- Рыбы
-
Рыбы 
Макропод (Macropodus opercularis)Научная классификация промежуточные рангиДомен: Эукариоты Царство: Животные Подцарство: Эуметазои Без ранга: Двусторонне-симметричные Без ранга: Вторичноротые Тип: Хордовые Подтип: Позвоночные Инфратип: Челюстноротые Клада: Рыбы Международное научное название Pisces
Классы - Костные рыбы (Osteichthyes)
- Хрящевые рыбы (Chondrichthyes)
- †Акантоды (Acanthodii)
- †Плакодермы (Placodermi)

Изображения
на ВикискладеEOL 4656086 Рыбы (лат. Pisces) — надкласс (в соответствии с современными принципами кладистики — парафилетическая группа) водных позвоночных животных. Обширная группа челюстноротых, для которых характерно жаберное дыхание на всех этапах постэмбрионального развития организма. Рыбы распространены как в солёных, так и в пресных водах, от глубоких океанических впадин до горных ручьев. Рыбы играют важную роль в большинстве водных экосистем как составляющая пищевых цепей. Они имеют большое экономическое значение для человека из-за употребления их в пищу.
Размеры современных рыб варьируют от 7,9 мм (Paedocypris progenetica[1]) до 13,7 м (китовая акула).
В мире известно, по разным данным, от 25 000 до 31 000 видов рыб. В России обитает около 3000 видов[2], в том числе в пресных водах встречается более 280 видов[3].
Изучению рыб посвящён раздел зоологии — ихтиология. Ввиду важности изучения и большого экономического значения рыб в некоторых ВУЗах (например, на биологическом факультете МГУ) есть, наряду с кафедрами зоологии позвоночных, отдельные кафедры ихтиологии.
Содержание
Происхождение и эволюция рыб
 Окаменелость панцирной рыбы Bothriolepis panderi или астеролеписа
Окаменелость панцирной рыбы Bothriolepis panderi или астеролеписа
Считалось, что наиболее древние известные бесчелюстные животные, напоминающие рыб, известны из раннего ордовика (около 450—470 миллионов лет назад). Однако в 1999 году в китайской провинции Юньнань были найдены окаменелости рыбообразного существа Haikouichthys из группы бесчелюстных возрастом около 530 миллионов лет (ранний кембрий). Возможно, подобные формы были предками всех позвоночных.
 Ископаемые останки примитивной рыбы Osteolepis macrolepidotys из середины девонского периода (Великобритания)
Ископаемые останки примитивной рыбы Osteolepis macrolepidotys из середины девонского периода (Великобритания)С развитием челюстей из одной из жаберных дуг появились первые рыбы. Кроме челюстей, рыбы имеют парные плавники, внутреннее ухо с тремя полукруглыми каналами и жаберные дуги. Несмотря на появление первых челюстноротых рыб еще в ордовике, они занимали подчиненное положение вплоть до девона. Таким образом, рыбы и бесчелюстные существовали больше 100 миллионов лет в условиях преобладания бесчелюстных, в отличие от настоящего времени. Хрящевые рыбы появились на рубеже силура и девона, около 420 миллионов лет назад, и достигли расцвета в карбоне. Костные рыбы водятся в мировых океанах как минимум с девона; не исключено, что они существовали уже в силуре.

Более половины всех живущих ныне видов позвоночных, а именно, согласно базе данных FishBase, около 31 тысячи видов, относятся к рыбам. Число признанных видов продолжает меняться вследствие открытий новых видов, а также таксономических ревизий отдельных групп рыб. Ныне живущие рыбы представлены двумя классами: хрящевые рыбы (Chondrichthyes) и костные рыбы (Osteichthyes).
Современных хрящевых рыб делят на две большие группы — Holocephali (или цельноголовые), и Elasmobranchii, куда входят акулы и скаты. Ныне существует 900—1000 видов хрящевых рыб. В свою очередь, костные рыбы делятся на два подкласса: лопастепёрые и лучепёрые.
Анатомия и физиология
Внешние покровы
За редкими исключениями, внешние покровы рыб представлены кожей с чешуей (у некоторых рыб чешуя отсутствует). Как и у всех других позвоночных, кожа рыб делится на дерму и эпидермис (верхний слой кожи эктодермального происхождения, состоящий из эпителиальной ткани). Эпидермис у рыб неороговевающий. Железы в эпидермисе секретуют мускусоподобный секрет, который защищает внешние покровы животного.
В формировании чешуи основную роль играет внутренний слой кожи — эпидермис. Хрящевые рыбы имеют плакоидную чешую, которая гомологична зубам всех позвоночных; перемещаясь в ходе эволюции на челюсти, плакоидные чешуи, собственно, и превращаются в зубы у акул и скатов. Плакоидная чешуя состоит из дентина, который формирует основу чешуй, а сверху покрыта эмалью. По химическому составу дентин и эмаль акул сходны с дентином и эмалью зубов человека. Утраченные плакоидные чешуи не возобновляются, но при росте рыбы их количество увеличивается. Плавниковые шипы некоторых хрящевых рыб (например, у черноморского катрана) тоже являются преобразованными плакоидными чешуями.
Костистым рыбам присуще несколько разных типов чешуи.
 Чешуя костных рыб
Чешуя костных рыбГаноидная чешуя имеется у наиболее примитивных из лучепёрых рыб, например, осетровых. Она сформирована костяными пластинками, которые сверху покрыты пластом похожего на дентин вещества — ганоина; часто такая чешуя покрывает тело рыбы сплошным защитным панцирем, как у представителей семейств Polypteridae и Lepisosteidae.
Для ископаемых кистеперых и Двоякодышащих рыб, а также для современных кистеперых рыб характерная космоидная чешуя, внешняя поверхность которой образована пластом космина (откуда происходит название), а сверх него — дентина. Космин подстилается пластом губчатой кости. У современных видов кистеперых и двоякодышащих рыб внешний дентиновый и внутренний губчатый пласты постепенно редуцируются — у современных видов рода Латимерия на поверхности чешуи сохранились лишь одиночные бугорки дентина.
Чешуя настоящих костистых рыб называется эласмоидной и делится на две разновидности: ктеноидную (зубчатую) и циклоидную (округлую) на основании формы внешней кромки. В отличие от большинства подвидов плакоидных и ганоидних чешуй, циклоидные и ктеноидные расположены так, что передние накладываются на задние, а сами чешуйки анатомически являются тонкими костными пластинками; в последнее время было установлено, что гребенчатая поверхность ктеноидной чешуи улучшает гидродинамические свойства рыб.
Окраска рыб может варьировать в очень широких пределах: от однотонной практически всех возможных цветов до маскировочной «камуфляжной» или, наоборот, подчеркнуто яркой «предупреждающей».
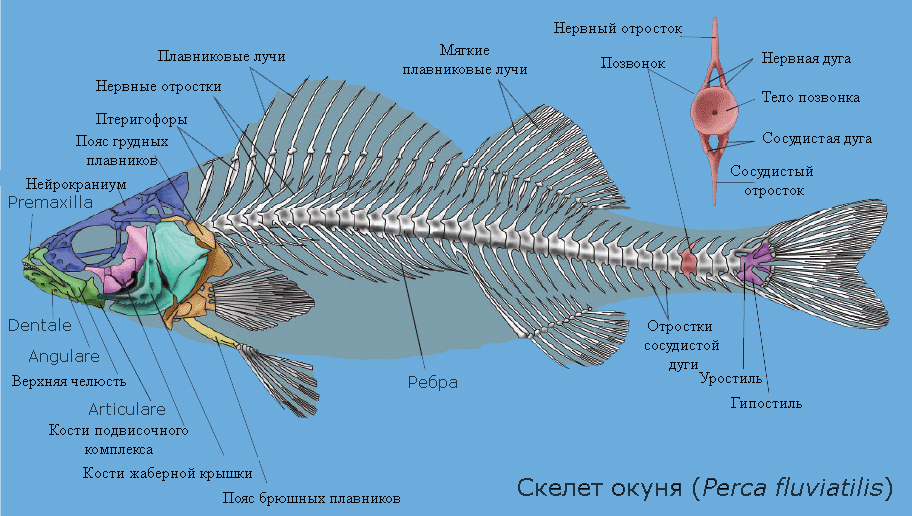
 Скелет рыб, на примере окуня
Скелет рыб, на примере окуняСкелет и мышечная система
Опорно-двигательная система рыб — система органов и тканей рыб, которая позволяет им осуществлять движения и корректировать свое положение в окружающей среде. Благодаря эволюционным видоизменениям части опорно-двигательной системы приспособлены для выполнения также и других специализированных функций.
В отличие от наземных позвоночных, имеющих которых череп с большим количеством сращенных костей, череп рыб содержит более чем 40 костных элементов, которые могут двигаться независимо. Это позволяет осуществлять вытягивание челюстей, раздвигание челюстей в стороны, опускать жаберный аппарат и дно ротовой полости.
Подвижные элементы прикрепляются к более жестко сочлененному нейрокраниуму, который окружает головной мозг. Нейрокраниум костных рыб в эволюции возникает из хрящевого черепа хрящевых рыб, к которому прирастают кожные костные пластинки.
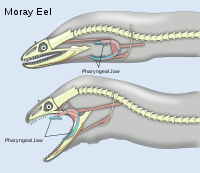 Мурены обладают двумя парами челюстей. Вторая пара, называемая глоточной, или фарингеальной, находится позади черепа. Хотя внутренние челюсти не выходят за пределы внешних, они помогают более крепко ухватить добычу
Мурены обладают двумя парами челюстей. Вторая пара, называемая глоточной, или фарингеальной, находится позади черепа. Хотя внутренние челюсти не выходят за пределы внешних, они помогают более крепко ухватить добычуЧелюсти в классах костных и хрящевых рыб эволюционно образовались из третьей пары жаберных дуг (о чем свидетельствуют рудименты первых двух пар дуг у акул — так называемые губные хрящи). У костистых рыб челюсти несут основные группы зубов на переднечелюстной (premaxilla) и верхнечелюстной костях (maxilla) (верхняя челюсть), на dentale и articulare (нижняя челюсть). Несколько специализированных групп костей формируют дно ротовой полости и объединяют челюсти с другими элементами черепа. Наиболее рострально (впереди) расположенная гиоидная дуга, которая играет важную роль при изменении объема ротовой полости. За ней идут жаберные дуги, которые несут жаберные дыхательные структуры, и наиболее каудально расположены так называемые глоточные челюсти, которые также могут нести зубы.
Во время питания мышцы, которые опускают комплекс нижней челюсти, смещают этот комплекс таким образом, что челюсти выдвигаются вперед. При этом в ротовой полости создаётся всасывающая сила за счёт опускания дна рта. Жаберные крышки при этом закрывают жабры. Такая комбинация движений приводит к всасыванию воды и затягиванию пищи в рот.
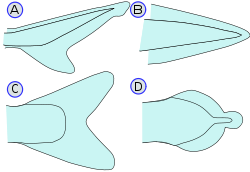 Виды хвостовых плавников рыб. (A) — Гетероцеркальный, (B) — Протоцеркальный, (C) — Гомоцеркальный, (D) — Дифицеркальный
Виды хвостовых плавников рыб. (A) — Гетероцеркальный, (B) — Протоцеркальный, (C) — Гомоцеркальный, (D) — ДифицеркальныйДвижущая сила при плавании рыб создаётся плавниками: парными (грудные и брюшные) и непарными (спинной, анальный, хвостовой). При этом у костных рыб плавники состоят из костных (у некоторых примитивных — из хрящевых) лучей, объединенных перепонкой. Присоединенные к основным лучам мышцы могут разворачивать или свертывать плавник, изменять его ориентацию или генерировать волнообразные движения. Хвостовой плавник, который у большинства рыб является основным движителем, поддерживается набором специальных сплюснутых костей (уростиль и др.) и ассоциированных с ними мышц в дополнение к боковым мышцам туловища. По соотношению размеров верхней и нижней лопасти хвостовой плавник может быть гомоцеркальным (когда обе лопасти имеют равную величину; это характерное для большинства лучевых рыб) или гетероцеркальным (когда одна лопасть, обычно верхняя, больше другой; характерное для акул и скатов, а также осетровых; у таких представителей костных рыб как меченосцы хвостовой плавник гетероцеркальный с большей нижней лопастью).
Позвоночник рыб состоит из отдельных, не срощенных позвонков. Позвонки рыб амфицельные (то есть их обе торцевые поверхности вогнутые), между позвонками находятся хрящевая прослойка; невральные дуги, расположенные сверху над телами позвонков, формируют позвоночный канал, защищающий спинной мозг. Позвоночник делится на два отдела - туловищный и хвостовой. От позвонков, которые находятся в туловище, в стороны отходят реберные отростки, к которым прикрепляются ребра. В хвостовом отделе позвоночника боковых отростков на позвонках нет, зато кроме невральной дуги имеется сосудистая (гемальная) дуга, которая прикрепляется к позвонку снизу и защищает проходящий в ней большой кровеносный сосуд — брюшную аорту. От невральных и гемальых дуг вертикально вверх и вниз отходят заостренные остистые отростки.
По правую сторону и левую сторону от позвоночника отходит мембрана из соединительной ткани, которая называется горизонтальной септой (перегородкой) и разделяет мышцы тела рыбы на дорсальную (верхнюю) и вентральную (нижнюю) части, которые называются миомерами.
Плавание рыб осуществляется благодаря сокращению мышц, которые объединены сухожилиями с позвоночником. Миомеры в теле рыбы имеют форму конусов, вложенных один в один и разделенных перегородками соединительной ткани (миосептами). Сокращение миомеров через сухожилие передается на позвоночник, побуждая его к волнообразному движению (ундуляции) по всей длине тела или лишь в хвостовом отделе.
В целом мускулатура рыб представлена двумя типами мышц. «Медленные» мышцы используются при спокойном плавании. Они содержат много миоглобина, который обуславливает их красный цвет. Метаболизм в них в основном аэробный, то есть в них происхожит полное окисление питательных веществ. Такие красные мышцы могут долго не утомляться, и потому используются при длинном монотонном плавании. В отличие от красных, «быстрые» белые мышцы с преимущественно гликолитичным метаболизмом способны к быстрому, но кратковременному сокращению. Они используются при быстрых внезапных рывках; при этом они могут давать большую, чем красные мышцы, мощность, но быстро утомляются.
Также у многих рыб мышцы могут выполнять и некоторые другие функции, кроме движения. У некоторых видов они участвуют в терморегуляции (термогенезе). У тунцов (Scombridae) благодаря активности мускулатуры температура мозга поддерживается на более высоком уровне, чем температура других частей тела, когда тунцы охотятся на кальмаров в глубоких холодных водах.
Электрические токи, которые генерируются при сокращении мышц, используются слонорылом как коммуникационный сигнал; у электрических скатов электрические импульсы, генерированные видоизмененными мышцами, используются для поражения других животных. Модификация мышечных клеток для выполнения функции электрической батареи эволюционно происходила независимо и неоднократно в разных таксонах: глазных мышц у рыб-звездочетов (Uranoscopidae), жевательной мускулатуры (электрические скаты) или осевой мускулатуры (электрические угри).
Нервная система и органы чувств
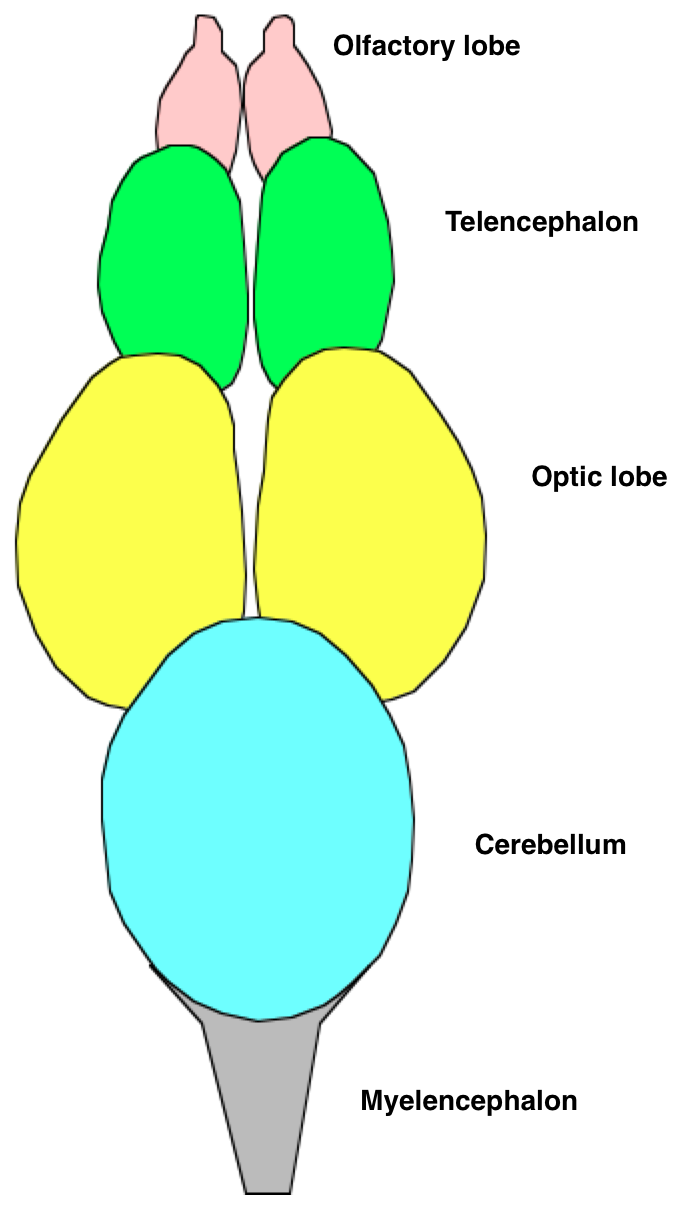
 Мозг рыбы
Мозг рыбыГоловной мозг рыб принято делить на три большие части: передний, средний и задний мозг. Передний мозг состоит из конечного мозга и промежуточного мозга. На ростральном (переднем) конце конечного мозга расположены обонятельные луковицы, которые получают сигналы от обонятельных рецепторов. Обонятельные луковицы обычно увеличены у рыб, которые активно используют нюх, например, у акул. В крыше среднего мозга находятся оптические доли. Задний отдел подразделяется на собственно задний мозг (к нему относится мост и мозжечок) и продолговатый мозг. Таким образом, у рыб, как и у других позвоночных, имеется пять отделов головного мозга (спереди назад): конечный, промежуточный, средний, задний и продолговатый.
Спинной мозг проходит внутри нервных дуг позвонков по всей длине позвоночника рыбы. Аналогично миомерам и позвоночнику, в строении спинного мозга наблюдается сегментация. В каждом сегменте тела сенсорные нейроны входят в спинной мозг через дорсальные корешки, а двигательные нейроны выходят из него через вентральные.
Глаза рыб по своему строению очень схожи с глазами других позвоночных. Важное отличие глаза рыб от глаза млекопитающих заключается в том, что для аккомодации рыбы не изменяют кривизну хрусталика, а приближают его к сетчатке или отдаляют от нее. Структура сетчатки у рыб варьирует в зависимости от места их обитания: у глубоководных видов глаза приспособлены для восприятия света преимущественно красной части спектра, а рыбы, которые живут на мелководье, воспринимают более широкий спектр.
Обоняние и вкус позволяют рыбам ориентироваться в химическом составе окружающей среды. Способность рыб к ощущению химических сигналов хорошо иллюстрируют лососи, которые, идя на нерест из моря к речным системам, определяют по запаху воды именно тот ручей или реку, в котором когда-то сами вышли из икры. Обонятельные рецепторы рыб расположены в ноздрях. Они, в отличие от ноздрей других позвоночных, не соединяются с носоглоткой (хоаны есть только у двоякодышащих рыб). Вкусовые рецепторы у многих рыб есть не только в ротовой полости, но и на жаберных структурах, усиках и даже плавниках и просто на поверхности тела.
Механорецепторы рыб содержатся во внутреннем ухе (парные органы слуха и равновесия), а также органах боковой линии. Внутреннее ухо пластинчатожаберных (акул и скатов) и костистых рыб состоит из трех полукружных каналов, расположенных в трех взаимно перпендикулярных плоскостях, и трех камер, каждая из которых содержит отолиты. Некоторые виды рыб (например, серебряный карась и разные виды сомов) имеют комплекс косточек, которые называются Вебберов аппарат и соединяют ухо с плавательным пузырем. Благодаря этой адаптации внешние вибрации усиливаются плавательным пузырем, как резонатором. Отолит в третьей камере обеспечивает рыбе ориентацию в пространстве.
 Боковая линия у акулы
Боковая линия у акулыПеремещение воды по поверхности рыбы ощущается структурами, которые называются нейромасты. Эти органы могут быть рассеяны поодиночке, или собраны под чешуёй в совокупности каналов, которые называются боковой линией. Нейромасты включают полушарие гелевой консистенции (капулу) и сенсорные волосовидные клетки, а также синапсы нервных волокон, которые находятся на волосовидных клетках. Перемещение воды служит причиной отгиба волосовидных клеток, который создаёт нервные импульсы. Эти импульсы позволяют составить довольно подробную картину окружающей среды: некоторые виды рыб, лишенные глаз, целиком ориентируются и перемещаются, полагаясь только на органы боковой линии.
Ощущение электрического поля — электрорецепция — присуще многим видам рыб — не только тем, которые могут сами генерировать электрические разряды. Электрические сигналы улавливаются с помощью специальных ямок на поверхности тела. Эти ямки заполнены гелеподобным веществом, которое проводит электрический ток и содержит в себе электрорецепторные клетки, которые образуют синапсы с нейронами.
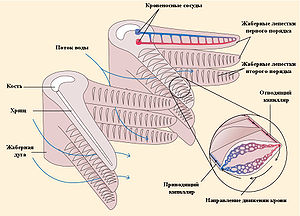 Схема работы жаберного аппарата рыб
Схема работы жаберного аппарата рыб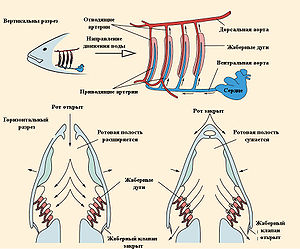 Схема газообмена у рыб
Схема газообмена у рыбКровеносная система и газообмен
У рыб один круг кровообращения и двухкамерное сердце. Кровеносная система замкнутая, транспортирует кровь от сердца через жабры и ткани тела. В отличие от сердца других позвоночных, сердце рыб не приспособлено для разделения (даже частичного) обогащенной кислородом крови (артериальной) от необогащённой (венозной). Структурно сердце рыб представляет собой последовательную серию из камер, заполненных венозной кровью: венозный синус, предсердие, желудочек и артериальный конус. Камеры сердца разделены клапанами, которые позволяют крови при сокращении сердца двигаться только в одном направлении (от венозного синуса к артериальному конусу), но не наоборот.
Основным органом газообмена рыб являются жабры, которые расположены по сторонам ротовой полости. У костистых рыб они закрыты жаберной крышкой, у других классов — свободно приоткрываются наружу. Во время вентиляции жабр вода попадает в ротовую полость через рот, а потом проходит между жаберными дугами и выходит наружу из-под жаберных крышек. Анатомически жабры состоят из полупроницаемых мембран и кровеносных сосудов, которые расположены на костных жаберных дугах. Специфической структурой, приспособленной для газообмена, являются жаберные лепестки, где под тонким эпителием находятся сильно разветвленные капилляры.
В дополнение к жабрам рыбы могут использовать и другие способы газообмена. На стадии личинки значительная часть газообмена осуществляется через кожу; несколько видов рыб имеют «легкие», где сохраняется увлажненный воздух (амия); некоторые виды могут дышать воздухом непосредственно (гурами).
Пищеварительная система и питание
Пищу захватывают и удерживают недифференцированными зубами, находящимися во рту. Изо рта, через глотку, а после - и пищевод, пища попадает в желудок, там подвергается обработке ферментами, содержащихся в желудочном соке, вырабатываемым желудком, после желудка пища попадает в тонкую кишку с протоками печени и поджелудочной железы, а оттуда через анальное отверстие или клоаку, у некоторых видов, выводятся наружу непереработанные остатки пищи.
Рыбам присущ широкий спектр пищевых объектов и способов питания. В целом все рыбы могут быть разделены на травоядных, хищников, детритофагов и всеядных. Растительноядные рыбы могут питаться как макро-, так и микроводорослями, а также водными цветочными растениями. Некоторые из таких рыб приспособлены к питанию планктоном, фильтруя его специализированными жаберными тычинками на жаберных дугах: например, разные виды толстолобиков (Hypophthalmichthys molitrix, Hypophthalmichthys nobilis) питаются исключительно за счет этого ресурса и являются строго определенными рыбами-фильтраторами микроскопических водорослей, которые живут в толще воды. Морские рыбы из семейства Pomacentridae питаются бентосными макроводорослями, причем каждая рыба имеет определенный участок, где она вырывает все водоросли, оставляя для роста только те виды, которыми питается — при этом не вырывая их во время питания, а лишь частично объедая.
Хищные рыбы используют в качестве пищевого ресурса широчайший спектр объектов. Планктонные фильтраторы (такие как Китовая акула) отфильтровывают зоопланктон из толщи воды; кроме того, рыбы могут питаться ракообразными, моллюсками, плоскими, круглыми и кольчатыми червями, а также другими рыбами. Наиболее специализированными из хищных рыб являются те, которые питаются эктопаразитами других рыб (Labridae, Chaetodontidae и прочие), которые выбирают паразитов и отмершие кусочки кожи с поверхности тела рыб-«клиентов», которые специально посещают места обитания «чистильщиков».
Много видов рыб могут изменять тип питания на протяжении жизни: питающиеся планктоном в молодом возрасте, могут переходить на питание рыбами или крупными беспозвоночными во взрослой фазе развития.
Выделительная система и осморегуляция
Следствием проживания в водной среде является ряд проблем с осморегуляцией, с которыми сталкиваются как пресноводные, так и морские рыбы. Осмотическое давление крови рыб может быть как ниже (у солоноводных рыб), так и выше (у пресноводных), чем осмотическое давление внешней среды. Хрящевые рыбы — изоосмотические, но при этом в их организме концентрация солей намного ниже, чем в окружающей среде. Выравнивание осмотического давления при этом достигается благодаря повышенному содержанию мочевины и триметиламиноксида (ТМАО) в крови. Поддержание низкой концентрации солей в организме хрящевых рыб осуществляется благодаря выделению солей почками, а также специализированной ректальной железой, которая соединяется с пищеварительным трактом. Ректальная железа концентрирует и выводит как ионы натрия, так и хлорид-ионы из крови и тканей организма.
Костистые рыбы не является изоосмотическими, поэтому в ходе эволюции выработали механизмы, которые позволяют осуществлять вывод или задержку ионов. Морские костистые рыбы с низкой (относительно окружающей среды) концентрацией ионов в организме постоянно теряют воду, которая под действием осмотического давления выходит из их тканей наружу. Эти потери компенсируются за счет питья и фильтрации соленой воды. Катионы натрия и хлорид-ионы выводятся из крови через жаберные мембраны, в то время как катионы магния и сульфатные анионы выводятся почками. Пресноводные рыбы сталкиваются с противоположной проблемой, благодаря наличию в организме солей в концентрации высшей, чем в окружающей среде. Осмотическое давление в их организме выравнивается благодаря захвату ионов из водной среды через жаберные мембраны, а также благодаря выделению большого количества мочевины.
Размножение
Типы размножения
Двуполое размножение
Двуполое размножение является наиболее обычной и широко распространенной его формой. При этом способе репродукции, самки и самцы внутри вида являются четко отделенными. При этом некоторые виды могут демонстрировать очень ярко выраженные вторичные половые признаки, или половой диморфизм. Эти характеристики вторичных половых признаков обычно проявляются только одним полом (в большинстве случаев — у самцов), не относятся к половому созреванию, могут интенсифицироваться на протяжении брачного сезона, не оказывают содействие индивидуальному выживанию. Вторичные половые признаки могут проявляться в виде различий в размерах тела, частей тела (например, удлиненные плавники), строения тела (например, выросты на голове), расположении зубов, окраске, а также встречаются в виде отличий между акустическими, химическими, электрическими и характеристиками самцов и самок. Двуполый способ размножения может включать в себя моногамию, полигамию и промискуитет.
Гермафродитизм
Второй способ размножения рыб включает в качестве элемента изменение пола особями одного вида, когда рыбы могут функционировать то как мужская, то как женская особь случайно или последовательно. Последовательное функционирование проявляется в виде функционирования как самцов на протяжении одной части жизни, и как самок — на протяжении другой. Существуют две формы последовательного изменения пола — протоандрия и протогения. Протоандрические гермафродиты — это особи, которые в начале своей жизни являются самцами, а позднее претерпевают кардинальные перестройки половой системы и становятся полностью функциональными самками. Такая форма преобразования пола широко распространена в семействе морских окуней (Serranidae). Все губаны (Labridae) являются протогеническими гермафродитами, когда все самцы являются преобразованными с возрастом самками. В этом семействе на смену пола могут влиять как факторы окружающей среды, так и социальные отношения в популяции. Социальная структура губанов заключается в наличии гаремов, которые состоят из самок и одного большого самца. Внутри группа структурирована по размеру, с самцом на верхушке иерархии. Если изъять из группы самку, другие самки (низшие по рангу) будут изменять свое иерархичное положение, обычно сдвигаясь на одну позицию вверх. Если же изъять из группы самца, самая крупная самка гарема старается занять его место, агрессивно отгоняя самцов, которые контролируют другие гаремы. Если ей это удается, и никому из окружающих самцов не удается присоединить этот гарем к собственному, то эта самка начинает демонстрировать поведение самца, и после около 14 дней ее половая система полностью изменяется, начиная продуцировать мужские половые клетки.
В таксонах, где половая принадлежность обусловлена социальной структурой, процесс изменения пола широко варьирует, и одна и та же особь может изменять пол несколько раз на протяжении жизни. С другой стороны, существуют таксоны (например, полосатые окуни, желтый окунь, большинство групперов) где половая принадлежность особей чередуется, но не испытает влияние социальной структуры. Случайные гермафродиты могут продуцировать как яйцеклетки, так и сперматозоиды — они потенциально имеют возможность самооплодотворения. Известные лишь три вида из отряда Cyprinodontiformes, которые функционируют как самооплодотворяющие гермафродиты: два вида рода Cynolebias и вид Rivulus marmoratus. При этом самооплодотворение у Rivulus marmoratus является внутренним, и в результате приводит к появлению гомозиготных, генетически идентичных потомков. Более обычная форма случайного гермафродитизма наблюдается в родах Hypoplectrus и Serranus семейства Окуневых (Percidae). Хотя эти рыбы способны продуцировать сперматозоиды и яйцеклетки одновременно, на протяжении одного нереста они функционируют как представители только одного пола. Учитывая то, что один акт нереста может длиться несколько часов, рыбы одной пары могут обмениваться половыми ролями, и продуцировать поочередно яйцеклетки (икру) или сперматозоиды (молоки).
Партеногенез
Несмотря на редкость этого типа размножения среди позвоночных, несколько видов рыб прибегают к нему. По определению, партеногенез заключается в развитии яйца без оплодотворения сперматозоидом этого же вида. У рыб существует вариант этого типа размножения, при котором необходимым является общий нерест с самцами того же самого вида или других видов. При этом роль самцов заключается в продуцировании сперматозоидов, которые контактируют с икринками, но не в состоянии проникнуть через их внешнюю мембрану (хорион). Контакт со сперматозоидами выполняет роль стимула, который побуждает яйцо начать развитие. При этом сперматозоиды не вносят в яйцо своего генетического материала, то есть все потомки при таком размножении будут самками, генетически идентичными с материнской особью. Классическим примером такого гермафродитизма есть гольяны (род Poeciliopsis) и европейский серебряный карась Carassius gibelio.
Формы размножения
В дополнение к трем типам размножения существуют три его разные формы внутри каждого типа: яйцерождение, яйцеживорождение и живорождение/
При яйцерождении мужские и женские гаметы выметываются в воду, где происходит оплодотворение. Иногда оплодотворение может быть и внутренним (как у скорпен (Scorpenidae) и американских сомов Auchenipteridae), но самка после этого выметывает оплодотворенную икру. На протяжении развития зародыш использует как желток яйца, так и имеющиеся в яйце капли жиров.
При яйцеживорождении яйца все время находятся в организме самки, оплодотворение внутреннее. Несмотря на нахождение яиц в теле самки, между материнским организмом и зародышем не возникает плацентарного контакта или контакта кругов кровообращения. Вместе с тем зародыш во время своего развития питается запасами питательных веществ, которые имеются в яйце. Мальки выходят из яиц внутри материнского организма, после чего немедленно выходят наружу. Наиболее известными рыбами, которым присуща такая форма размножения, являются представители Poeciliidae — широко распространенные в аквариумах гуппи и меченосцы. Также данная форма размножения характерна для латимерии.
Размножение в форме живорождения во многом подобно яйцеживорождению, но при этом между яйцом и материнским организмом возникает плацентарный контакт, или контакт между их кругами кровообращения. Таким образом, эмбрион при развитии получает необходимые питательные вещества из организма матери. Наиболее обычна данная форма размножения для акул, но она присущая также некоторым и костистым рыбам — например, жителям горных водоемов из семейства Goodeidae и представителям семейства Embiotocidae.
Забота о потомстве
Нехарактерная для рыб забота о потомстве наблюдается преимущественно у видов в приливно-отливной зоне, в узких заливах и бухтах, а также в реках и озерах.
Самец пятнадцатииглой морской колюшки сооружает своеобразное гнездо из кусочков водорослей, скрепляя их специальным секретом. Потом он загоняет в гнездо самок, которые откладывают икру, а самец остаётся охранять икру и молодь. Самка горчака может вытягивать своеобразный яйцеклад в длинную трубку и откладывает икру в мантийную полость пресноводных двустворчатых моллюсков. Самец выпускает сперму рядом с моллюском, и, когда тот захватывает корм, втягивая в себя воду, и икра оплодотворяется.
Особое приспособление имеется у самца австралийского гулливерова куртуса. Самец вынашивает икринки у себя на лбу. Грозди икринок, прикрепленных клейкими нитевидными выростами с обоих концов, крепко удерживаются крючком, образованным лучами его спинного плавника.
Сложное поведение, связанное с заботой о потомстве, развилось у морских игл и морских коньков. Обитающие в бассейне реки Амазонки дискусы выкармливают малька выделениями на боках.
Классификация
С таксономической точки зрения рыбы — парафилетическая группа, так как в неё включают кистеперых рыб, ближайших общих предков наземных позвоночных — четвероногих (Tetrapoda), которые, очевидно, рыбами не считаются.
Делятся на три современных класса: хрящевые, лучепёрые и лопастепёрые рыбы. Отношения основных подгрупп рыб представлены ниже:
- Рыбы (Pisces)
- Класс Панцирные рыбы или плакодермы (Placodermi) †
- Класс Хрящевые рыбы (Chondrichthyes)
- Класс Акантоды (Acanthodii) †
- Класс Костные рыбы (Osteichthyes)
- Подкласс Лучепёрые рыбы (Actinopterygii)
- Подкласс Лопастепёрые рыбы (Sarcopterygii)
- Надотряд Кистепёрые рыбы (Crossopterygii)
- Надотряд Двоякодышащие (Dipnoi)
Экология рыб
Внешней средой для рыб является вода, которая занимает огромные пространства. Около 361 млн.км², или 71 % всей поверхности Земного шара, занято морями и океанами и 2,5 млн.км² — внутренними водоемами. Они распространены от высокогорных водоёмов (более 6000 м над уровнем океана) до наибольших глубин Мирового океана (11034 м). Встречаются рыбы в полярных водах и тропических зонах. В настоящее время рыбы — господствующая группа животных в водных биоценозах. Наряду с китообразными, они завершают цепи питания. Рыбы приспособились к различным условиям водной среды, которая как фактор, влияющий на рыб, делится на абиотическую и биотическую.
Биотические факторы — это живые организмы, окружающие рыбу и вступающие с ней в различные взаимоотношения.
Абиотические факторы — это физико-химические свойства среды обитания. К наиболее важным абиотическим факторам относятся температура воды, солёность, содержание газов и др.
По местам обитания различают морских, пресноводных и проходных рыб.
При достаточно большом разнообразии видов по образу жизни всех рыб можно включить в состав нескольких экотипов:
- литоральные (планктон) — обитают в прибрежной зоне (бычки, морские собачки).
- пелагические рыбы (нектон) — держатся в толще воды;
- донные (нектобентос) — например, скаты, камбалы, сомы;
Многие виды содержатся в неволе — в аквариумах. Содержание и разведение аквариумных рыб в некоторых случаях помогает сохранить редкие виды, вымирающие в исконных местах обитания.
Экономическое значение и угрозы
В экономическом отношении значительную роль играет рыболовный промысел. Другой отраслью является торговля декоративными рыбами. И то, и другое способно ставить под угрозу популяции тех или иных видов. Другими угрозами являются загрязнение воды, перегорождение рек, потепление, интродуцирование чужих видов, а также высыхание рек.
Прочие факты
- Рыба — один из ранних символов христианства. По-гречески «рыба» — Ίχθύς, «ichthys», что является акронимом греческой фразы «Ἰησοὺς Χριστὸς Θεoὺ ῾Υιὸς Σωτήρ», что переводится на русский как «Иисус Христос Божий Сын Спаситель». Эта фраза встречается на каменных плитах — святых реликвиях в Иерусалиме. Изображение двух рыб по сторонам от вертикального якоря с верхушкой в виде креста использовалось как секретный «пропуск-пароль» во времена, когда первые христиане преследовались римлянами.
См. также
- Всероссийский научно-исследовательский институт рыбного хозяйства и океанографии
- Список семейств рыб
- Рыбы — символы штатов США
Примечания
Литература
- Жизнь животных. Энциклопедия в шести томах. Том 4. Часть первая. (Рыбы). Общая редакция члена-корреспондента АН СССР профессора Л. А. Зенкевича. М. Просвещение, 1971. 656 стр.
- Павлов Д. А. Морфологическая изменчивость в раннем онтогенезе костистых рыб — М.: ГЕОС, 2007. — 262 с.
- Павлов Д. С., Лупандин А. И., Костин В. В. Механизмы покатной миграции молоди речных рыб — М.: Наука, 2007. — 212 с.
- Попов П. А. Рыбы Сибири: распространение, экология, вылов — Новосибирск, 2007. — 525 с.
- Баклашова Т. А. Ихтиология — М.: Пищевая пром-сть, 1980. — 324 с.
Ссылки
Удивительные факты из жизни рыб
- FishBase // WorldFish Center — БД по рыбам мира (на декабрь 2006 — 29600 видов, 222490 народных названия, 43600 изображения, 39200 ссылок)
- Стадии зрелости половых желез у рыб — ИнтерНевод
- Энциклопедия рыб
Категории:- Животные по алфавиту
- Рыбы
- Парафилетические группы











Wikimedia Foundation. 2010.