- Насекомые
-
Насекомые 
Божья коровка CoccinellaНаучная классификация промежуточные рангиДомен: Эукариоты Царство: Животные Подцарство: Эуметазои Без ранга: Двусторонне-симметричные Без ранга: Первичноротые Без ранга: Линяющие Без ранга: Panarthropoda Тип: Членистоногие Подтип: Трахейнодышащие Надкласс: Шестиногие Класс: Насекомые Международное научное название Подклассы и инфраклассы: - Бескрылые насекомые (Apterygota)
- Monocondylia
- Крылатые насекомые (Pterygota)
- Открыточелюстные (Ectognatha, устаревший)
- Щетинохвостковые (Triplura)
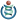
Систематика
на Викивидах
Изображения
на ВикискладеITIS 99208 NCBI 50557 EOL 344 Насеко́мые (лат. Insecta, Insecta-Ectognatha, Ectognatha) — класс (по другой классификации — надкласс[1]) беспозвоночных членистоногих животных. Вместе с многоножками относятся к подтипу трахейных[2]. Обладают наибольшим разнообразием среди всех остальных животных на Земле; включают бабочек, жуков, мух, муравьёв, пчёл и других. Наука, изучающая насекомых, называется энтомологией.
Насекомые — самый многочисленный класс живых существ на Земле, занимающий всевозможные экологические ниши. Произошло это предположительно потому, что в их развитии получили приоритет три принципа[3]:
- Предельная простота организации;
- Узкая жизненная специализация;
- Необычайная плодовитость.
Содержание
Этимология названия
Русское слово насекомое, первоначально «(животное) насеченное, с насечками», которые отделяют один членик от другого, калькировано в XVIII веке с фр. insecte, произошедшего от лат. insectum (страдательное причастие от глагола inseco «разрезаю, надрезаю, рассекаю»). Латинское слово, в свою очередь, было скалькировано с др.-греч. ἔντομον (< ἐντομή «надрез, зазубрина, насечка» < τέμνω «режу»)[4]. Слово насекомое впервые упоминается в памятниках русского языка в XVIII веке, в словарях фиксируется с 1731 года[5].
Общая характеристика
Кратко класс можно охарактеризовать следующим образом. Это наземные членистоногие, у которых тело явно разделено на голову, грудь и брюшко, а основные, служащие для передвижения, конечности находятся в числе 3 пар на грудном отделе. Брюшко у взрослых особей несёт видоизменённые остатки конечностей — грифельки и др., лишь у отряда бессяжковых сохранились слабые брюшные ноги. Дыхание осуществляется с помощью трахейной системы или кожное — всей поверхностью тела. Те насекомые, которые дышат с помощью всей поверхности тела и имеют проницаемые для дыхательных газов и испарения покровы, ограничены в своём распространении только влажными средами. Они встречаются в основном в почве и в гниющих остатках организмов. Те высшие насекомые, у которых покровы непроницаемы и развита трахейная система, могут обитать и в сухих средах. Эти насекомые широко расселились по нашей планете. Именно такие, ведущие открытый образ жизни и часто ярко окрашенные высшие насекомые и известны большинству читателей. Встречаются повсеместно, включая Антарктиду (Belgica antarctica).
Численность
Оценка числа описанных видов насекомых варьируется от примерно 720 тыс.[6] до более чем 1 млн видов[7]. Оценки общей истинной численности насекомых широко варьируются от примерно 2 млн[8], 5—6 млн[9] до около 8 млн[10] видов.
Расчёты, основанные на экстраполяции количества видов жесткокрылых и чешуекрылых в Новой Гвинее показывают, что предположительное число видов насекомых может находиться в пределах 3,7—5,9 млн видов[2]. Насекомые очень разнообразны и большинство их видов плохо изучено, поэтому истинная оценка количества существующих видов является очень затруднительной. Некоторые из описанных видов известны только по находкам из одной-единственной местности или даже по единственному экземпляру[11].
Размеры
См. также: Крупнейшие членистоногиеРазмеры представителей группы колеблются в широких пределах. Крупнейшим насекомым считается палочник Phobaeticus chani[12]. Наиболее крупный экземпляр был обнаружен в 1989 году в лесах острова Калимантан и с вытянутыми конечностями его длина составляет 567 мм[13]. Предыдущим рекордсменом по длине тела был палочник Phobaeticus kirbyi, максимальная длина тела которого составляет 328 мм, а вытянутыми конечностями — 546 мм[14].
Самым крупным жуком в мире считается дровосек-титан (Titanus giganteus) из Южной Америки, достигающий длины до 167 мм[15], а по некоторым неподтверждённым источникам даже до 210 мм[16]. Вторым по величине является южноамериканский жук-геркулес (Dynastes hercules), отдельные особи самцов которого достигают длины 160—165 мм[17].
Южноамериканская совка тизания агриппина (Thysania agrippina) является крупнейшей по размаху крыльев бабочкой в мире — в 1934 году в Бразилии была поймана самая крупная особь с размахом крыльев 30,8 см[18]. Самой крупной по размаху крыльев дневной бабочкой является орнитоптера королевы Александры (Ornithoptera alexandrae) — размах крыльев самки достигает 28 см[18]. Самки сатурнии Coscinocera hercules, обитающей в Австралии и Новой Гвинее, обладают самой большой площадью крыльев — до 263,2 см², а их размах может достигать 28 см. Одной из самых больших бабочек в мире также является сатурния Attacus atlas, крупнейший задокументированный экземпляр которой был пойман на острове Ява и имел размах крыльев 26,2 см[19].
Одна из самых маленьких бабочек в мире — микро-моль Stigmella ridiculosa (семейство Nepticulidae) с Канарских островов имеет размах крыльев всего около 2 мм[20]. Самым маленьким жуком является перокрылка Nanosella fungi из США, длина тела которой около 0,25 мм, что почти в 800 раз меньше, чем у самого большого жука[21].
Наружное строение
 Морфология насекомого
Морфология насекомого
A — голова, B — грудь, C — брюшко
1. антенна,
2. глазки (нижний),
3. глазки (верхний),
4. сложный глаз,
5. мозг (cerebral ganglia),
6. переднегрудь,
7. дорсальная артерия,
8. трахеи,
9. среднегрудь,
10. заднегрудь,
11. переднее крыло,
12. заднее крыло,13. средняя кишка (желудок),
14. сердце,
15. яичник,
16. задняя кишка (кишечник, прямая кишка и анальное отверстие),
17. анус,
18. вагина,
19. абдоминальный ганглий,
20. мальпигиевы сосуды,
21. подушечка,22. коготки,
23. лапка,
24. голень,
25. бедро,
26. вертлуг,
27. fore-gut (crop, gizzard),
28. грудной ганглий,
29. тазик,
30. слюнная железа,
31. подглоточный ганглий,
32. ротовой аппаратПокрывающее тело кутикула является частью кожи, образуя плотный наружный экзоскелет, но в ряде случаев она мягкая и тонкая. Наружная кутикула подразделена на отдельные щитки — склериты (sclerites) и вследствие своей плотности благоприятна для развития на ней различных образований: вдавлений, бороздок, бугорков, рёбрышек, мелких волосков -хетоидов и др. К кутикуле причленяются также подвижные кожные волоски — хеты, имеющие иногда характер щетинок или чешуек.
Окраска тела и его частей очень разнообразна, зависит от пигментов, располагающихся в кутикуле или подстилающей её гиподерме, либо обусловлена оптическими явлениями, связанными со структурой кутикулы. Ввиду большой стойкости оптическая и кутикулярная пигментная окраски остаются почти неизменными после смерти; гиподермальная же пигментная, напротив, быстро разрушается. Нередко пигмент образует вдоль или поперёк тела полосы, в последнем случае они называются перевязями (fasciae). Часто это связано с явлением мимикрии.
Тело насекомых подразделяется на три сегментированных отдела: голова, грудь и брюшко. Каждый сегмент подразделяется на четыре части — верхнее полукольцо называется тергит, нижнее полукольцо называется стернит, боковые стенки — плейриты. При обозначении взаимного расположения частей тела и органов терминами «дорсальный» (dorsalis) обозначают верхнюю сторону тела, а «вентральный» (ventralis) — нижнюю сторону. Также выделяют мезосому (у муравьёв из трёх грудных сегментов и первого абдоминального сегмента проподеума) и метасому (стебелёк и брюшко).
Голова
Голова (caput) внешне нечлениста, но эволюционно произошла в результате слияния 5 сегментов в процессе олигомеризации тела. Сохранившимися конечностями этих сегментов являются усики, или антенны первые (antennae) и 3 пары ротовых челюстей — нечленистые верхние челюсти, или мандибулы (mandibulae), членистые нижние челюсти, или максиллы (maxillae) и членистая, внешне непарная нижняя губа (labium), являющаяся второй парой нижних челюстей, слившихся между собой. Различают несколько типов ротовых органов, из которых первичным является грызущий, предназначенный для разрывания и поглощения более или менее твёрдой пищи. В процессе эволюции возникло несколько модификаций этого исходного типа для потребления жидкой пищи, которые у разных сосущих групп насекомых устроены по-разному. В одних случаях сосание связано с проколом пищевого субстрата и возникает колюще-сосущий ротовой аппарат (клопы, тли, комары и др.), в других приём пищи не сопровождается проколом, как, например, у большинства бабочек. Особую модификацию представляет мускоидный тип ротового аппарата, возникший у мух и приспособленный к потреблению как жидкой, так и твёрдой пищи. Другой путь развития исходного ротового аппарата наблюдается у скрыточелюстных, нижняя губа которых слилась с т. н. оральными складками, образовав парные челюстные карманы, в которые погружены мандибулы и максиллы.
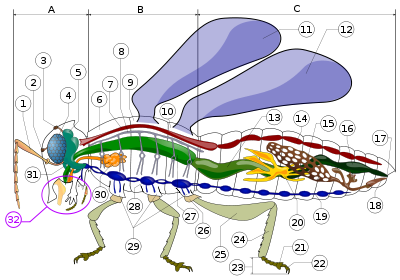
 Типы усиков насекомых
Типы усиков насекомыхТвёрдой основой головы является черепная коробка (epicranium). На голове различают переднюю поверхность — лоб (frons), который сверху переходит в темя (vertex) и далее назад — затылок (occiput). Спереди ото лба лежит хорошо обособленная пластинка — наличник (clypeus) и далее вперед (вниз) — верхняя губа (labrum), подвижный пластинчатый кожный выступ, прикрывающий сверху ротовые органы. На боках головы, под глазами, различают щёки (genae), сзади переходящие в виски (tempora), а снизу лежит горло (gula). С боков головы расположены сложные глаза (oculi), состоящие из множества зрительных единиц — омматидиев и являющиеся основными органами зрения. Кроме этого, между сложными глазами обычно расположено 1—3 простых глаз, или глазков (ocelli). В зависимости от биологии положение головы неодинаково. Различают гипогнатическую голову (caput hypognathum) — с ротовыми органами, обращёнными вниз, подобно ногам, и прогнатическую голову (caput prognathum) — с ротовыми органами, обращёнными вперёд. Первый тип обычно характерен для растительноядных, а второй — для хищных насекомых.
Усики сидят по бокам лба, между глазами или впереди них, нередко в хорошо обособленной усиковой впадине. Они очень разнообразны, характерны для разных групп насекомых. В своей основе усики состоят из утолщённого основного членика, называемого рукояткой (scapus), за которым следует ножка (pedicellus), и, начиная с третьего членика, располагается основная часть — жгутик (flagellum). Различают несколько типов усиков (см. рис.).
Грудь
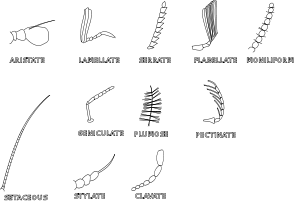
 Строение и типы конечностей насекомых.
Строение и типы конечностей насекомых.
Тип конечности: 1. Бегательная (жужелица), 2. Прыгательная (саранча), 3. Копательная (медведки), 4. Плавательная (плавунец), 5. Хватательная (богомол), 6. Собирательная (медоносная пчела).
Часть конечности: a. тазик, b. вертлуг, c. бедро, d. голень, e. лапка.Грудь (thorax) состоит из трёх сегментов — передне-, средне- и заднегруди (pro-, meso-, metathorax). Тергиты груди называются спинкой (notum), а стерниты — грудкой (sternum). Соответственно 3 сегментам груди различают передне-, средне- и заднеспинку (pro-, meso-, metanotum) и также передне-, средне и заднегрудку (pro-, meso-, metasternum). Каждый плейрит груди подразделяется швом, по крайней мере, на два склерита — спереди эпистерн (episternum) и сзади эпимер (epimerum). Прикрепление к груди органов движения превращает её в локомоторный центр тела, увеличивает размеры за счёт развития мощной мускулатуры и сильному изменению и усложнению описанных выше склеритов.
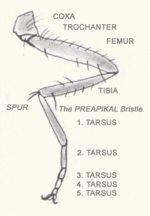 Нога насекомого
Нога насекомогоНоги (pedes) причленены к груди снизу, обычно сидят в тазиковых впадинах и состоят из тазика (coxa), вертлуга (trochanter), бедра (femora), голени (tibia) и лапки (tarsus). Тазик и вертлуг обеспечивают необходимую подвижность ноге. В некоторых случаях вертлуг состоит из двух члеников. Бедро является самой крупной и сильной частью ноги, так как имеет мощную мускулатуру. Его сочленение с голенью называют коленным, а прилегающую к нему часть — коленом (geniculus). Голень по длине примерно равна бедру, но тоньше его, снабжена шипами (spinae), а на вершине — шпорами (calcariae). Лапка обычно расчленена, состоит из 2-5 члеников, на вершине несёт пару коготков (unguiculi), между которыми располагается широкая присоска — аролий (arolium) или узкий эмподий (empodium). Соответственно образу жизни ноги подверглись разнообразной специализации, поэтому различают несколько их типов.
Крылья
Крылья насекомых (alae) представляют собой пластинчатые выросты покровов, связанные с комплексом обслуживающих их мышц и преобразованных склеритов груди. В типичном случае их две пары: передняя, прикреплённая к среднегруди, и задняя — на заднегруди. Крылья состоят из тонкой крыловой пластинки (membrana), укреплённой на твёрдых жилках (venae), играющих роль прочной основы крыла. Жилки делятся на продольные и поперечные, образующие у более низкоорганизованных групп густую сеть и многочисленные ячейки (cellulae). Поперечные жилки у высших форм сокращаются в числе и иногда исчезают совсем, так как главную поддерживающую роль несут продольные, попеременно выпуклые и вогнутые жилки. Поэтому продольные жилки, хотя и претерпели у высших групп значительную эволюцию, всё же сохранили свою основу.
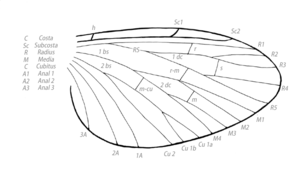 Жилкование
ЖилкованиеВ исходном состоянии различают следующие продольные жилки: костальная (costa или C); субкостальная (subcosta, Sc); радиальная (radius, R), снабжённая сзади ветвью — радиальным сектором (radii sector, RS); срединная, или медиана (media, M), иногда делится на переднюю (MA) и заднюю (MP) ветви; кубитальная (cubitus, Cu), в числе двух, нередко сближенных жилок — передней (CuA) и задней (CuP); анальные (analis, A) в числе 1—4. В некоторых случаях на задних крыльях позади анальных располагается ещё серия югальных (jugalis, Ju) жилок, входящих вместе с анальными в состав складывающегося веера (vannus) крыла. Между продольными жилками располагается серия полей (campo), называемых по пробегающей спереди жилке: костальное, срединное и т. д. Более крупными подразделениями являются области (regio) крыла: главная — спереди от анальных жилок, анальная — между анальными жилками, югальная — между югальными.
Этот исходный план жилкования подвергается существенным модификациям путём образования ряда ветвей или выпадения отдельных жилок, их перемещения, слияния и т. д. Наиболее существенно в эволюции крыльев появление у новокрылых насекомых югальной области, обеспечившей особое складывание крыльев вдоль тела. Однако, в процессе дальнейшей эволюции Neoptera югальная область и способность складывать крылья вдоль тела неоднократно утрачивались в разных группах. Также можно отметить происходивший процесс костализации крыла, то есть смещения всех продольных жилок к переднему краю крыла, укреплявшего механическую основу и повышавшего аэродинамические характеристики. Наивысшей степени костализация достигла у перепончатокрылых и двукрылых. Многие отряды пошли по другому пути: у них летательная функция целиком переместилась на задние крылья, а передние превратились в плотные, ороговевшие надкрылья (elytrae), играющие роль покрышек для нежных задних крыльев. В ряде случаев надкрылья полностью утратили жилкование (жуки, кожистокрылые).
С совершенствованием полёта от относительной независимости крылья насекомых «переходили» к сцеплению друг с другом различными способами, функционируя как единый орган с перемещением основной нагрузки на первую пару. Другими словами, происходил процесс диптеризации (от лат. названия отряда двукрылых). С дальнейшей эволюцией задняя пара сокращается в размерах, а затем и утрачивается. Возникает высший этап полёта насекомых — морфологическая двукрылость. Всё сказанное определяет выдающуюся роль строения крыльев в классификации и понимании эволюции насекомых.
Брюшко
Брюшко (abdomen) состоит из многих, в целом однотипных, сегментов, исходно из 10, не считая хвостового компонента — тельсона, но в таком виде оно есть лишь у некоторых первичнобескрылых и зародышей. Нередко число сегментов сокращается до 5—6 и менее. На VIII и IX сегментах находятся половые отверстия и очень часто — наружные половые придатки, поэтому эти сегменты обычно называют генитальными. Прегенитальные сегменты у взрослых, как правило, лишены придатков, а постгенитальные имеют развитый лишь тергит X сегмента, тогда как XI сегмент редуцирован и свойственные ему придатки — церки (cerci) переместились на X сегмент. Строение церков весьма разнообразно, а у высших форм они атрофированы. Остатками постгенитальных сегментов являются лежащие дорсально вокруг анального отверстия склериты — сверху анальная пластинка (epiproctus), по бокам и снизу нижние анальные створки (paraproctes). Иногда (тараканообразные, уховертки) анальной пластинкой называют именно X тергит. Придатками генитальных сегментов являются у самца грифельки (styli) — на IX стерните, у самки — яйцеклад (oviductus) — парные выросты генитальных сегментов, являющиеся видоизмененными конечностями. IX стернит самца образует гипандрий, или генитальную пластинку (hypandrium), но нередко генитальной пластинкой именуют последний видимый стернит вообще, который у самок некоторых Polyneoptera может быть VIII или даже VII. У высших групп все эти части подвергаются редукции или модификации (так, у жалящих перепончатокрылых яйцеклад превращён в жало, хотя используется и по прямому назначению тоже). При редукции настоящего яйцеклада у некоторых групп (например, у многих жуков) возникает вторичный, телескопический яйцеклад из сильно уменьшенных в диаметре вершинных сегментов брюшка.
На конце брюшка самца располагается копулятивный аппарат, имеющий сложное и невероятно разнообразное строение у разных отрядов. Обычно в его составе имеется непарная часть — пенис (penis), имеющий сильно склеротизированную концевую часть — эдеагус (aedeagus). Строение копулятивного аппарата имеет важнейшее значение в систематике, так как его строение заметно различается даже у видов-двойников; часто его изучение позволяет решить труднейшие вопросы классификации родов, семейств и пр.
Внутреннее строение
Дыхательная система
Дыхательная система большинства насекомых представлена множеством трахей, пронизывающих всё тело. Трахеи ветвятся и опутывают все внутренние органы. Концевые ветви трахей заканчиваются звёздчатой трахейной клеткой, от которой отходят тончайшие трахейные трубочки (трахеолы), проникающие даже внутрь клеток окружающих тканей. Трахейная система насекомых является открытой (свободно сообщающейся с окружающим воздухом), лишь у некоторых личинок, живущих в воде, имеется закрытая система. В случае закрытой системы снабжение трахейной полости кислородом осуществляется с помощью трахейных жабр, улавливающих кислород, растворённый в воде.
Трахейная система насекомых выполняет не только роль снабжения организма кислородом, но и транспортную функцию по доставке кислорода к тканям. Необходимость наличия трахейной системы, пронизывающей всё тело, не позволяет насекомым достигать крупных размеров (по сравнению с позвоночными, например).
Кровеносная система
Кровеносная система насекомых незамкнутая. Гемолимфа только частично заключена в орган кровообращения — спинной сосуд. В остальном же она заполняет полость тела (миксоцель) и промежутки между органами, омывая их.[Л 1]
Главная функция гемолимфы — снабжение органов питательными веществами и удаление из них продуктов обмена. Гемолимфа также содержит гормоны, выделяемые железами внутренней секреции и участвующие в регуляции физиологических процессов. Дыхательная функция гемолимфы незначительна и ограничена растворённым в ней кислородом. Исключение составляет только кровь личинок комаров-звонцов, которая содержит переносчик кислорода, близкий к гемоглобину. Гемолимфа также создаёт внутреннее давление в организме насекомого, поддерживая форму тела у мягкотелых (например, личинок). Давление гемолимфы используется для осуществления некоторых видов механического перемещения, например, расправление крыльев, развёртывание хоботка, разрыв покровов при линьке. У некоторых насекомых (саранчовые, кузнечики) гемолимфа выбрызгивается в целях самозащиты (автогеморрагия) или содержит биологически активные вещества и выделяется наружу при опасности (нарывные жуки).[Л 2]
Важна также функция иммунитета, которую выполняет гемолимфа.[Л 2] Иммунная система насекомых включает яды и антибиотики плазмы (например, кантаридин и насутин), белковые ферменты плазмы (лизоцим), а также амёбоидные гемоциты, которые осуществляют фагоцитоз одноклеточных паразитов и образуют гемоцитарную капсулу вокруг многоклеточных. У насекомых нет специфического иммунитета с образованием антител по типу иммунитета позвоночных. Однако у некоторых насекомых наблюдались случаи кратковременного специфического иммунитета, где роль антител играли вещества небелковой природы.[Л 3]
Спинной сосуд представляет собой мышечную трубку, расположенную в перикардиальном синусе и подвешенную на коротких тяжах к спинной стенке тела. Задний отдел спинного сосуда — сердце, состоящее из нескольких соединённых последовательно пульсирующих камер. Передний отдел (аорта) имеет вид простой трубки. Каждая камера сердца имеет по два входных отверстия (устьица или остии) с клапанами, через которые гемолимфа поступает внутрь из полости тела. В отверстиях между камерами сердца также имеются клапаны, обеспечивающие движение гемолимфы в одном направлении. Задний конец сердца обычно закрыт.[Л 1]
В результате пульсаций камер сердца осуществляется движение гемолимфы в задне-переднем направлении. При диастоле (расширении камеры) гемолимфа входит в неё через устьица, а при систоле (сокращении) нагнетается вперёд, в аорту.[Л 1] Аорта открывается отверстием в полость головы, куда изливается гемолимфа. Движению гемолимфы способствует также сокращение диафрагм. Сокращение верхней диафрагмы увеличивает емкость околосердечной полости, куда устремляется гемолимфа в момент диастолы. Сокращение нижней диафрагмы способствует передвижению крови в полости тела спереди назад. Таким образом, совместная работа сердца и диафрагм осуществляет циркуляцию гемолимфы в организме насекомого: по спинному сосуду — сзади вперёд, в полости тела — спереди назад. Движение гемолимфы в придатках тела (усики, ноги, крылья, хвостовые придатки) осуществляется дополнительными (местными) органами. Обычно это либо пульсирующие ампулы (например, в основаниях усиков) или подвижные мембраны (в ногах).[Л 4] Число камер сердца — до восьми. Число сокращений сердца зависит от вида насекомого, его физического состояния, фазы развития и колеблется в пределах от 15 до 150 циклов в минуту.[Л 4]
Гемолимфа насекомых состоит из жидкой плазмы и клеточных элементов — гемоцитов. Плазма либо бесцветная, либо окрашена в желтоватый или зеленоватый цвет. Кровь личинок комаров-звонцов красноватого цвета из-за наличия переносящего кислород пигмента, родственного гемоглобину. Содержание воды в гемолимфе 75-90 %. Плазма содержит неорганические соли (Na+, K+, Ca2+, Mg2+, Cl-, PO43-, CO32-), питательные вещества, ферменты, гормоны, пигменты. Реакция гемолимфы слабокислая или нейтральная (pH = 6-7).[Л 4] Солёность гемолимфы соответствует 0,7-1,3 % раствору хлорида натрия.[Л 5]
Гемоциты имеют мезодермальное происхождение. Бывают различной формы, могут быть подвижными и неподвижными. Содержание гемоцитов зависит от вида насекомого. Например, у божьей коровки до 80 тыс. гемоцитов в 1 мм³ крови.[Л 4] У других насекомых — от 10 до 100 тыс.[Л 5]
Различают три вида гемоцитов:[Л 4]
- хромофильные лейкоциты (амёбоциты);
- гемоциты с зернистой плазмой;
- гемоциты с гомогенной плазмой (фагоциты).
При повреждении покровов у многих насекомых гемолимфа свёртывается и образует сгусток. При этом сначала происходит коагуляция плазмы, затем — агглютинация (склеивание) гемоцитов.[Л 6]
Нервная система
Нервная система осуществляет регуляцию процессов, происходящих в организме насекомого с помощью передачи нервных импульсов по рефлекторным дугам и выработки нейрогормонов некоторыми нейронами.
Центральная нервная система представляет собой цепь ганглиев, соединённых коннективами и состоит из так называемого мозга, подглоточного ганглия и брюшной нервной цепочки. Мозг представляет собой три сросшиеся ганглия и соответственно делится на три отдела: протоцеребрум, отвечающий за зрение и сложные поведенческие реакции, дейтоцеребрум (антенны) и тритоцеребрум (нижняя губа, мышцы вокруг ротового отверстия, внутренние органы). От мозга отходят две коннективы, обнимающие глотку сверху вниз. Дальнейшие ганглии лежат таким образом под пищеварительным трактом. Подглоточный ганглий состоит из трёх сросшихся ганглиев и регулирует работу челюстей, гипофаринкса и слюнных желёз. Грудные ганглии развиты сильнее брюшных, так как берут на себя регуляцию работы ног и крыльев. Брюшные ганглии прогрессивных форм олигомеризуются.
Вегетативная нервная система состоит из стоматогастрической системы, влияющей на переднюю кишку и сердце и туловищного отдела — непарного нерва между коннективами ганглий, влияющего не работу трахей, дыхалец и скелетных мышц.
Стоматогастрическая система представляет собой соединённый с тритоцеребрумом фронтальный ганглий перед мозгом, соединённый с ним возвратным нервом затылочный ганглий за мозгом, а также кардиальные и прилежащие тела. Кардиальные тела осуществляют секрецию своих гормонов, а также накопление гормонов нейросекреторных клеток мозга. Основная функция прилежащих тел — синтез ювенильных гормонов.
Выделительная система
Наиболее примитивными органами выделения являются лабиальные органы щетинохвосток. Это гомологи метамерных органов кольчатых червей, а также антеннальных и максиллярных желёз ракообразных, лабиальные органы имеются также у ногохвосток.
Органы парные, состоят из парных начального мешочка, извитого лабиринта, прямого выводного протока и единого выводного отверстия над основанием нижней губы. В выводной проток впадает железистый придаток.
У более совершенных групп насекомых основным органом выделения являются мальпигиевы сосуды. Кроме того, в выделении играют роль мочевые клетки жирового тела, накапливающие мочевую кислоту либо чтобы хранить вечно, либо чтобы потом с выходом клеток в плавание по гемолимфе передать мальпигиевым сосудам.
У некоторых насекомых (тутовый шелкопряд, гладыши) выделительную функцию выполняют также некоторые клетки гиподермы, скидываемые во время линьки.
Биология и экология
Жизненный цикл
Насекомые являются первично яйцекладущими животными; у немногих видов известно яйцеживорождение. После завершения развития в яйце (эмбриогенеза) происходит вылупление. С этого момента насекомое вступает во второй этап своего развития — фазу личинки, с которой и начинается послезародышевый период. Постэмбриональное развитие состоит в росте тела личинки, что сопровождается периодическими линьками — сбрасыванием старой кутикулы. В конце концов, происходит превращение во взрослое насекомое — имаго. У одних насекомых это происходит с последней линькой личинки, у других — посредством особой фазы развития, называемой куколкой. Говорят, что насекомые имеют разный метаморфоз. Личинки насекомых весьма разнообразны, но могут быть сведены к 2 основным типам — сходным со взрослой особью и несходным; первые называются нимфами, вторые — истинными личинками. Нимфы сходны со взрослой стадией как морфологически, так и биологически: они живут обычно в той же среде, что и имаго, и сходно питаются. Истинные личинки резко отличаются от взрослых особей внешним видом, средой обитания и пищей. Именно таким личинкам для превращения во взрослое насекомое требуется стадия куколки.
Имаго является заключительной стадией жизненного цикла, но далеко не всегда переход в эту стадию завершает развитие особи; это наблюдается лишь в немногих случаях, когда насекомое сразу по достижении состояния имаго является вполне половозрелым и способным к размножению (подёнки, шелкопряды и др.). Биологические функции (питание, расселение и размножение) распределяются между личинкой и имаго различным образом, при этом функция размножения почти всегда принадлежит имаго (кроме случаев неотении).
Весь жизненный цикл, называемый генерацией, протекает у разных видов в течение неодинакового времени. Многие виды имеют короткую продолжительность генерации дают одну, две или три генерации в один год и называются, соответственно, одно-, дву- и трёхгенерационными. У других одна генерация продолжается много лет (так, у чёрного таракана развитие длится 4 года, а у семнадцатилетней цикады — соответственно, 17 лет!). Другой существенной стороной жизненного цикла видов является время протекания в природе тех или иных фаз развития. Так, зимовка видов, обитающих в умеренном поясе может проходить на стадии яйца, личинки, куколки либо имаго; соответственно этому меняется время протекания других фаз в течение вегетационного периода. Следовательно, виды могут отличаться друг от друга не только количеством генераций в году, но и временем протекания отдельных стадий развития, то есть годичным циклом.
Иногда годичный цикл осложняется задержкой развития той или иной фазы — диапаузой, сопровождающейся понижением обмена веществ и прекращением питания. Внешне создается впечатление остановки развития. Диапауза контролируется как внешними (температура, длина светового дня и др.), так и внутренними (гормональными) факторами, являясь одной из форм приспособления насекомых к жизни в странах с резкими сезонными изменениями климата. Многие диапазирующие насекомые при вынужденном развитии без диапаузы (например, в лабораторных условиях) испытывают нарушения онтогенеза или погибают.
Питание

Пищевая избирательность проявляется у насекомых сложно и многообразно, путём использования для питания самых различных источников органического вещества: тканей животных и растений, продуктов жизнедеятельности организмов, мертвых тел и остатков животного и растительного происхождения. Приспособление к постоянному питанию каким-либо из этих видов органики породило среди насекомых различные пищевые режимы, или пищевую специализацию I порядка. Так, очень многие виды являются растительноядными, или фитофагами, другие составляют группу хищников и паразитов, а остальные представлены всякого рода потребителями мертвой органики: сапрофаги питаются гниющими веществами, некрофаги — трупами животных, копрофаги — помётом, детритофаги — растительными остатками на поверхности почвы. При использования каких-либо отдельных тканей, органов или частей животного возникают дальнейшие подразделения рассмотренных специализаций: среди фитофагов появляются потребители листьев — филлофаги, плодов — карпофаги, древесины — ксилофаги, корней — ризофаги, а также галлообразователи; среди хищников и паразитов могут быть кровососы, эктопаразиты, эндопаразиты.
Иной формой является пищевая специализация II порядка — появление избирательности уже внутри вышеназванных основных источников пищи. Так, среди фитофагов, хищников и паразитов есть одноядные монофаги, ограниченноядные олигофаги и многоядные полифаги. Хищники и паразиты других насекомых (например, трихограммы) нередко обозначаются термином энтомофаги.
Само собой разумеется, что между всеми рассмотренными типами специализации есть переходы, но они вопреки ожиданию не столь многочисленны.
Места обитания
Насекомые обитают в подавляющем большинстве известных наземных биотопов, заняв такие негостеприимные экосистемы, как вершины гор, глубокие пещеры а также зарождающиеся экосистемы недавно образовавшихся островов вулканического происхождения. Известны и морские насекомые, относящиеся к особому семейству водомерок из отряда полужесткокрылых (кроме них, в прибрежных солёных водах изредка поселяются другие, типично пресноводные клопы).
Избирательность стаций (местообитаний) является весьма существенным и характерным свойством насекомых. Каждый вид имеет свой набор стаций — в одних случаях разнообразный, в других ограниченный экологическими рамками и иногда сведённый к единственному типу местообитания. Однако часто вид не ограничивается заселение только лишь одной стации: проявляется закономерное изменение видами своих местообитаний. Смена стаций может быть зональной, вертикальной, сезонной и годичной.
Зональная смена стаций свойственна многим трансзональным видам (то есть видам, ареал которых пересекает несколько природных зон): при продвижении к северу ареала избираются более сухие, хорошо прогреваемые открытые местообитания, в то время как при продвижении к югу заселяются более увлажненные и затененные стации, часто с густым растительным покровом. Особой формой зональной смены стаций является зональная смена ярусов — переход в сухих частях ареала наземных видов к подземному образу жизни. Вертикальная смена стаций аналогична зональной, но характерна для горных условий. Если горная система подпирается аридными ландшафтами — степями или пустынями, а низинная часть ареала отодвинута от горной к северу, тогда с повышением уровня виды переходят в более увлажнённые стации. При контакте низинной и горной частей ареала повышение вертикального уровня вызывает перемещение видов из более влажных стаций в менее увлажнённые и более открытые.
Сезонная и годичная смены стаций протекают уже во времени являются следствием вынужденных миграций вида в результате изменений микроклимата, погодных условий и состояния растительного покрова. Первая характерна для областей с жарким летом и заключается в переходе в более увлажнённые стации. Годичная смена стаций вызывается отклонением климатических показателей от средней нормы и приводит в сухие годы к перемещению ряда видов во влажные стации и, наоборот, во влажные годы — к перемещению в сухие стации.
Экологической основой смены стаций является строгое подчинение своему экологическому стандарту. В результате изменяется тип заселяемых стаций, что приводит в конечном итоге к изменению экологии вида, стимулирует внутривидовую дифференциацию и становится фактором эволюции. Нередки случаи, когда в разных зонах вид представлен разными подвидами или даже дифференцировался на два очень близких вида.
Географически увеличенная стация представляет собой ареал. Для многих насекомых известны сплошные ареалы, хотя в классе, как выясняется в последнее время, относительно высока доля видов-двойников, порой различающихся лишь по биохимическим особенностям. Также, как и ареалы других животных, ареал насекомых характеризуется принадлежностью к одной или нескольким зоогеографическим областям. Некоторые исследователи полагают, что можно создать единую подробную схему зоогеографического районирования, приемлемую для всех насекомых. Другие считают, что создание такой универсальной схемы невозможно, поскольку даже такая единая в таксономическом отношении группа, как насекомые, не представляет собой чего-то единого в отношении закономерностей географического распространения. Поэтому бессмысленно пытаться создать единую зоогеографическую карту, а следует разрабатывать карты для отдельных экологических групп насекомых. В частности, такая карта для реофильных амфибиотических насекомых (большинство подёнок) совершенно несопоставима с картой для растительноядных наземных насекомых и существенно отличается от карты лимнофильных амфибиотических насекомых, как, например, стрекозы.
Для общей характеристики расположения ареалов самых разных насекомых обычно используют широко признанные названия крупных разделов суши: Палеарктика, Неарктика, Голарктика, Амфипацифический сектор, Эфиопская область, Ориентальная область, Арктогея, Неотропики и Австралийская область.
Экстремальные условия обитания
Насекомые заселяют даже приполярные области: Антарктиду и Арктику (Гренландию, Новую Землю, остров Врангеля, Чукотский полуостров и другие). Эндемик Антарктики бескрылый комар-звонец Belgica antarctica из семейства Chironomidae (отряд двукрылые) к тому же крупнейшие истинно сухопутное (не покидающие поверхность земли) животное Антарктиды.[22] Гусеницы бабочки Gynaephora groenlandica (из семейства волнянок, Гренландия и Канада) способны во время зимовок переживать температуры до −70 °C.[23] На острове Гренландия есть несколько эндемичных видов насекомых, например жук Atheta groenlandica.[24]
Термофильный полуводный жук Zaitzevia thermae (из семейства речники, Elmidae) способен обитать в термальных источниках. В штате Монтана (США) он обнаружен в необычно тёплых водах — до 29 °C.[25]
Размножение
Насекомые раздельнополы. Органы размножения самки обычно представлены парными яичниками и тянущимися по бокам яйцеводами, которые сливаются в один непарный проток, впадающий во влагалище. У самок имеются семяприемники и придаточные половые железы. У самцов имеются парные семенники, от которых отходят семяпроводы, тянущиеся по бокам тела. В нижней части семяпроводы расширяются, образуя семенные пузырьки, предназначенные для хранения спермы. Семяпроводы объединяются в общий семяизвергательный канал, открывающийся на способном увеличиваться или выдвигаться совокупительном органе. Придаточные железы секретируют семенную жидкость.
Звуки в жизни насекомых
Роль звуков велика в жизни насекомых, хотя изучена слабо. Большой вклад в её изучение внесли биоакустики Е. К. Еськов, А. В. Попов, Р. Д. Жантиев[3].
Поскольку насекомые имеют малые размеры и не обладают единым развитым дыхательным трактом, в котором мог бы разместиться источник звука (гортань), они задействуют другие способы звукоизвлечения. Например, трение задней ноги (имеющей специальные бугорки) об надкрылья, как саранча, или надкрылий между собой (где левое обычно исполняет роль смычка), как у кузнечиков и сверчков. Звуки некоторых представителей последних могут разноситься на расстояние до полутора километров. Цикады используют специальную звуковую мембрану, приводимую в колебание мышцами. Получающийся металлический звук резонирует в специальных полостях внутри тела, достигая большой громкости. Южноамериканская цикада способна издавать звук, похожий на свист паровоза. Термиты при опасности в большом числе бьют головами о субстрат (материал термитника), создавая барабанный бой и оповещая таким образом других термитов[3].
Медоносные пчёлы издают звуки, заставляя вибрировать часть торакса путём частых мышечных сокращений. Звук усиливается крыловыми пластинами. В отличие от многих насекомых пчёлы способны издавать звуки разной высоты и тембров, что позволяет им передавать информацию посредством разных характеристик звука[3].
Органы чувств
Зрение
Зрение играет большую роль в жизни насекомых. Особенности зрения насекомых обусловлено фасеточным строением глаз. Насекомые близоруки — область точного зрения у них не превышает 1—2 см. Они отлично видят движение и цвет, в том числе ультрафиолет. Можно сказать, что позвоночные видят обычно формы и некие цвета, а насекомые — прежде всего движения и иные цвета.
Восприятие запаха
Восприятие запаха у насекомых составляет особое стереохимическое чувство. Обонятельные органы большинства позвоночных расположены внутри тела и неподвижны. У насекомых же чувствительные клетки, воспринимающие запах, расположены преимущественно на усиках (а также на ногах и некоторых других придатках тела), то есть на подвижных органах, на некотором расстоянии от оси тела. Каждый усик может двигаться, так что запах насекомые воспринимают вместе с пространством и направлением, для них это одно единое чувство.
Описать это можно таким образом, что млекопитающие (и люди в том числе) чувствуют изолированный запах, а насекомые ощущают «запаховое тело». Когда, например, цветок выделяет определённый аромат, то этот аромат «вставлен» в тело цветка определённым образом, в зависимости от того, какие органы цветка его выделяют; запаховое тело «вставлено» в тело цветка, как нога в башмак. Это «облако аромата» меняет форму и насыщенность в зависимости от наличия ветра и расстояния до предмета. Все это вместе и составляет «запах» для насекомого. Аналогом (хотя и весьма приблизительным) такого чувства у человека является чувство осязания: человек одновременно воспринимает фактуру предмета, а также расстояние от предмета до тела, направление, его температуру и влажность. Все это вместе входит в наше единое «чувство поверхности», к которой человек прикасается. Таким «общим чувством» для насекомых является чувство стереохимическое: объёмный запах.
Слух
Органы слуха расположены не на голове, поскольку она мала и насекомому нужно определять направление на источник звука. Для этого между приёмниками звука должно быть достаточное расстояние для создания бинаурального эффекта. У кузнечика, в частности, слуховые отверстия находятся на коленях передних ног, что позволяет расставить их пошире при необходимости[3].
Поведение
См. также: Общественные насекомыеРоль в природе
Насекомые составляют около 90 % всех животных на Земле[26], по разным оценкам в современной фауне от 2 до 10 млн видов насекомых, из них пока известно чуть более 1 млн. Активно участвуя в круговороте веществ, насекомые играют глобальную планетарную роль в природе.
Более 80 % растений опыляются насекомыми, и можно с уверенностью сказать, что цветок — результат совместной эволюции растений и насекомых. Приспособления цветковых растений для привлечения насекомых разнообразны: пыльца, нектар, эфирные масла, аромат, форма и окраска цветка. Приспособления насекомых: сосущий хоботок бабочек, грызуще-лижущий хоботок пчелиных; особые пыльцесобирательные аппараты — у пчёл и шмелей щеточка и корзинка на задних ногах, у пчёл-мегахил — брюшная щёточка, многочисленные волоски на ногах и теле.
Огромную роль насекомые играют в почвообразовании. Такое участие связано не только с разрыхлением почвы и обогащением её перегноем почвенными насекомыми и их личинками, но и с разложением растительных и животных остатков — опада растений, трупов и экскрементов животных, одновременно выполняется санитарная роль и круговорот веществ в природе.
Санитарную роль выполняют следующие виды насекомых:
- копрофаги — жуки-навозники, мухи-навозницы, коровницы;
- некрофаги — жуки-мертвоеды, могильщики, кожееды, мухи-мясоедки, падальницы;
- насекомые — разрушители отмерших растительных остатков: древесины, веток, листьев, хвои — жуки-сверлильщики, личинки усачей, златок, рогохвостов, комаров-долгоножек, муравьи-древоточцы, грибные комарики и т. д.;
- насекомые — санитары водоёмов питаются взвешенными или осевшими на дно гниющими органическими веществами (детритом) — личинки комаров-дергунов, или звонцов, поденок, ручейников, очищают воду и служат биоиндикатором её санитарного состояния.
Насекомые — важнейший элемент пищевых пирамид. Насекомыми питаются многие животные: рыбы, земноводные (у лягушки до 95 % рациона), пресмыкающиеся (ящерицы поедают до 10—20 насекомых в сутки), птицы, млекопитающие (чисто насекомоядными являются землеройки, кроты, летучие мыши, муравьеды, броненосцы, как дополнение к основной пище насекомые служат многим другим видам). Среди беспозвоночных, употребляющих в пищу насекомых, скорпионы, фаланги, сольпуги, сенокосцы, пауки, сколопендры, костянки и огромная армия насекомых-энтомофагов (хищников и паразитов).
Взаимодействие с человеком
Ряд насекомых переносят опасные болезни, некоторые являются паразитами или просто досаждают человеку (см. Энтомозы). Но есть и насекомые, приносящие человеку непосредственную пользу: к примеру, благодаря пчёлам возможно производство мёда.
Существуют певчие насекомые[3]:
В Японии можно услышать поющих насекомых, и в Токио по крайней мере сорок торговцев занимаются их продажей. Во Всех японских городах можно видеть весной и летом на верандах домов маленькие бамбуковые клетки, откуда слышится странный свист, монотонное жужжание, подчас бесконечно долгие трели, металлические вибрации, наполняющие сумерки изящной оригинальной музыкой. Самым любимым поющим насекомым является похожее на жука с удлинённым и плоским телом — судзумуси (японский сверчок), что по-японски значит: насекомое-колокол, так как звук его голоса напоминает маленький серебряный колокольчик.
Наука о насекомых
Изучением насекомых занимается энтомология и многочисленные её подразделы: диптерология (наука о двукрылых насекомых), колеоптерология (о жуках), лепидоптерология (о бабочках), мирмекология (о муравьях), ортоптерология (о прямокрылых) и многие другие. Научная история энтомологии берёт своё начало примерно с XVI века. Среди известных энтомологов такие крупнейшие биологи, как Чарльз Дарвин и Карл Фриш (Нобелевский лауреат 1973 года), писатель Владимир Набоков и дважды лауреат Пулитцеровской премии профессор Эдвард Уилсон. Лаборатории и кафедры энтомологии существуют во многих крупнейших институтах (например, в ЗИН РАН) и университетах (с 1925 года — в МГУ, с 1919 года — в СПбГУ, с 1920 года — в МСХА). К старейшим энтомологическим обществам относятся Энтомологическое общество Франции (1832), Королевское энтомологическое общество Лондона (1833), Русское энтомологическое общество (1859) и другие. Свои научные труды они публикуют в многочисленных энтомологических журналах. С 1910 года регулярно проводятся международные энтомологические конгрессы (23-й прошёл в 2008 году в Дурбане, ЮАР).
Классификация
Происхождение
См. также: ПалеоэнтомологияУстановление происхождения класса насекомых сталкивается с определёнными трудностями. Ключевая проблема состоит в отсутствии окаменелостей, по которым можно было бы установить филогенетические отношения насекомых[27].
Традиционно на основе морфологических сравнений (в особенности трахеальных дыхательных систем) ближайшими родственниками насекомых считали многоножек. В соответствии с этой точкой зрения обе группы объединяли в таксон «неполноусые» (Atelocerata).
Однако построение филогений на основе последовательностей генов и недавние морфологические исследования свидетельствуют, что насекомые ближе к ракообразным, чем к многоножкам. Сторонники этой филогенетической гипотезы объединили ракообразных и насекомых в единый таксон Pancrustacea. Данная гипотеза также небесспорна. Ракообразные известны из отложений раннего кембрия (около 511 млн лет), в то время как насекомые появляются в палеонтологической летописи только в раннем девоне (около 410 млн лет). Таким образом, гипотеза о происхождении ракообразных и насекомых от единого общего предка порождает временной промежуток около 100 млн лет, в течение которого насекомые должны были существовать. Отсутствие останков насекомых в палеонтологической летописи в течение указанного периода обычно объясняется тем, что они не сохранились до наших дней[27].
Новейшие морфологические сравнения и филогенетические реконструкции на основе геномных последовательностей указывают, что насекомые являются потомками ракообразных, а не сестринским таксоном. Это заключение хорошо согласуется с палеонтологическими данными. Однако морфологические и молекулярные данные не согласуются при определении ближайших родственников насекомых среди ракообразных: морфологические данные указывают на связь насекомых с высшими ракообразными, а молекулярные — с жаброногими[27].
В соответствии с последней гипотезой эволюционная ветвь насекомых отделилась от ракообразных в позднем Силуре — раннем Девоне. Эта оценка согласуется и с палеонтологическими данными, и с оценкой на основе молекулярных часов[27].
Насекомые в культуре

Этот раздел не завершён. Вы поможете проекту, исправив и дополнив его.- Левша подковал танцующего робота-блоху в повести Лескова «Сказ о тульском косом Левше и о стальной блохе».
- Пчёлы и муравьи — одни из главных героев в некоторых детских мультфильмах и мульсериалах («Муравей Антц», «Приключения Флика», «Приключения пчёлки Майи» и др.).
- Пришельцы в фантастике часто имеют внешний вид и социальную организацию насекомых («Звёздный десант» и его экранизации, «Лексс» и др.).
См. также
Примечания
- Литература
- ↑ 1 2 3 Бей-Биенко, 1980, с. 60
- ↑ 1 2 Бей-Биенко, 1980, с. 62
- ↑ Тыщенко, 1986, с. 113—116
- ↑ 1 2 3 4 5 Бей-Биенко, 1980, с. 61
- ↑ 1 2 Тыщенко, 1986, с. 98
- ↑ Тыщенко, 1986, с. 112
- Другие источники
- ↑ Горностаев Г. Н. 1998. Насекомые. Энциклопедия природы России. — М.: ABF. 560 с.
- ↑ 1 2 Chapman, A. D. Numbers of living species in Australia and the World. — Canberra: Australian Biological Resources Study, 2006. — P. 60. — ISBN 978-0-642-56850-2
- ↑ 1 2 3 4 5 6 Морозов В. П. Занимательная биоакустика. Изд. 2-е, доп., перераб. — М.: Знание, 1987. — 208 с. + 32 с. вкл. — С. 36-42
- ↑ Фасмер М. Этимологический словарь русского языка. — Прогресс. — М., 1964–1973. — Т. 3. — С. 47.
- ↑ Черных П. Я. Историко-этимологический словарь современного русского языка. — М.: Русский язык, 1993. — Т. 1. — С. 560.
- ↑ May, R.M. (2000). The Dimensions of Life on Earth. pp. 30-45 in Raven, P.H. and Williams, T. (eds) Nature and Human Society: The Quest for a Sustainable World. Washington, DC: National Academy Press
- ↑ Myers, P. (2001a). Insecta, Animal Diversity Web. http://animaldiversity.ummz.umich.edu/site/accounts/information/Insecta.html
- ↑ Nielsen, E.S. and Mound, L.A. (2000). Global Diversity of Insects: The Problems of Estimating Numbers. pp. 213—222 in Raven, P.H. and Williams, T. (eds). Nature and Human Society: The Quest for a Sustainable World. Washington, DC: National Academy Press
- ↑ Raven, P.H. and Yeates, D.K. (2007). Australian biodiversity: threats for the present, opportunities for the future. Australian Journal of Entomology 46: 177—187
- ↑ Hammond, P.M. (1995). The current magnitude of biodiversity pp. 113—128 in Heywood, V.H. (ed.). Global biodiversity assessment. Cambridge, UK: Cambridge University Press
- ↑ Grove S. J., Stork N. E. An inordinate fondness for beetles. Invertebrate Taxonomy 14. — 2000. — P. 733—739.
- ↑ Bragg P. E. (2008): In Hennemann & Conle. Revision of Oriental Phasmatodea: The tribe Pharnaciini Günther, 1953, including the description of the world’s longest insect, and a survey of the family Phasmatidae Gray, 1835 with keys to the subfamilies and tribes (Phasmatodea: «Anareolatae»: Phasmatidae). Zootaxa, vol. 1906, pp. 1—316
- ↑ World’s longest insect revealed. Лондонский музей естествознания (16 октября 2008). Архивировано из первоисточника 20 марта 2012. Проверено 8 декабря 2009.
- ↑ Brock, P.D. 1999. The amazing world of stick and leaf-insects. Cravitz Printing Co., Essex, England
- ↑ Vanessa Hequet. Longicornes de Guyane. — Cayenne, 1996.
- ↑ Mares J. & V. Lapacek. Nejkrasnejsi brouci tropu. — Praha, 1980.
- ↑ Yukio Yaznda & Shuji Okejima. Käfer der Welt. 700 col illus. — 1990. — P. 126.
- ↑ 1 2 Каабак Л. В., Сочивко А. В. Бабочки мира. — М.: Аванта+, 2003. ISBN 5-94623-008-5
- ↑ Hugo Kons, Jr. Chapter 32 — Largest Lepidopteran Wing Span (англ.). Book of Insect Records. University of Florida (17 May 1998). Архивировано из первоисточника 18 августа 2011.
- ↑ Вейбрен Ландман. Бабочки. Иллюстрированная энциклопедия. — М.: Лабиринт Пресс, 2002. — 272 с. — (Иллюстрированная энциклопедия). — ISBN 5-9287-0274-4
- ↑ Лобанов А. Л. Жуки-рекордсмены (рус.). ЗИН РАН (2000). Архивировано из первоисточника 20 мая 2012. Проверено 12 июня 2011.
- ↑ Luke Sandro, Juanita Constible. Antarctic Bestiary — Terrestrial Animals. Laboratory for Ecophysiological Cryobiology, Miami University. Архивировано из первоисточника 9 июля 2012. Проверено 26 февраля 2012.
- ↑ Morewood, W. Dean & Richard A. Ring. Revision of the life history of the High Arctic moth Gynaephora groenlandica (Wocke) (Lepidoptera: Lymantriidae) // Can. J. Zool.. — 1998. — Vol. 76. — № 7. — P. 1371–1381.
- ↑ Robert Bergersen. Is Greenland a Zoogeographical Unit of Its Own? // Journal of Biogeography. — январь 1995. — Vol. 22. — № 1. — P. 1-6.
- ↑ Чрезвычайно узкий эндемик из термального источника в США (Zaitzevia thermae) носит имя Ф. А. Зайцева (Проверено 3 августа 2012)
- ↑ Erwin, Terry L. (1982). «Tropical forests: their richness in Coleoptera and other arthropod species». Coleopt. Bull. 36: 74-75.
- ↑ 1 2 3 4 Glenner H. et al. The Origin of Insects. Science, 2006, V. 314, p. 1883—1884
Литература
- Бей-Биенко Г. Я. Общая энтомология: Учебник для университетов и сельхозвузов. — 3-е изд., доп. — М.: Высшая школа, 1980. — 416 с.
- Жизнь животных. Энциклопедия в 6 т. Т. 3. (том посвящён сухопутным членистоногим). Общая редакция члена-корреспондента АН СССР профессора Л. А. Зенкевича. — М.: Просвещение, 1969. — 576 с.
- Тыщенко В. П. Физиология насекомых: Учеб. пособие для студентов ун-тов, обучающихся по спец. «Биология». — М.: Высшая школа, 1986. — 303 с.
Ссылки
- Информационная система ZINsecta — новая классификация насекомых, подготовленная Лабораторией систематики насекомых на сайте Зоологического института РАН
Современные Членистоногие, подтипы, надклассы и классы Царство Животные • Подцарство Eumetazoa • Раздел Bilateria • Подраздел Protostomia • Надтип Ecdysozoa
Хелицеровые (Chelicerata) Паукообразные (Arachnida) • Меростомовые (Merostomata) • Морские пауки (Pantopoda)
Многоножки (Myriapoda) Губоногие (Chilopoda) • Двупарноногие (Diplopoda) • Пауроподы (Pauropoda) • Симфилы (Symphyla)
Шестиногие (Hexapoda) Насекомые (Insecta) • Скрыточелюстные (Entognatha)
Ракообразные (Crustacea) Жаброногие (Branchiopoda) • Ремипедии (Remipedia) • Цефалокариды (Cephalocarida) • Челюстеногие (Maxillopoda) • Ракушковые (Ostracoda) • Высшие раки (Malacostraca)
Категории:- Животные по алфавиту
- Насекомые
- Классы животных







{kind=link}
Wikimedia Foundation. 2010.