces pombe[12] с 13,8 млн пар оснований.
Аскомицетные и базидиомицетные дрожжи
Различить дрожжи, принадлежащие к разным отделам грибов можно как по характеристикам их жизненного цикла, так и без его наблюдения по признакам аффинитета. К ним относится: синтез каротиноидов (встречается только у базидиомицетных дрожжей), тип убихинонов (с 5—7 изопреноидными остатками у аскомицетных и с 8—10 у базидиомицетных, хотя есть исключения), тип почкования (см. раздел Жизненный цикл), содержание ГЦ пар в ДНК (26—48 % у аскомицетных, 44—70 % у базидиомицетных), наличие уреазы (характерна за несколькими исключениями только базидиомицетным) и др.
Типичное разделение
- Saccharomycotina
- Taphrinomycotina
- Schizosaccharomycetes
- Urediniomycetes
- Sporidiales
Особенности метаболизма
Дрожжи являются хемоорганогетеротрофами и используют органические соединения как для получения энергии, так и в качестве источника углерода. Им необходим кислород для дыхания, однако при его отстутствии многие виды способны получать энергию за счёт брожения с выделением спиртов (факультативные анаэробы). В отличие от бактерий, среди дрожжей нет облигатных анаэробов, гибнущих при наличии кислорода в среде. При пропускании воздуха через сбраживаемый субстрат дрожжи прекращают брожение и начинают дышать (поскольку этот процесс эффективнее), потребляя кислород и выделяя углекислый газ. Это ускоряет рост дрожжевых клеток (эффект Пастера). Однако даже при доступе кислорода в случае высокого содержания глюкозы в среде дрожжи начинают её сбраживать (эффект Кребтри).[13]
Дрожжи достаточно требовательны к условиям питания. В анаэробных условиях дрожжи могут использовать в качестве источника энергии только углеводы, причём в основном гексозы и построенные из них олигосахариды. Некоторые виды (Pichia stipitis, Pachysolen tannophilus) усваивают и пентозы, например, ксилозу. Schwanniomyces occidentalis и Saccharomycopsis fibuliger способны сбраживать крахмал, Kluyveromyces fragilis — инулин. В аэробных условиях круг усваиваемых субстратов шире: помимо углеводов также жиры, углеводороды, ароматические и одноуглеродные соединения, спирты, органические кислоты. Гораздо больше видов способно использовать пентозы в аэробных условиях. Тем не менее, сложные соединения (лигнин, целлюлоза) для дрожжей недоступны.
Источниками азота для всех дрожжей могут быть соли аммония, примерно половина видов имеет нитратредуктазу и может усваивать нитраты. Пути усвоения мочевины различны у аскомицетовых и базидиомицетовых дрожжей. Аскомицетовые сначала карбоксилируют её, затем гидролизуют, базидиомицетовые — сразу гидролизуют уреазой.
Для практического применения важны продукты вторичного метаболизма дрожжей, выделяемые в малых количествах в среду: сивушные масла, ацетоин (ацетилметилкарбинол), диацетил, масляный альдегид, изоамиловый спирт, диметилсульфид и др. Именно от них зависят органолептические свойства полученных с помощью дрожжей продуктов.[14]
Распространение
Местообитания дрожжей связаны преимущественно с богатыми сахарами субстратами: поверхностью плодов и листьев, где они питаются прижизненными выделениями растений, нектаром цветов, раневыми соками растений, мёртвой фитомассой и т. д., однако они распространены также в почве (особенно в подстилке и органогенных горизонтах) и природных водах. Дрожжи (р. Candida, Pichia, Ambrosiozyma) постоянно присутствуют в кишечнике и ходах ксилофагов (питающихся древесиной насекомых), богатые дрожжевые сообщества развиваются на листьях, поражённых тлёй. Представители рода Lypomyces являются типичными почвенными обитателями.
Жизненный цикл
Отличительной особенностью дрожжей является способность к вегетативному размножению в одноклеточном состоянии. При сопоставлении с жизненными циклами грибов это выглядит как почкование спор или зиготы. Многие дрожжи также способны к реализации полового жизненного цикла (его тип зависит от аффинитета), в котором могут быть и мицелиальные стадии.
У некоторых дрожжеподобных грибов, образующих мицелий, возможен его распад на клетки (артроспоры). Это роды Endomyces, Galactomyces, Arxula, Trichosporon. У последних двух артроспоры после образования начинают почковаться. Trichosporon также образует вегетативные эндоспоры внутри клеток мицелия.
Циклы аскомицетных дрожжей
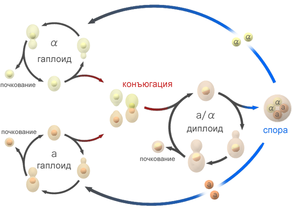 Жизненный цикл аскомицетных гапло-диплоидных дрожжей.
Жизненный цикл аскомицетных гапло-диплоидных дрожжей.Наиболее характерным типом вегетативного размножения для одноклеточных аскомицетных дрожжей является почкование, лишь Schizosaccharomyces pombe размножаются не почкованием, а бинарным делением[15]. Место закладки почки является важным диагностическим признаком: полярное почкование за счёт образования шрамов почкования приводит к формированию апикулярных (лимоновидных, Saccharomycodes, Hanseniaspora, Nadsonia) и грушевидных (Schizoblastosporion) клеток; многостороннее не видоизменяет форму клетки (Saccharomyces, Pichia, Debaryomyces, Candida). У родов Sterigmatomyces, Kurtzmanomyces, Fellomyces почкование происходит на длинных выростах (стеригмах).
Почкование у аскомицетных дрожжей голобластическое: клеточная стенка материнской клетки размягчается, выгибается наружу и даёт начало клеточной стенке дочерней.
Часто, особенно у аскомицетных дрожжей родов Candida и Pichia, клетки после почкования не расходятся и образуют псевдомицелий, отличающийся от истинного отчётливо видными перетяжками на месте септ и более короткими по сравнению с предшествующими конечными клетками.
Гаплоидные аскомицетные дрожжевые клетки имеют два типа спаривания: a и α. Термин «пол» не используется, поскольку клетки морфологически идентичны и различаются только одним генетическим локусом mat (от англ. mating — спаривание). Клетки разных типов у могут сливаться и образовывать диплоид a/α, который после мейоза даёт 4 гаплоидных аскоспоры: две a и две α. Вегетативное размножение аскомицетных дрожжей возможно у разных видов либо только на гаплоидной стадии, либо только на диплоидной, либо на обеих (гапло-диплоидные дрожжи).
Циклы базидиомицетных дрожжей
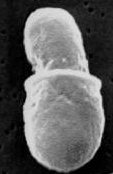 Энтеробластическое почкование у Malassezia sp.
Энтеробластическое почкование у Malassezia sp.Почкование базидиомицетных дрожжей энтеробластическое: клеточная стенка материнской клетки разрывается, из разрыва выходит почка и синтезирует свою клеточную стенку с нуля. Деление дрожжевых клеток для базидиомицетов не характерно.
Помимо обычного почкования многие виды исключительно базидиомицетных дрожжей (р. Sporidiobolus, Sporobolomyces, Bullera) способны образовывать вегетативные баллистоспоры: споры на выросте, наполненном гликогеном. Из-за гидролиза гликогена давление увеличивается и спора отстреливается на расстояние до нескольких миллиметров. При тесте на образование баллистоспор дрожжи высеваются на пластинку агаризованной питательной среды, закреплённую на крышке чашки Петри. Рост дрожжей на среде под этой пластинкой означает наличие у них баллистоспор и их принадлежность к базидиомицетам.
 Плодовое тело Tremella mesenterica.
Плодовое тело Tremella mesenterica.При половом размножении у базидиомицетов при слиянии гаплоидных дрожжевых клеток (плазмогамия) слияние ядер (кариогамия) не происходит и формируется дикариотическая клетка, дающая начало мицелию. Уже на мицелии происходит кариогамия и образуются базидиоспоры, часто даже на плодовом теле (порядок Tremallales). Единственными дрожжами среди базидиомицетов, не образующими мицелия даже при половом цикле размножения являются Xanthophyllomyces dendrorhus.
Следует отметить, что у базидиомицетовых дрожжей типы спаривания различаются обычно не одним, а большим количеством локусов. Могут сливаться только те клетки, у которых все эти локусы различны, то есть типов спаривания больше двух.
Типы спаривания
 Расположение неактивных локусов HML и HMR и активного mat на хромосоме III
Расположение неактивных локусов HML и HMR и активного mat на хромосоме IIIПри половом размножении дрожжей сливаться могут не любые 2 клетки, а только гаплоидные клетки разных типов спаривания. Существуют два типа таких клеток, которые различаются между собой по одному генетическому локусу, обозначаемому mat[16] (от английского mating). Локус может находится в одном из двух аллельных состояний: mat а и mat α. Mat а клетки синтезируют половые гормоны, которые дают сигнал α -клеткам. α -клетки отвечают а-клеткам, активируя мембранные рецепторы, которые воспринимают только феромоны от клетки противоположенного типа спаривания.[17] Поэтому слияние двух одинаковых клеток невозможно.
После слияния образуется диплоидная клетка с генотипом а/α, которой необходимо стать бесполой, чтоб больше не сливаться, и затем осуществить мейоз. Клетка этого добивается следующим образом. Ген mat а кодирует белок а1, который выполняет две функции: он подавляет считывание мРНК для белка α1 с гена mat α, поэтому фенотип α не развивается (не синтезируются α-феромоны), но он не мешает синтезу белка α2, который репрессирует а-специфичные гены, и фенотип а тоже не развивается. Во-вторых, белки а1 и α2 вместе активируют α/а-специфичные гены, которые необходимы для осуществления мейоза.
Дрожжи могут изменять свой тип спаривания с помощью рекомбинации ДНК. Это изменение у клеток происходит с частотой примерно 10-6 на клетку. Кроме локуса mat в клетке ещё имеется по копии генов mat а и mat α: соответственно HMR(Hidden MAT Right) и HML (Hidden MAT Left).[18] Но эти локусы находятся в молчащем состоянии. Клетка заменяет работающий локус mat на копию. При этом копия снимается с того локуса, который находится в противоположенном аллельном состоянии. За этот процесс отвечает ген НО. Этот ген активен только в гаплоидном состоянии. Он кодирует эндонуклеазы, которые разрезают ДНК в локусе mat. Затем экзонуклеазы убирают участок mat и на его место встает копия HMR или HML.[19]
Применение
Некоторые виды дрожжей с давних пор используются человеком при приготовлении хлеба, пива, вина, кваса и др. В сочетании с перегонкой процессы брожения лежат в основе производства крепких спиртных напитков. Полезные физиологические свойства дрожжей позволяют использовать их в биотехнологии. В настоящее время их применяют в производстве ксилита[20], ферментов, пищевых добавок, для очистки от нефтяных загрязнений.
Также дрожжи широко используются в науке в качестве модельных организмов для генетических исследований и в молекулярной биологии. Пекарские дрожжи были первыми из эукариот, у которых была полностью определена последовательность геномной ДНК. Важным направлением исследований является изучение прионов у дрожжей.
Традиционные процессы
Хлебопечение
 Гранулированные сухие активные дрожжи — коммерческий продукт для хлебопечения
Гранулированные сухие активные дрожжи — коммерческий продукт для хлебопеченияПриготовление печёного дрожжевого хлеба — одна из древнейших технологий. В этом процессе используется преимущественно Saccharomyces cerevisiae. Они проводят спиртовое брожение с образованием множества вторичных метаболитов, обуславливающие вкусовые и ароматические качества хлеба. Спирт испаряется при выпечке. Кроме того, в тесте формируются пузыри углекислого газа, заставляющие его «подниматься» и после выпечки придающие хлебу губчатую структуру и мягкость. Аналогичный эффект вызывает внесение в тесто соды и кислоты (обычно лимонной), но в этом случае не образуются вкусовые соединения.
Мука обычно бедна сбраживаемыми сахарами, поэтому в тесто добавляют яйца или сахар. Для получения большего количества вкусовых соединений тесто прокалывают или перемешивают, высвобождая углекислый газ, а потом снова оставляют «подниматься». Появляется, однако, риск, что дрожжам не хватит сбраживаемого субстрата.
Виноделие
 Ягоды винограда со слоем дрожжей на них.
Ягоды винограда со слоем дрожжей на них.Дрожжи в естественных условиях присутствуют на поверхности плодов винограда, часто они заметны как светлый налёт на ягодах, образованный преимущественно Hanseniaspora uvarum. Хотя «дикие» эпифитные дрожжи и могут привести к непредсказуемому результату брожения, обычно они не выдерживают конкуренции с обитающими в винных бочках бродильщиками.
Собранный виноград давят, получая сок (муст, виноградное сусло) с 10—25 % сахара. Для получения белых вин от него отделяют смесь косточек и кожуры (мезга), в мусте для красных вин она остаётся. Затем в результате брожения сахара превращаются в этанол. Вторичные метаболиты дрожжей, а также соединения, полученные из них при созревании вина определяют его аромат и вкус. Для получения ряда вин (например, шампанского) вторично сбраживают уже перебродившее вино.
Прекращение брожения связано либо с исчерпанием запасов сахаров (сухое вино), либо с достижением порога токсичности этанола для дрожжей. Хересные дрожжи, в отличие от обычных дрожжей (которые погибают, когда концентрации спирта в растворе достигает 12 %), более устойчивы. Первоначально хересные дрожжи были известны только на юге Испании (в Андалусии), где благодаря их свойствам получали крепкое вино — херес (до 24 % при длительной выдержке). Со временем хересные дрожжи были также обнаружены в Армении, Грузии, Крыму и др. Хересные дрожжи также используют при производстве некоторых крепких сортов пива.
Пивоварение и квасоварение
 Ячменный солод
Ячменный солодВ пивоварении в качестве сырья используется зерно (чаще всего ячмень), содержащее много крахмала, но мало сбраживаемых дрожжами сахаров. Поэтому перед брожением крахмал гидролизуют. Для этого используются амилазы, образуемые самим зерном при прорастании. Пророщенный ячмень носит название солод. Солод размалывают, смешивают с водой и варят, получая сусло, которое впоследствии сбраживается дрожжами. Различают пивные дрожжи низового и верхового брожения (эту классификацию ввёл датчанин Христиан Хансен).
Дрожжи верхового брожения (например, Saccharomyces cerevisiae) формируют «шапку» на поверхности сусла, предпочитают температуры 14—25°C (поэтому верховое брожение также называется тёплым) и выдерживают более высокие концентрации спирта. Дрожжи низового (холодного) брожения (Saccharomyces uvarum, Saccharomyces carlsbergensis) имеют оптимум развития при 6—10°C и оседают на дно ферментёра.
При создании пшеничного пива часто используется Torulaspora delbrueckii. При изготовлении ламбика применяются случайно попавшие в ферментёр дрожжи, обычно они принадлежат к роду Brettanomyces.
Квас производится по аналогичной схеме, однако помимо ячменного широко применяется ржаной солод. К нему добавляется мука и сахар, после чего смесь заливается водой и варится с образованием сусла. Важнейшим отличием квасоварения от производства пива является использование при сбраживании сусла помимо дрожжей молочнокислых бактерий.
Использование дрожжей в современной биотехнологии
Промышленное производство спирта
Спиртовое брожение — процесс, приводящий к образованию этанола (CH3CH2OH) из водных растворов углеводов (сахаров), под действием некоторых видов дрожжей (см. ферментация) как вид метаболизма.
В биотехнологии для производства спирта используют сахарный тростник, фуражную кукурузу и другие дешёвые источники углеводов. Для получения сбраживаемых моно- и олигосахаридов они разрушаются серной кислотой или амилазами грибного происхождения. Затем проводится сбраживание и ректификационная перегонка спирта до стандартной концентрации около 96 % об.[21]. Дрожжи рода Saccharomyces были генетически модифицированы для сбраживания ксилозы[22] — одного из основных мономеров гемицеллюлозы, что позволяет повысить выход этанола при использовании растительного сырья, содержащего наряду с целлюлозой и значительные количества гемицеллюлоз. Всё это может снизить цену и улучшить его положение в конкурентной борьбе с углеводородным топливом[23].
Пищевые и кормовые дрожжи
Дрожжи богаты белками, их содержание может доходить до 66 %, при этом 10 % массы приходится на незаменимые аминокислоты. Дрожжевая биомасса может быть получена на отходах сельского хозяйства, гидролизатах древесины, её выход не зависит от климатических и погодных условий. Поэтому её использование чрезвычайно выгодно для обогащения белками пищи человека и корма сельскохозяйственных животных. Добавление дрожжей в колбасы началось ещё в 1910-е годы в Германии, в 1930-е кормовые дрожжи начали производить в СССР, где эта отрасль особенно развилась.
В СССР первый крупный завод по производству белка — паприна, мощностью 70 000 т. в год, был пущен в 1973 году. В качестве сырья использовались отходы нефтепереработки. В 1980-е годы в СССР производилось 1 млн тонн микробного белка, в том числе и дрожжевого, что составляло 2/3 от общемировых объёмов, среди лидеров биотехнологического производства дрожжевого кормового белка и липофильно-жировых комплексов были ГДР и Венгрия.
Однако в 1990-е гг., в связи с возникшими гигиеническими и экологическими проблемами производства и применения микробного белка, а также с экономическим кризисом производство резко сократилось. Накопившиеся данные свидетельствовали о проявлении ряда отрицательных эффектов применения паприна в откорме птицы и животных. По экологическим и гигиеническим причинам снизился и интерес к данной отрасли и во всём мире.
Тем не менее на Западе сейчас производятся и продаются различные дрожжевые экстракты: вегемит, мармит, боврил, ценовис. Существуют подобные производства и в России, но их объёмы невелики[24]. Для получения экстрактов используются либо автолизаты дрожжей (клетки разрушаются и белок становится доступным благодаря ферментам самих клеток), либо их гидролизаты (разрушение специальными веществами). Они применяются как пищевые добавки и для придания блюдам вкусовых качеств; кроме того, существуют косметические средства на основе дрожжевых экстрактов.
Продаются также дезактивированные (убитые тепловой обработкой), но не разрушенные пищевые дрожжи, особенно популярные у веганов из-за высокого содержания белка и витаминов (особенно группы B), а также малого количества жиров. Некоторые из них обогащены витамином B12 бактериального происхождения.
Применение в медицине
- Высушенные пивные дрожжи используют для производства лекарственных препаратов и БАД.
- Длительное время выпускался препарат Гефефитин, как общеукрепляющее лекарственное средство.
- Жидкие пивные дрожжи традиционно прописывались ослабленным, лицам с аллергическими заболеваниями
- Существует ряд препаратов на основе Saccharomyces boulardii, поддерживающих и восстанавливающих флору желудочно-кишечного тракта. Показано, что S. boulardii снимает симптомы острой диареи у детей[25][26], предотвращает реинфекцию Clostridium difficile[27], снижает частоту сокращений мускулатуры кишечника у больных синдромом раздражённого кишечника[28], снижает риск возникновения различных видов диареи[29][30][31].
Применение в качестве модельного объекта
Многие данные по цитологии, биохимии и генетике эукариот были впервые получены на дрожжах рода Saccharomyces. Особенно это положение касается биогенеза митохондрий: дрожжи оказались одними из немногих организмов, способных существовать только за счёт гликолиза и не гибнущих в результате мутаций в геноме митохондрий, препятствующем их нормальному развитию[32]. Для генетических исследований важен короткий жизненный цикл дрожжей и возможность быстрого получения большого числа их особей и поколений, что позволяет изучать даже очень редкие явления.
В настоящее время интенсивно ведётся изучение прионов дрожжей, поскольку те близки по строению к открытым ранее прионам млекопитающих, однако абсолютно безопасны для человека[33][34]; их также существенно проще исследовать.
Чайный гриб
 Чайный гриб в банке
Чайный гриб в банкеЧайный гриб является ассоциацией дрожжей и уксуснокислых бактерий. Наиболее часто наблюдались ассоциации дрожжей Brettanomyces bruxellensis, Candida stellata, Schizosaccharomyces pombe, Torulaspora delbrueckii, Zygosaccharomyces bailii и других, с рядом штаммов семейства Acetobacteraceae[35]. Его использование в Российской империи началось в 1900-е годы, видимо, он был завезен после русско-японской войны.
В 50-е годы XX века в СССР активно исследовали различные природные вещества для их медицинского использования. В брошюре «Чайный гриб и его лечебные свойства» (Г. Ф. Барбанчик, 1954) отмечены антимикробные и противоатеросклеротические свойства зооглеи чайного гриба и его культуральной жидкости.
Коммерческие продукты, продаваемые под названием «сухие дрожжи»
В состав таких дрожжей входят не только клетки микроорганизмов, но и минеральные добавки, некоторые ферменты.
Дрожжи как фактор порчи пищевых продуктов
Дрожжи способны расти на средах с низкими pH (5,5 и даже ниже), особенно в присутствии углеводов, органических кислот и других легко утилизируемых источников органического углерода[36]. Они хорошо развиваются при температурах 5—10°С, когда мицелиальные грибы уже неспособны к росту.
В процессе жизнедеятельности дрожжи метаболизируют компоненты пищевых продуктов, образуя собственные специфические конечные продукты метаболизма. При этом физические, химические и, как следствие, органолептические свойства продуктов изменяются — продукт «портится»[37]. Разрастания дрожжей на продуктах нередко видны невооруженным глазом как поверхностный налёт (например, на сыре или на мясных продуктах) или проявляют себя, запуская бродильный процесс (в соках, сиропах и даже в достаточно жидком варенье).
Дрожжи рода Zygosaccharomyces уже долгое время являются одними из важнейших агентов порчи продукции пищевой промышленности. Особенно затрудняет борьбу с ними тот факт, что они могут расти в присутствии высоких концентраций сахарозы, этанола, уксусной кислоты, бензойной кислоты и диоксида серы[38], являющихся важнейшими консервантами.
Патогенные дрожжи
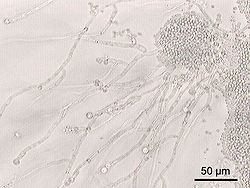 Candida albicans, образующая скопления дрожжевых клеток и псевдомицелий
Candida albicans, образующая скопления дрожжевых клеток и псевдомицелийНекоторые виды дрожжей являются факультативными и условными патогенами, вызвая заболевания у людей с ослабленной иммунной системой.
Дрожжи рода Candida являются компонентами нормальной микрофлоры человека, однако при общем ослаблении организма травмами, ожогами, хирургическим вмешательством, длительном применении антибиотиков, в раннем детском возрасте и в старости и т. д. грибы рода кандида могут массово развиваться, вызывая заболевание — кандидоз. Существуют различные штаммы этого гриба, в том числе достаточно опасные. В нормальных условиях в человеческом организме дрожжи рода Candida ограничиваются в своём развитии естественной бактериальной микрофлорой человека (лактобактерии и пр.), но при развитии патологического процесса многие из них образуют высокопатогенные сообщества с бактериями.
Cryptococcus neoformans вызывает криптококкоз, особенно опасный для ВИЧ-инфицированных людей: среди них заболеваемость криптококкозом достигает 7—8 % в США и 3—6 % в Западной Европе. Клетки C. neoformans окружены прочной полисахаридной капсулой, которая препятствует их распознаванию и уничтожению лейкоцитами. Дрожжи этого вида наиболее часто обнаруживаются в помёте птиц, при том что сами птицы не болеют.
Род Malassezia включает облигатных симбионтов теплокровных животных и человека, не встречающихся нигде, кроме их кожных покровов. При нарушениях иммунитета вызывают питириаз (пёстрый лишай), фолликулит и себорейный дерматит. У здоровых людей при нормальном функционировании сальных желез Malassezia никак себя не проявляют и даже играют положительную роль, препятствуя развитию более опасных патогенов.
См. также
Примечания
В Викисловаре есть статья «дрожжи» - Yeast growth and the cell cycle
- Yeast virtual library
- Science of Breadmaking
- Ancient Egyptian Bread Making
- Brewing Yeast Propagation and Maintenance: Principles and Practices
- Учебное пособие "Биология дрожжей"
Wikimedia Foundation. 2010.