- Метаболизм
-
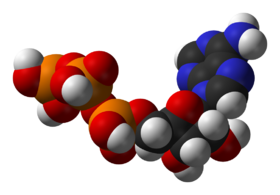 Структура аденозинтрифосфата — главного посредника в энергетическом обмене веществ
Структура аденозинтрифосфата — главного посредника в энергетическом обмене веществ
Метаболи́зм (от греч. μεταβολή — «превращение, изменение»), или обмен веществ — набор химических реакций, которые возникают в живом организме для поддержания жизни. Эти процессы позволяют организмам расти и размножаться, сохранять свои структуры и отвечать на воздействия окружающей среды. Метаболизм обычно делят на две стадии: в ходе катаболизма сложные органические вещества деградируют до более простых; в процессах анаболизма с затратами энергии синтезируются такие вещества, как белки, сахара, липиды и нуклеиновые кислоты.
Обмен веществ происходит между клетками организма и межклеточной жидкостью, постоянство состава которой поддерживается кровообращением: за время прохождения крови в капиллярах через проницаемые стенки капилляров плазма крови 40 раз полностью обновляется с интерстициальной жидкостью. Серии химических реакций обмена веществ называют метаболическими путями, в них при участии ферментов одни биологически значимые молекулы последовательно превращаются в другие. Ферменты играют важную роль в метаболических процессах потому, что:
- действуют как биологические катализаторы и снижают энергию активации химической реакции;
- позволяют регулировать метаболические пути в ответ на изменения среды клетки или сигналы от других клеток.
Особенности метаболизма влияют на то, будет ли пригодна определенная молекула для использования организмом в качестве источника энергии. Так, например, некоторые прокариоты используют сероводород в качестве источника энергии, однако этот газ ядовит для животных.[1] Скорость обмена веществ также влияет на количество пищи, необходимой для организма.
Основные метаболические пути и их компоненты одинаковы для многих видов, что свидетельствует о единстве происхождения всех живых существ.[2] Например, некоторые карбоновые кислоты, являющиеся интермедиатами цикла трикарбоновых кислот присутствуют во всех организмах, начиная от бактерий и заканчивая многоклеточными организмами эукариот.[3] Сходства в обмене веществ, вероятно, связаны с высокой эффективностью метаболических путей, а также с их ранним появлением в истории эволюции.[4][5]
Содержание
Биологические молекулы
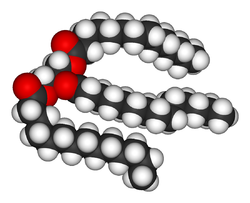 Структура липида триглицерида
Структура липида триглицеридаОрганические вещества, входящие в состав всех живых существ (животных, растений, грибов и микроорганизмов), представлены в основном аминокислотами, углеводами, липидами (часто называемые жирами) и нуклеиновыми кислотами. Так как эти молекулы имеют важное значение для жизни, метаболические реакции сосредоточены на создании этих молекул при строительстве клеток и тканей или разрушении их с целью использования в качестве источника энергии. Многие важные биохимические реакции объединяются вместе для синтеза ДНК и белков.
Тип молекулы Название формы мономера Название формы полимера Примеры форм полимера Аминокислоты Аминокислоты Белки (полипептиды) Фибриллярные и глобулярные белки Углеводы Моносахариды Полисахариды Крахмал, гликоген, целлюлоза Нуклеиновые кислоты Нуклеотиды Полинуклеотиды ДНК и РНК Аминокислоты и белки
Белки являются линейными биополимерами и состоят из остатков аминокислот, соединённых пептидными связями. Некоторые белки являются ферментами и катализируют химические реакции. Другие белки выполняют структурную или механическую функцию (например, образуют цитоскелет).[6] Белки также играют важную роль в передаче сигнала в клетках, иммунных реакциях, агрегации клеток, активном транспорте через мембраны и регуляции клеточного цикла.[7]
Липиды
Липиды входят в состав биологических мембран, например, плазматических мембран, являются компонентами коферментов и источниками энергии.[7] Липиды являются гидрофобными или амфифильными биологическими молекулами, растворимыми в органических растворителях таких, как бензол или хлороформ.[8] Жиры — большая группа соединений, в состав которых входят жирные кислоты и глицерин. Молекула трёхатомного спирта глицерина, образующая три сложные эфирные связи с тремя молекулами жирных кислот, называется триглицеридом.[9] Наряду с остатками жирных кислот, в состав сложных липидов может входить, например, сфингозин (сфинголипиды), гидрофильные группы фосфатов (в фосфолипидах). Стероиды, например холестерол, представляют собой ещё один большой класс липидов.[10]
Углеводы
Сахара могут существовать в кольцевой или линейной форме в виде альдегидов или кетонов, имеют несколько гидроксильных групп. Углеводы являются наиболее распространёнными биологическими молекулами. Углеводы выполняют следующие функции: хранение и транспортировка энергии (крахмал, гликоген), структурная (целлюлоза растений, хитин у животных).[7] Наиболее распространенными мономерами сахаров являются гексозы — глюкоза, фруктоза и галактоза. Моносахариды входят в состав более сложных линейных или разветвленных полисахаридов.[11]
Нуклеотиды
Полимерные молекулы ДНК и РНК представляют собой длинные неразветвленные цепочки нуклеотидов. Нуклеиновые кислоты выполняют функцию хранения и реализации генетической информации, которые осуществляются в ходе процессов репликации,транскрипции, трансляции, и биосинтеза белка.[7] Информация, закодированная в нуклеиновых кислотах, защищается от изменений системами репарации и мультиплицируется при помощи репликации ДНК.
Некоторые вирусы имеют РНК-содержащий геном. Например, вирус иммунодефицита человека использует обратную транскрипцию для создания матрицы ДНК из собственного РНК-содержащего генома.[12] Некоторые молекулы РНК обладают каталитическими свойствами (рибозимы) и входят в состав сплайсосом и рибосом.
Нуклеозиды — продукты присоединения азотистых оснований к сахару рибозе. Примерами азотистых оснований являются гетероциклические азотсодержащие соединения — производные пуринов и пиримидинов. Некоторые нуклеотиды также выступают в качестве коферментов в реакциях переноса функциональных групп.[13]
Коферменты
- Подробное рассмотрение темы: Коферменты
Метаболизм включает широкий спектр химических реакций, большинство из которых относятся к нескольким основным типам реакций переноса функциональных групп.[14] Для переноса функциональных групп между ферментами, катализирующими химические реакции, используются коферменты.[13] Каждый класс химических реакций переноса функциональных групп катализируется отдельными ферментами и их кофакторами.[15]
Аденозинтрифосфат (АТФ) — один из центральных коферментов, универсальный источник энергии клеток. Этот нуклеотид используется для передачи химической энергии, запасенной в макроэргических связях между различными химическими реакциями. В клетках существует небольшое количество АТФ, который постоянно регенерируется из ADP и AMP. Организм человека за сутки расходует массу АТФ, равную массе собственного тела.[15] АТР выступает в качестве связующего звена между катаболизмом и анаболизмом: при катаболических реакциях образуется АТФ, при анаболических — энергия потребляется. АТФ также выступает донором фосфатной группы в реакциях фосфорилирования.
Витамины — низкомолекулярные органические вещества, необходимые в небольших количествах, причём, например, у человека большинство витаминов не синтезируется, а получается с пищей или через микрофлору КТ. В организме человека большинство витаминов являются кофакторами ферментов. Большинство витаминов приобретают биологическую активность в измененном виде, например, все водорастворимые витамины в клетках фосфорилируются или соединяются с нуклеотидами.[16] Никотинамидадениндинуклеотид (NADH) является производным витамина B3 (ниацина), и представляет собой важный кофермент — акцептора водорода. Сотни различных ферментов дегидрогеназ отнимают электроны из молекул субстратов и переносят их на молекулы NAD+, восстанавливая его до NADH. Окисленная форма кофермента является субстратом для различных редуктаз в клетке.[17] NAD в клетке существует в двух связанных формах NADH и NADPH. NAD+/NADH больше важен для протекания катаболических реакций, а NADP+/NADPH чаще используется в анаболических реакциях.
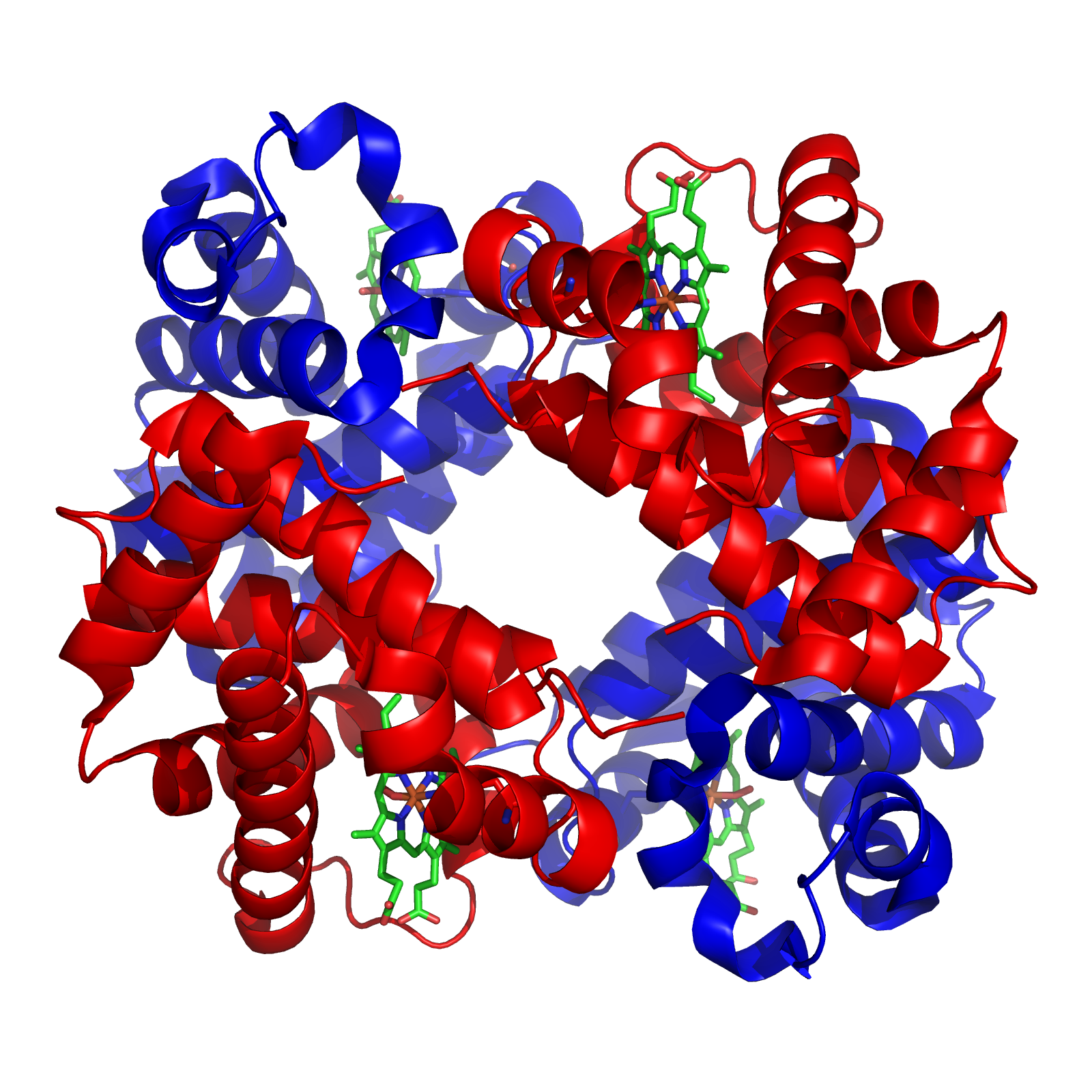
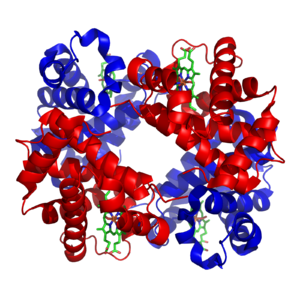 Структура гемоглобина. Белковые субъединицы окрашены красным и синим, а железосодержащий гем — зелёным. Из PDB 1GZX.
Структура гемоглобина. Белковые субъединицы окрашены красным и синим, а железосодержащий гем — зелёным. Из PDB 1GZX.Минералы и кофакторы
Неорганические элементы играют важнейшую роль в обмене веществ. Около 99 % массы млекопитающего состоит из углерода, азота, кальция, натрия, магния, хлора, калия, водорода, фосфора, кислорода и серы.[18] Биологически значимые органические соединения (белки, жиры, углеводы и нуклеиновые кислоты) содержат большое количество углерода, водорода, кислорода, азота и фосфора.[18]
Многие неорганические соединения являются ионными электролитами. Наиболее важны для организма ионы натрия, калия, кальция, магния, хлоридов, фосфатов и гидрокарбонатов. Баланс этих ионов внутри клетки во внеклеточной среде определяет осмотическое давление и рН.[19] Концентрации ионов также играют важную роль для функционирования нервных и мышечных клеток. Потенциал действия в возбудимых тканях возникает при обмене ионами между внеклеточной жидкостью и цитоплазмой.[20] Электролиты входят и выходят из клетки через ионные каналы в плазматической мембране. Например, в ходе мышечного сокращения в плазматической мембране, цитоплазме и Т-трубочках перемещаются ионы кальция, натрия и калия.[21]
Переходные металлы в организме являются микроэлементами, наиболее распространены цинк и железо.[22][23] Эти металлы используются некоторыми белками (например, ферментами в качестве кофакторов) и имеют важное значение для регуляции активности ферментов и транспортных белков.[24] Кофакторы ферментов обычно прочно связаны со специфическим белком, однако могут модифицироваться в процессе катализа, при этом после окончания катализа всегда возвращаются к своему первоначальному состоянию (не расходуются). Металлы-микроэлементы усваиваются организмом при помощи специальных транспортных белков и не встречаются в организме в свободном состоянии, так как связаны со специфическими белками-переносчиками (например, ферритином или металлотионеинами).[25][26]
Катаболизм
Катаболизмом называют метаболические процессы, при которых расщепляются относительно крупные органические молекулы сахаров, жиров, аминокислот. В ходе катаболизма образуются более простые органические молекулы, необходимые для реакций анаболизма (биосинтеза). Часто, именно в ходе реакций катаболизма организм мобилизует энергию, переводя энергию химических связей органических молекул, полученных в процессе переваривания пищи, в доступные формы: в виде АТФ, восстановленных коферментов и трансмембранного электрохимического потенциала. Термин катаболизм не является синонимом «энергетического обмена»: у многих организмов (например, у фототрофов) основные процессы запасания энергии не связаны напрямую с расщеплением органических молекул. Классификация организмов по типу метаболизма может быть основана на источнике получения энергии и углерода, что отражено в таблице ниже. Органические молекулы используются в качестве источника энергии органотрофами, литотрофы используют неорганические субстраты, а фототрофы потребляют энергию солнечного света. Однако, все эти различные формы обмена веществ зависят от окислительно-восстановительных реакций, которые связаны с передачей электронов от восстановленных доноров молекул, таких как органические молекулы, вода, аммиак, сероводород, на акцепторные молекулы, такие как кислород, нитраты или сульфат.[27] У животных эти реакции сопряжены с расщеплением сложных органических молекул до более простых, таких как двуокись углерода и воду. В фотосинтезирующих организмах — растениях и цианобактериях — реакции переноса электрона не высвобождают энергию, но они используются как способ запасания энергии, поглощаемой из солнечного света.[28]
Классификация организмов на основе их метаболизма Источник энергии Солнечный свет фототрофы Первичные молекулы хемотрофы Донор электронов Органическое соединение органотрофы Неорганическое соединение литотрофы Источник углерода Органическое соединение гетеротрофы Неорганическое соединение автотрофы Катаболизм у животных может быть разделён на три основных этапа. Во-первых, крупные органические молекулы, такие как белки, полисахариды и липиды расщепляются до более мелких компонентов вне клеток. Далее эти небольшие молекулы попадают в клетки и превращается в ещё более мелкие молекулы, например, ацетил-КоА. В свою очередь, ацетильная группа кофермента А окисляется до воды и углекислого газа в цикле Кребса и дыхательной цепи, высвобождая при этом энергию, которая запасается в форме АТР.
Пищеварение
- Подробное рассмотрение темы: Пищеварение и Желудочно-кишечный тракт
Такие макромолекулы, как крахмал, целлюлоза или белки, должны расщепляться до более мелких единиц прежде, чем они могут быть использованы клетками. Несколько классов ферментов принимают участие в деградации: протеазы, которые расщепляют белки до пептидов и аминокислот, гликозидазы, которые расщепляют полисахариды до олиго- и моносахаридов.
Микроорганизмы выделяют гидролитические ферменты в пространство вокруг себя,[29][30] чем отличаются от животных, которые выделяют такие ферменты только из специализированных железистых клеток.[31] Аминокислоты и моносахариды, образующиеся в результате активности внеклеточных ферментов, затем поступают в клетки с помощью активного транспорта.[32][33]
Получение энергии
- Подробное рассмотрение темы: Клеточное дыхание, Брожение, Липолиз
В ходе катаболизма углеводов сложные сахара расщепляются до моносахаридов, которые усваиваются клетками.[34] Попав внутрь, сахара (например, глюкоза и фруктоза) в процессе гликолиза превращаются в пируват, при этом вырабатывается некоторое количество АТР.[35] Пировиноградная кислота (пируват) является промежуточным продуктом в нескольких метаболических путях. Основной путь метаболизма пирувата — превращаение в ацетил-КоА и далее поступление в цикл трикарбоновых кислот. При этом в цикле Кребса в форме АТР запасается часть энергии, а также восстанавливаются молекулы NADH и FAD. В процессе гликолиза и цикла трикарбоновых кислот образуется диоксид углерода, который является побочным продуктом жизнедеятельности. В анаэробных условиях в результате гликолиза из пирувата при участии фермента лактатдегидрогеназы образуется лактат, и происходит окисление NADH до NAD+, который повторно используется в реакциях гликолиза. Существует также альтернативный путь метаболизма моносахаридов — пентозофосфатный путь, в ходе реакций которого энергия запасается в форме восстановленного кофермента NADPH и образуются пентозы, например, рибоза, необходимая для синтеза нуклеиновых кислот.
Жиры на первом этапе катаболизма гидролизуются в свободные жирные кислоты и глицерин. Жирные кислоты расщепляются в процессе бета-окисления с образованием ацетил-КоА, который в свою очередь далее катаболизируется в цикле Кребса, либо идет на синтез новых жирных кислот. Жирные кислоты выделяют больше энергии, чем углеводы, так как жиры содержат удельно больше атомов водорода в своей структуре.
Аминокислоты либо используются для синтеза белков и других биомолекул, либо окисляются до мочевины, диоксида углерода и служат источником энергии.[36] Окислительный путь катаболизма аминокислот начинается с удаления аминогруппы ферментами трансаминазами. Аминогруппы утилизируются в цикле мочевины; аминокислоты, лишённые аминогрупп называют кетокислотами. Некоторые кетокислоты — промежуточные продукты цикла Кребса. Например, при дезаминировании глутамата образуется альфа-кетоглутаровая кислота.[37] Гликогенные аминокислоты также могут быть преобразованы в глюкозу в реакциях глюконеогенеза.[38]
Энергетические превращения
Окислительное фосфорилирование
- Подробное рассмотрение темы: Окислительное фосфорилирование, Хемиосмос и Митохондрия
При окислительном фосфорилировании электроны, удалённые из пищевых молекул в метаболических путях (например, в цикле Кребса), переносятся на кислород, а выделяющаяся энергия используется для синтеза АТР. У эукариот данный процесс осуществляется при участии ряда белков, закреплённых в мембранах митохондрий, называемые дыхательной цепью переноса электронов. У прокариот эти белки присутствуют во внутренней мембране клеточной стенки.[39] Белки цепи переноса электронов используют энергию, полученную при передаче электронов от восстановленных молекул (например NADH) на кислород, для перекачки протонов через мембрану.[40]
При перекачке протонов создаётся разница концентраций ионов водорода и возникает электрохимический градиент.[41] Эта сила возвращает протоны обратно в митохондрии через основание АТР-синтазы. Поток протонов заставляет вращаться кольцо из c-субъединиц фермента, в результате чего активный центр синтазы изменяет форму и фосфорилирует аденозиндифосфат, превращая его в АТР.[15]
Энергия из неорганических соединений
Хемолитотрофами называют прокариот, имеющих особый тип обмена веществ, при котором энергия образуется в результате окисления неорганических соединений. Хемолитотрофы могут окислять молекулярный водород,[42] соединения серы (например, сульфиды, сероводород и тиосульфат),[1] оксид железа(II)[43] или аммиак.[44] При этом энергия от окисления этих соединений образуется с помощью акцепторов электронов, таких как кислород или нитриты.[45] Процессы получения энергии из неорганических веществ играют важную роль в таких биогеохимических циклах, как ацетогенез, нитрификация и денитрификация.[46][47]
Энергия из солнечного света
Энергия солнечного света поглощается растениями, цианобактериями, пурпурными бактериями, зелёными серными бактериями и некоторыми простейшими. Этот процесс часто сочетается с превращением диоксида углерода в органические соединения, как часть процесса фотосинтеза (см. ниже). Системы захвата энергии и фиксации углерода у некоторых прокариот могут работать раздельно (например, у пурпурных и зелёных серных бактерий).[48][49]
У многих организмов поглощение солнечной энергии в принципе аналогично окислительному фосфорилированию, так как при этом энергия запасается в форме градиента концентрации протонов и движущая сила протонов приводит к синтезу АТР.[15] Электроны, необходимые для этой цепи переноса, поступают от светособирающих белков, называемых центрами фотосинтетических реакций (примером являются родопсины). В зависимости от вида фотосинтетических пигментов классифицируют два типа центров реакций; в настоящее время большинство фотосинтезирующих бактерий имеют только один тип, в то время как растения и цианобактерии два.[50]
У растений, водорослей и цианобактерий, фотосистема II использует энергию света для удаления электронов из воды, при этом молекулярный кислород выделяется как побочный продукт реакции. Электроны затем поступают в комплекс цитохрома b6f, который использует энергию для перекачки протонов через тилакоидную мембрану в хлоропластах.[7] Под действием электрохимического градиента протоны движутся обратно через мембрану и запускают АТР-синтазу. Электроны затем проходят через фотосистему I и могут быть использованы для окисления кофермента NADP+, для использования в цикле Кальвина или рециркуляции для образования дополнительных молекул АТР.[51]
Анаболизм
- Подробное рассмотрение темы: Анаболизм
Анаболизм — совокупность метаболических процессов биосинтеза сложных молекул с затратой энергии. Сложные молекулы, входящие в состав клеточных структур, синтезируются последовательно из более простых предшественников. Анаболизм включает три основных этапа, каждый из которых катализируется специализированным ферментом. На первом этапе синтезируются молекулы-предшественники, например, аминокислоты, моносахариды, терпеноиды и нуклеотиды. На втором этапе предшественники с затратой энергии АТР преобразуются в активированные формы. На третьем этапе активированные мономеры объединяются в более сложные молекулы, например, белки, полисахариды, липиды и нуклеиновые кислоты.
Не все живые организмы могут синтезировать все биологически активные молекулы. Автотрофы (например, растения) могут синтезировать сложные органические молекулы из таких простых неорганических низкомолекулярных веществ, как углекислый газ и вода. Гетеротрофам необходим источник более сложных веществ, таких как моносахариды и аминокислоты, для создания более сложных молекул. Организмы классифицируют по их основным источникам энергии: фотоавтотрофы и фотогетеротрофы получают энергию из солнечного света, в то время как хемоавтотрофы и хемогетеротрофы получают энергию из неорганических реакций окисления.
Связывание углерода
- Подробное рассмотрение темы: Фотосинтез и Хемосинтез
 Растительные клетки содержат хлоропласты (зелёного цвета), в тилакоидах которых происходят процессы фотосинтеза. Plagiomnium affine из семейства Mniaceae отдела Настоящие мхи (Bryophyta)
Растительные клетки содержат хлоропласты (зелёного цвета), в тилакоидах которых происходят процессы фотосинтеза. Plagiomnium affine из семейства Mniaceae отдела Настоящие мхи (Bryophyta)Фотосинтезом называют процесс биосинтеза сахаров из углекислого газа, при котором необходимая энергия поглощается из солнечного света. У растений, цианобактерий и водорослей, при кислородном фотосинтезе происходит фотолиз воды, при этом, как побочный продукт, выделяется кислород. Для преобразования CO2 в 3-фосфоглицерат используется энергия АТР и NADPH, запасенная в фотосистемах. Реакция связывания углерода осуществляется с помощью фермента рибулозобисфосфаткарбоксилазы и является частью цикла Кальвина.[52] У растений классифицируют три типа фотосинтеза — по пути трехуглеродых молекул, по пути четырехуглеродых молекул (С4), и CAM фотосинтез. Три типа фотосинтеза отличаются по пути связывания углекислого газа и его вхождения в цикл Кальвина; у C3 растений связывание CO2 происходит непосредственно в цикле Кальвина, а при С4 и CAM CO2 предварительно включается в состав других соединений. Разные формы фотосинтеза являются приспособлениями к к интенсивному потоку солнечных лучей и к сухим условиям.[53]
У фотосинтезирующих прокариот механизмы связывания углерода более разнообразны. Углекислый газ может быть фиксирован в цикле Кальвина, в обратном цикле Кребса,[54] или в реакциях карбоксилирования ацетил-КоА.[55][56] Прокариоты — хемоавтотрофы также связывают CO2 через цикл Кальвина, но для протекания реакции используют энергию из неорганических соединений.[57]
Углеводы и гликаны
- Подробное рассмотрение темы: Глюконеогенез и Гликозилирование
В процессе анаболизма сахаров простые органические кислоты могут быть преобразованы в моносахариды, например, в глюкозу, и затем использованы для синтеза полисахаридов, таких как крахмал. Образование глюкозы из соединений, как пируват, лактат, глицерин, 3-фосфоглицерат и аминокислот называют глюконеогенезом. В процессе глюконеогенеза пируват превращается глюкозо-6-фосфат через ряд промежуточных соединений, многие из которых образуются и при гликолизе.[35] Однако, глюконеогенез не просто является гликолизом в обратном направлении, так как несколько химических реакций катализируют специальные ферменты, что дает возможность независимо регулировать процессы образования и распада глюкозы.[58][59]
Многие организмы запасают питательные вещества в форме липидов и жиров, однако, позвоночные не имеют ферментов, катализирующих превращение ацетил-КоА (продукта метаболизма жирных кислот) в пируват (субстрат глюконеогенеза).[60] После длительного голодания позвоночные начинают синтезировать кетоновые тела из жирных кислот, которые могут заменять глюкозу в таких тканях, как головной мозг.[61] У растений и бактерий, данная метаболическая проблема решается использованием глиоксилатного цикла, который обходит этап декарбоксилирования в цикле лимонной кислоты и позволяет превращать ацетил-КоА в оксалоацетат, и далее использовать для синтеза глюкозы.[60][62]
Полисахариды выполняют структурные и метаболические функции, а также могут быть соединены с липидами (гликолипиды) и белками (гликопротеиды) при помощи ферментов олигосахаридтрансфераз.[63][64]
Жирные кислоты, изопреноиды и стероиды
- Подробное рассмотрение темы: Стероиды
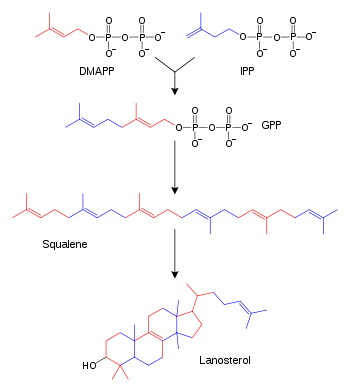 Синтез стероидов из изопентилпирофосфата, диметилаллилпирофосфата, геранилпирофосфата и сквалена. Некоторые промежуточные продукты не показаны
Синтез стероидов из изопентилпирофосфата, диметилаллилпирофосфата, геранилпирофосфата и сквалена. Некоторые промежуточные продукты не показаныЖирные кислоты образуются синтазами жирных кислот из ацетил-КоА. Углеродный скелет жирных кислот удлиняется в цикле реакций, в которых сначала присоединяется ацетильная группа, далее карбонильная группа восстанавливается до гидроксильной, затем происходит дегидратация и последующее восстановление. Ферменты биосинтеза жирных кислот классифицируют на две группы: у животных и грибов все реакции синтеза жирных кислот осуществляются одним многофункциональным белком I типа,[65] в пластидах растений и у бактерий каждый этап катализируют отдельные ферменты II типа.[66][67]
Терпены и терпеноиды являются представителями самого многочисленного класса растительных натуральных продуктов.[68] Представители данной группы веществ являются производными изопрена и образуются из активированных предшественников изопентилпирофосфата и диметилаллилпирофосфата, которые, в свою очередь, образуются в разных реакциях обмена веществ.[69] У животных и архей изопентилпирофосфат и диметилаллилпирофосфат синтезируются из ацетил-КоА в мевалонатном пути,[70] в то время как у растений и бактерий субстратами не-мевалонатного пути являются пируват и глицеральдегид-3-фосфат.[69][71] В реакциях биосинтеза стероидов молекулы изопрена объединяются и образуют сквалены, которые далее формируют циклические структуры с образованием ланостерола.[72] Ланостерол может быть преобразован в другие стероиды, например холестерин и эргостерин.[72][73]
Белки
- Подробное рассмотрение темы: Биосинтез белка
Организмы различаются по способности к синтезу 20 общих аминокислот. Большинство бактерий и растений могут синтезировать все 20, но млекопитающие способны синтезировать лишь 11 заменимых аминокислот.[7] Таким образом, в случае млекопитающих 9 незаменимых аминокислот должны быть получены из пищи. Все аминокислоты синтезируются из промежуточных продуктов гликолиза, цикла лимонной кислоты или пентозомонофосфатного пути. Перенос аминогрупп с аминокислот на альфа-кетокислоты называется трансаминированием. Донорами аминогрупп являются глутамат и глутамин. [74]
Аминокислоты, соединенными пептидными связями, образуют белки. Каждый белок имеет уникальную последовательность аминокислотных остатков (первичная структура белка). Подобно тому, как буквы алфавита могут комбинироваться с образованием почти бесконечных вариаций слов, аминокислоты могут связываться в той или иной последовательности и формировать разнообразные белки. Фермент Аминоацил-тРНК-синтетаза катализирует АТР-зависимое присоединение аминокислот к тРНК сложноэфирными связями, при этом образуются аминоацил-тРНК.[75] Аминоацил-тРНК являются субстратами для рибосом, которая объединяют аминокислоты в длинные полипептидные цепочки, используя матрицу мРНК.[76]
Нуклеотиды
Нуклеотиды образуются из аминокислот, углекислого газа и муравьиной кислоты в цепи реакций, для протекания которых требуется большое количество энергии.[77][78] Именно поэтому большинство организмов имеют эффективные системы сохранения ранее синтезированных нуклеотидов и азотистых оснований.[77][79] Пурины синтезируются как нуклеозиды (в основном связанные с рибозой). Аденин и гуанин образуются из инозин-монофосфата, который синтезируется из глицина, глутамина и аспартата при участии метенил-тетрагидрофолата. Пиримидины синтезируются из оротата, который образуется из глутамина и аспартата.[80]
Ксенобиотики и окислительный метаболизм
- Подробное рассмотрение темы: Антиоксиданты
Все организмы постоянно подвергаются воздействию соединений, накопление которых может быть вредно для клеток. Такие потенциально опасные чужеродные соединения называются ксенобиотиками.[81] Ксенобиотики, например синтетические лекарства и яды природного происхождения, детоксифицируются специализированными ферментами. У человека такие ферменты представлены, например, цитохром-оксидазами,[82] глюкуронилтрансферазой,[83] и глутатион S-трансферазой.[84] Эта система ферментов действует в три этапа: на первой стадии ксенобиотики окисляются, затем происходит конъюгирование водорастворимых групп в молекулы, далее модифицированные водорастворимые ксенобиотики могут быть удалены из клеток и метаболизированы перед их экскрецией. Описанные реакции играют важную роль в разложении микробами загрязняющих веществ и биоремедиации загрязнённых земель и разливов нефти.[85] Многие подобные реакции протекают при участии многоклеточных организмов, однако, ввиду невероятного разнообразия, микроорганизмы справляются с гораздо более широким спектром ксенобиотиков, чем многоклеточные организмы, и способны даже разрушать стойкие органические загрязнители, например хлорорганические соединения.[86]
Связанной с этим проблемой для аэробных организмов является оксидативный стресс.[87] В процессе окислительного фосфорилирования и образования дисульфидных связей при укладке белка образуются активные формы кислорода, например пероксид водорода.[88] Эти повреждающие оксиданты удаляются антиоксидантами, например глутатионом и ферментами каталазой и пероксидазами.[89][90]
Термодинамика живых организмов
Живые организмы подчиняются началам термодинамики, которые описывают превращения тепла и работы. Второе начало термодинамики гласит, что в любой изолированной системе энтропия не уменьшается. Хотя невероятная сложность живых организмов очевидно противоречит этому закону, жизнь возможна, так как все организмы открытые системы, которые обмениваются веществом и энергией с окружающей средой. Таким образом живые системы не находятся в термодинамическом равновесии, но вместо этого выступает диссипативной системой, которая поддерживают своё состояние сложно организованности, вызывая большее увеличение энтропии окружающей средой.[91] В метаболизм клеток это достигается путём сочетания спонтанных процессов катаболизма с не спонтанных процессов анаболизма. В термодинамических условиях, метаболизм поддерживает порядок за счёт создания беспорядка.[92]
Регуляция и контроль
- Подробное рассмотрение темы: Гормоны, Передача сигнала в клетке
Гомеостазом называют постоянство внутренней среды организма. Так как внешняя среда, окружающая большинство организмов, постоянно меняется, для поддержания постоянных условий внутри клеток, реакции обмена веществ должны точно регулироваться.[93][94] Регуляция метаболизма позволяет организмам отвечать на сигналы и активно взаимодействовать с окружающей средой.[95] В случае фермента, регуляция заключается в повышении и снижении его активности в ответ на сигналы. С другой стороны, фермент оказывает некоторый контроль над метаболическим путем, который определяется как эффект от изменения активности фермента на данный метаболический путь.[96]
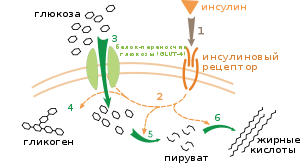 Влияние инсулина на поглощение глюкозы и обмен веществ. Инсулин связывается со своим рецептором (1), который в свою очередь запускает касакад реакций активации множества белков (2). К ним относятся: транслокация переносчика GLUT-4 к плазматической мембране и поступление глюкозы в клетку (3), синтез гликогена (4), гликолиз (5) и синтез жирных кислот (6).
Влияние инсулина на поглощение глюкозы и обмен веществ. Инсулин связывается со своим рецептором (1), который в свою очередь запускает касакад реакций активации множества белков (2). К ним относятся: транслокация переносчика GLUT-4 к плазматической мембране и поступление глюкозы в клетку (3), синтез гликогена (4), гликолиз (5) и синтез жирных кислот (6).Выделяют несколько уровней регуляции метаболизма. В метаболическом пути происходит саморегуляция на уровне субстрата или продукта; например, уменьшение количества продукта может компенсированно увеличить поток субстрата реакции по данному пути.[97] Этот тип регулирования часто включает аллостерическое регулирование активности некоторых ферментов в метаболических путях.[98] Внешний контроль включает клетку многоклеточного организма, изменяющую свой метаболизм в ответ на сигналы от других клеток. Эти сигналы, как правило, в виде растворимых мессенджеров, например гормоны и факторы роста, определяются специфическими рецепторами на поверхности клеток.[99] Затем эти сигналы передаются внутрь клетки системой вторичных мессенджеров, которые зачастую связаны с фосфорилированием белков.[100]
Хорошо изученный пример внешнего контроля — регуляция метаболизма глюкозы инсулином.[101] Инсулин вырабатывается в ответ на повышение уровня глюкозы в крови. Гормон связывается с инсулиновым рецептором на поверхности клетки, затем активируется каскад протеинкиназ, которые обеспечивают поглощение молекул глюкозы клетками и преобразовать их в молекулы жирных кислот и гликогена.[102] Метаболизм гликогена контролируется активностью фосфорилазы (фермента, который расщепляет гликоген) и гликогенсинтазы (фермента, который образует его). Эти ферменты взаимосвязаны; фосфорилирование ингибируется гликогенсинтазой, но активируется фосфорилазой. Инсулин вызывает синтез гликогена путём активации белковых фосфатаз и уменьшает фосфорилирование этих ферментов.[103]
Эволюция
- Подробное рассмотрение темы: Филогенетика
Главные пути метаболизма, описанные выше, например, гликолиза и цикла Кребса, присутствуют у всех трёх доменах живых существ и обнаруживаются у последнего универсального общего предка.[3][104] Этот универсальный предок был прокариотом и, вероятно, метаногеном с аминокислотным, нуклеотидным, углеводным и липидным метаболизмом.[105][106] Сохранение этих древних метаболических путей в эволюции может быть результатом того, что эти реакции оптимальны для решения конкретных проблем с метаболизмом. Так, конечные продукты гликолиза и цикла Кребса образуются с высокой эффективностью и с минимальным количеством стадий.[4][5] Первые метаболические пути на основе ферментов могли быть частями пуринового метаболизма нуклеотидов с предыдущим метаболических путей были частью древнего мира РНК.[107]
Многие модели были предложены для описания механизмов, посредством которых новые метаболические пути эволюционировали. К ним относятся последовательное добавление новых ферментов на короткий предковый путь, дупликация, а затем дивергенция всех путей, а также набор уже существующих ферментов и их сборка в новый путь реакций.[108] Относительную важность этих механизмов неясна, однако геномные исследования показали, что ферменты в метаболическом пути, скорее всего, имеют общее происхождение, предполагая, что многие пути эволюционировали шаг за шагом с новыми функциями, созданными из уже существующих этапов пути.[109] Альтернативная модель основана на исследованиях, в которых прослеживается эволюция структуры белков в метаболических связях; предполагают, что ферменты собирались для выполнения схожих функций в различных метаболических путях[110] Эти процессы сборки привели к эволюционированию ферментативной мозаики.[111] Некоторые части обмена веществ возможно существовали в качестве «модулей», которые могли быть повторно использованы в различных путях для выполнения схожих функций.[112]
Эволюция также может приводить к потере метаболических функций. Например, у некоторых паразитов метаболические процессы, которые не важны для выживания, утрачены и готовые аминокислоты, нуклеотиды и углеводы получаются от хозяина.[113] Подобные упрощения метаболических возможностей наблюдают у эндосимбиотических организмов.[114]
Методы исследования
- Подробное рассмотрение темы: Протеомика, Метабономика
 Метаболическая сеть цикла Кребса Arabidopsis thaliana. Ферменты и метаболиты обозначены красными квадратами, а взаимодействия между ними — как чёрные линии.
Метаболическая сеть цикла Кребса Arabidopsis thaliana. Ферменты и метаболиты обозначены красными квадратами, а взаимодействия между ними — как чёрные линии.Классически, метаболизм изучается упрощённым подходом, который фокусируется на одном метаболическом пути. Особенно ценно использование меченых атомов на организменном, тканевом и клеточном уровнях, которые определяют пути от предшественников до конечных продуктов путём выявления радиоактивно меченых промежуточных продуктов.[115] Ферменты, которые катализируют эти химические реакции, могут затем быть выделены для исследования их кинетики и ответа на ингибиторы. Параллельный подход заключается в выявлении небольших молекул в клетки или ткани; полный набор этих молекул называется метаболом. В целом, эти исследования дают хорошее представление о структуре и функциях простых путей метаболизма, но недостаточны в применении к более сложных системам, например полной метаболизм клетки.[116]
Идея сложности метаболических сетей в клетках, которые содержат тысячи различных ферментов, отражена на изображении справа, показывающее взаимодействия только между 43 белками и 40 метаболитами, которые регулируются 45000 генов.[117] Тем не менее, сейчас можно использовать такие данные о геномах для воссоздания полной сети биохимических реакций и образовывать более целостные математические модели, которые могут объяснить и предсказать их поведение.[118] Эти модели особенно сильны, когда используются для интеграции данных о путях и метаболитах, полученных на основе классических методов, с данными по экспрессии генов из протеомных и ДНК-микрочиповых исследований.[119] С помощью этих методов, модель человеческого метаболизма в настоящее время создаётся, которая будет служить ориентиром для будущих исследований лекарств и биохимических исследований.[120] Эти модели в настоящее время используются в анализах сети, для классификации болезней человека по группам, которые различаются по общим белкам или метаболитам.[121][122]
Яркий пример бактериальных метаболических сетей — устройство галстук-бабочки[123][124][125], структура которой позволяет вводить широкий спектр питательных веществ и производить большое разнообразие продуктов и сложных макромолекул, используя сравнительно немного общих промежуточных веществ.
Основная технологическая основа этой информации — метаболическая инженерия. Здесь организмы, например дрожжи, растения или бактерии, генетически модифицируются, чтобы сделать их более эффективными в биотехнологии и помочь в производстве лекарств, например антибиотиков или промышленных химических веществ, таких как 1,3-пропандиола и шикимовой кислоты.[126] Эти генетические модификации обычно направлены на уменьшение количества энергии, используемой для производства продукции, повышения урожайности и снижения производственных отходов.[127]
История
 Санторио взвешивает сам себя до и после принятия пищи, из Ars de statica medicina, впервые опубликованной в 1614 году
Санторио взвешивает сам себя до и после принятия пищи, из Ars de statica medicina, впервые опубликованной в 1614 годуИстория изучения метаболизма охватывает несколько столетий. Исследования начинались с изучения организмов животных, в современной биохимии изучают отдельные метаболические реакции. Понятие обмена веществ впервые встречается в работах Ибн аль-Нафиса (1213—1288), который писал, что «тело и его части находятся в постоянном состоянии распада и питания, так что оно неизбежно претерпевает постоянные изменения».[128] Первые контролируемые эксперименты по метаболизму у человека были опубликованы Санторио Санторио в 1614 году в книге итал. Ars de statica medicina.[129] Он рассказал, как он сам взвесил себя до и после приёма пищи, сна, работы, секса, натощак, после питья и выделения мочи. Он обнаружил, что большая часть пищи, которую он принял, была утрачена в результате процесса, названного «незаметным испарением».
В ранних исследованиях механизмы метаболических реакций не были обнаружены и считалось, что живой тканью управляет живая сила.[130] В XIX веке при исследовании ферментации сахара спирта дрожжами Луи Пастер сделал вывод, что брожение катализируется веществами из дрожжевых клеток, которые он назвал ферментами. Пастер писал, что «алкогольное брожение — действие, связанное с жизнью и организуется дрожжевыми клетками, не связано со смертью или разложением клеток».[131] Это открытие, вместе с публикацией Фридриха Вёлера в 1828 году о химическом синтезе мочевины,[132] доказали, что органические соединения и химические реакции, обнаруженные в клетках, не имеют различий в принципе, как и любые другие разделы химии.
Открытие ферментов в начале XX века Эдуардом Бухнером разделило изучение метаболических реакций от изучения клеток и дало начало развитию биохимии как науки.[133] Одним из успешных биохимиков начала двадцатого века был Ханс Адольф Кребс, который внёс огромный вклад в изучение метаболизма.[134] Кребс описал цикл мочевины и позднее, работая вместе с Хансом Корнбергом, цикл лимонной кислоты и глиоксилатный цикл.[135][62] В современных биохимических исследованиях широко используют новые методы, такие как хроматография, рентгеноструктурный анализ, ЯМР-спектроскопия, электронная микроскопия и метод классической молекулярной динамики. Эти методы позволяют открывать и подробно изучать множество молекул и метаболических путей в клетках.
См. также
Примечания
- ↑ 1 2 Friedrich C (1998). «Physiology and genetics of sulfur-oxidizing bacteria». Adv Microb Physiol 39: 235–89. DOI:10.1016/S0065-2911(08)60018-1. PMID 9328649.
- ↑ Pace NR (January 2001). «The universal nature of biochemistry». Proc. Natl. Acad. Sci. U.S.A. 98 (3): 805–8. DOI:10.1073/pnas.98.3.805. PMID 11158550.
- ↑ 1 2 Smith E, Morowitz H (2004). «Universality in intermediary metabolism». Proc Natl Acad Sci USA 101 (36): 13168–73. DOI:10.1073/pnas.0404922101. PMID 15340153.
- ↑ 1 2 Ebenhöh O, Heinrich R (2001). «Evolutionary optimization of metabolic pathways. Theoretical reconstruction of the stoichiometry of ATP and NADH producing systems». Bull Math Biol 63 (1): 21–55. DOI:10.1006/bulm.2000.0197. PMID 11146883.
- ↑ 1 2 Meléndez-Hevia E, Waddell T, Cascante M (1996). «The puzzle of the Krebs citric acid cycle: assembling the pieces of chemically feasible reactions, and opportunism in the design of metabolic pathways during evolution». J Mol Evol 43 (3): 293–303. DOI:10.1007/BF02338838. PMID 8703096.
- ↑ Michie K, Löwe J (2006). «Dynamic filaments of the bacterial cytoskeleton». Annu Rev Biochem 75: 467–92. DOI:10.1146/annurev.biochem.75.103004.142452. PMID 16756499.
- ↑ 1 2 3 4 5 6 Nelson David L. Lehninger Principles of Biochemistry. — New York: W. H. Freeman and company, 2005. — P. 841. — ISBN 0-7167-4339-6
- ↑ Fahy E, Subramaniam S, Brown H, Glass C, Merrill A, Murphy R, Raetz C, Russell D, Seyama Y, Shaw W, Shimizu T, Spener F, van Meer G, VanNieuwenhze M, White S, Witztum J, Dennis E (2005). «A comprehensive classification system for lipids». J Lipid Res 46 (5): 839–61. DOI:10.1194/jlr.E400004-JLR200. PMID 15722563.
- ↑ Nomenclature of Lipids. IUPAC-IUB Commission on Biochemical Nomenclature (CBN). Архивировано из первоисточника 22 августа 2011. Проверено 8 марта 2007.
- ↑ Hegardt F (1999). «Mitochondrial 3-hydroxy-3-methylglutaryl-CoA synthase: a control enzyme in ketogenesis». Biochem J 338 (Pt 3): 569–82. DOI:10.1042/0264-6021:3380569. PMID 10051425.
- ↑ Raman R, Raguram S, Venkataraman G, Paulson J, Sasisekharan R (2005). «Glycomics: an integrated systems approach to structure-function relationships of glycans». Nat Methods 2 (11): 817–24. DOI:10.1038/nmeth807. PMID 16278650.
- ↑ Sierra S, Kupfer B, Kaiser R (2005). «Basics of the virology of HIV-1 and its replication». J Clin Virol 34 (4): 233–44. DOI:10.1016/j.jcv.2005.09.004. PMID 16198625.
- ↑ 1 2 Wimmer M, Rose I (1978). «Mechanisms of enzyme-catalyzed group transfer reactions». Annu Rev Biochem 47: 1031–78. DOI:10.1146/annurev.bi.47.070178.005123. PMID 354490.
- ↑ Mitchell P (1979). «The Ninth Sir Hans Krebs Lecture. Compartmentation and communication in living systems. Ligand conduction: a general catalytic principle in chemical, osmotic and chemiosmotic reaction systems». Eur J Biochem 95 (1): 1–20. DOI:10.1111/j.1432-1033.1979.tb12934.x. PMID 378655.
- ↑ 1 2 3 4 Dimroth P, von Ballmoos C, Meier T (March 2006). «Catalytic and mechanical cycles in F-ATP synthases. Fourth in the Cycles Review Series». EMBO Rep 7 (3): 276–82. DOI:10.1038/sj.embor.7400646. PMID 16607397.
- ↑ Stanford School of Medicine Nutrition Courses. — SUMMIT, 2006.
- ↑ Pollak N, Dölle C, Ziegler M (2007). «The power to reduce: pyridine nucleotides—small molecules with a multitude of functions». Biochem J 402 (2): 205–18. DOI:10.1042/BJ20061638. PMID 17295611.
- ↑ 1 2 Heymsfield S, Waki M, Kehayias J, Lichtman S, Dilmanian F, Kamen Y, Wang J, Pierson R (1991). «Chemical and elemental analysis of humans in vivo using improved body composition models». Am J Physiol 261 (2 Pt 1): E190–8. PMID 1872381.
- ↑ Sychrová H (2004). «Yeast as a model organism to study transport and homeostasis of alkali metal cations» (PDF). Physiol Res 53 Suppl 1: S91–8. PMID 15119939.
- ↑ Levitan I (1988). «Modulation of ion channels in neurons and other cells». Annu Rev Neurosci 11: 119–36. DOI:10.1146/annurev.ne.11.030188.001003. PMID 2452594.
- ↑ Dulhunty A (2006). «Excitation-contraction coupling from the 1950s into the new millennium». Clin Exp Pharmacol Physiol 33 (9): 763–72. DOI:10.1111/j.1440-1681.2006.04441.x. PMID 16922804.
- ↑ Mahan D, Shields R (1998). «Macro- and micromineral composition of pigs from birth to 145 kilograms of body weight». J Anim Sci 76 (2): 506–12. PMID 9498359.
- ↑ Husted S, Mikkelsen B, Jensen J, Nielsen N (2004). «Elemental fingerprint analysis of barley (Hordeum vulgare) using inductively coupled plasma mass spectrometry, isotope-ratio mass spectrometry, and multivariate statistics». Anal Bioanal Chem 378 (1): 171–82. DOI:10.1007/s00216-003-2219-0. PMID 14551660.
- ↑ Finney L, O'Halloran T (2003). «Transition metal speciation in the cell: insights from the chemistry of metal ion receptors». Science 300 (5621): 931–6. DOI:10.1126/science.1085049. PMID 12738850.
- ↑ Cousins R, Liuzzi J, Lichten L (2006). «Mammalian zinc transport, trafficking, and signals». J Biol Chem 281 (34): 24085–9. DOI:10.1074/jbc.R600011200. PMID 16793761.
- ↑ Dunn L, Rahmanto Y, Richardson D (2007). «Iron uptake and metabolism in the new millennium». Trends Cell Biol 17 (2): 93–100. DOI:10.1016/j.tcb.2006.12.003. PMID 17194590.
- ↑ Nealson K, Conrad P (1999). «Life: past, present and future». Philos Trans R Soc Lond B Biol Sci 354 (1392): 1923–39. DOI:10.1098/rstb.1999.0532. PMID 10670014.
- ↑ Nelson N, Ben-Shem A (2004). «The complex architecture of oxygenic photosynthesis». Nat Rev Mol Cell Biol 5 (12): 971–82. DOI:10.1038/nrm1525. PMID 15573135.
- ↑ Häse C, Finkelstein R (December 1993). «Bacterial extracellular zinc-containing metalloproteases». Microbiol Rev 57 (4): 823–37. PMID 8302217.
- ↑ Gupta R, Gupta N, Rathi P (2004). «Bacterial lipases: an overview of production, purification and biochemical properties». Appl Microbiol Biotechnol 64 (6): 763–81. DOI:10.1007/s00253-004-1568-8. PMID 14966663.
- ↑ Hoyle T (1997). «The digestive system: linking theory and practice». Br J Nurs 6 (22): 1285–91. PMID 9470654.
- ↑ Souba W, Pacitti A (1992). «How amino acids get into cells: mechanisms, models, menus, and mediators». JPEN J Parenter Enteral Nutr 16 (6): 569–78. DOI:10.1177/0148607192016006569. PMID 1494216.
- ↑ Barrett M, Walmsley A, Gould G (1999). «Structure and function of facilitative sugar transporters». Curr Opin Cell Biol 11 (4): 496–502. DOI:10.1016/S0955-0674(99)80072-6. PMID 10449337.
- ↑ Bell G, Burant C, Takeda J, Gould G (1993). «Structure and function of mammalian facilitative sugar transporters». J Biol Chem 268 (26): 19161–4. PMID 8366068.
- ↑ 1 2 Bouché C, Serdy S, Kahn C, Goldfine A (2004). «The cellular fate of glucose and its relevance in type 2 diabetes». Endocr Rev 25 (5): 807–30. DOI:10.1210/er.2003-0026. PMID 15466941.
- ↑ Sakami W, Harrington H (1963). «Amino acid metabolism». Annu Rev Biochem 32: 355–98. DOI:10.1146/annurev.bi.32.070163.002035. PMID 14144484.
- ↑ Brosnan J (2000). «Glutamate, at the interface between amino acid and carbohydrate metabolism». J Nutr 130 (4S Suppl): 988S–90S. PMID 10736367.
- ↑ Young V, Ajami A (2001). «Glutamine: the emperor or his clothes?». J Nutr 131 (9 Suppl): 2449S–59S; discussion 2486S–7S. PMID 11533293.
- ↑ Hosler J, Ferguson-Miller S, Mills D (2006). «Energy transduction: proton transfer through the respiratory complexes». Annu Rev Biochem 75: 165–87. DOI:10.1146/annurev.biochem.75.062003.101730. PMID 16756489.
- ↑ Schultz B, Chan S (2001). «Structures and proton-pumping strategies of mitochondrial respiratory enzymes». Annu Rev Biophys Biomol Struct 30: 23–65. DOI:10.1146/annurev.biophys.30.1.23. PMID 11340051.
- ↑ Capaldi R, Aggeler R (2002). «Mechanism of the F(1)F(0)-type ATP synthase, a biological rotary motor». Trends Biochem Sci 27 (3): 154–60. DOI:10.1016/S0968-0004(01)02051-5. PMID 11893513.
- ↑ Friedrich B, Schwartz E (1993). «Molecular biology of hydrogen utilization in aerobic chemolithotrophs». Annu Rev Microbiol 47: 351–83. DOI:10.1146/annurev.mi.47.100193.002031. PMID 8257102.
- ↑ Weber K, Achenbach L, Coates J (2006). «Microorganisms pumping iron: anaerobic microbial iron oxidation and reduction». Nat Rev Microbiol 4 (10): 752–64. DOI:10.1038/nrmicro1490. PMID 16980937.
- ↑ Jetten M, Strous M, van de Pas-Schoonen K, Schalk J, van Dongen U, van de Graaf A, Logemann S, Muyzer G, van Loosdrecht M, Kuenen J (1998). «The anaerobic oxidation of ammonium». FEMS Microbiol Rev 22 (5): 421–37. DOI:10.1111/j.1574-6976.1998.tb00379.x. PMID 9990725.
- ↑ Simon J (2002). «Enzymology and bioenergetics of respiratory nitrite ammonification». FEMS Microbiol Rev 26 (3): 285–309. DOI:10.1111/j.1574-6976.2002.tb00616.x. PMID 12165429.
- ↑ Conrad R (1996). «Soil microorganisms as controllers of atmospheric trace gases (H2, CO, CH4, OCS, N2O, and NO)». Microbiol Rev 60 (4): 609–40. PMID 8987358.
- ↑ Barea J, Pozo M, Azcón R, Azcón-Aguilar C (2005). «Microbial co-operation in the rhizosphere». J Exp Bot 56 (417): 1761–78. DOI:10.1093/jxb/eri197. PMID 15911555.
- ↑ van der Meer M, Schouten S, Bateson M, Nübel U, Wieland A, Kühl M, de Leeuw J, Sinninghe Damsté J, Ward D (July 2005). «Diel variations in carbon metabolism by green nonsulfur-like bacteria in alkaline siliceous hot spring microbial mats from Yellowstone National Park». Appl Environ Microbiol 71 (7): 3978–86. DOI:10.1128/AEM.71.7.3978-3986.2005. PMID 16000812.
- ↑ Tichi M, Tabita F (2001). «Interactive control of Rhodobacter capsulatus redox-balancing systems during phototrophic metabolism». J Bacteriol 183 (21): 6344–54. DOI:10.1128/JB.183.21.6344-6354.2001. PMID 11591679.
- ↑ Allen J, Williams J (1998). «Photosynthetic reaction centers». FEBS Lett 438 (1–2): 5–9. DOI:10.1016/S0014-5793(98)01245-9. PMID 9821949.
- ↑ Munekage Y, Hashimoto M, Miyake C, Tomizawa K, Endo T, Tasaka M, Shikanai T (2004). «Cyclic electron flow around photosystem I is essential for photosynthesis». Nature 429 (6991): 579–82. DOI:10.1038/nature02598. PMID 15175756.
- ↑ Miziorko H, Lorimer G (1983). «Ribulose-1,5-bisphosphate carboxylase-oxygenase». Annu Rev Biochem 52: 507–35. DOI:10.1146/annurev.bi.52.070183.002451. PMID 6351728.
- ↑ Dodd A, Borland A, Haslam R, Griffiths H, Maxwell K (2002). «Crassulacean acid metabolism: plastic, fantastic». J Exp Bot 53 (369): 569–80. DOI:10.1093/jexbot/53.369.569. PMID 11886877.
- ↑ Hügler M, Wirsen C, Fuchs G, Taylor C, Sievert S (May 2005). «Evidence for autotrophic CO2 fixation via the reductive tricarboxylic acid cycle by members of the epsilon subdivision of proteobacteria». J Bacteriol 187 (9): 3020–7. DOI:10.1128/JB.187.9.3020-3027.2005. PMID 15838028.
- ↑ Strauss G, Fuchs G (1993). «Enzymes of a novel autotrophic CO2 fixation pathway in the phototrophic bacterium Chloroflexus aurantiacus, the 3-hydroxypropionate cycle». Eur J Biochem 215 (3): 633–43. DOI:10.1111/j.1432-1033.1993.tb18074.x. PMID 8354269.
- ↑ Wood H (1991). «Life with CO or CO2 and H2 as a source of carbon and energy». FASEB J 5 (2): 156–63. PMID 1900793.
- ↑ Shively J, van Keulen G, Meijer W (1998). «Something from almost nothing: carbon dioxide fixation in chemoautotrophs». Annu Rev Microbiol 52: 191–230. DOI:10.1146/annurev.micro.52.1.191. PMID 9891798.
- ↑ Boiteux A, Hess B (1981). «Design of glycolysis». Philos Trans R Soc Lond B Biol Sci 293 (1063): 5–22. DOI:10.1098/rstb.1981.0056. PMID 6115423.
- ↑ Pilkis S, el-Maghrabi M, Claus T (1990). «Fructose-2,6-bisphosphate in control of hepatic gluconeogenesis. From metabolites to molecular genetics». Diabetes Care 13 (6): 582–99. DOI:10.2337/diacare.13.6.582. PMID 2162755.
- ↑ 1 2 Ensign S (2006). «Revisiting the glyoxylate cycle: alternate pathways for microbial acetate assimilation». Mol Microbiol 61 (2): 274–6. DOI:10.1111/j.1365-2958.2006.05247.x. PMID 16856935.
- ↑ Finn P, Dice J (2006). «Proteolytic and lipolytic responses to starvation». Nutrition 22 (7–8): 830–44. DOI:10.1016/j.nut.2006.04.008. PMID 16815497.
- ↑ 1 2 Kornberg H, Krebs H (1957). «Synthesis of cell constituents from C2-units by a modified tricarboxylic acid cycle». Nature 179 (4568): 988–91. DOI:10.1038/179988a0. PMID 13430766.
- ↑ Opdenakker G, Rudd P, Ponting C, Dwek R (1993). «Concepts and principles of glycobiology». FASEB J 7 (14): 1330–7. PMID 8224606.
- ↑ McConville M, Menon A (2000). «Recent developments in the cell biology and biochemistry of glycosylphosphatidylinositol lipids (review)». Mol Membr Biol 17 (1): 1–16. DOI:10.1080/096876800294443. PMID 10824734.
- ↑ Chirala S, Wakil S (2004). «Structure and function of animal fatty acid synthase». Lipids 39 (11): 1045–53. DOI:10.1007/s11745-004-1329-9. PMID 15726818.
- ↑ White S, Zheng J, Zhang Y (2005). «The structural biology of type II fatty acid biosynthesis». Annu Rev Biochem 74: 791–831. DOI:10.1146/annurev.biochem.74.082803.133524. PMID 15952903.
- ↑ Ohlrogge J, Jaworski J (1997). «Regulation of fatty acid synthesis». Annu Rev Plant Physiol Plant Mol Biol 48: 109–136. DOI:10.1146/annurev.arplant.48.1.109. PMID 15012259.
- ↑ Dubey V, Bhalla R, Luthra R (2003). «An overview of the non-mevalonate pathway for terpenoid biosynthesis in plants» (PDF). J Biosci 28 (5): 637–46. DOI:10.1007/BF02703339. PMID 14517367.
- ↑ 1 2 Kuzuyama T, Seto H (2003). «Diversity of the biosynthesis of the isoprene units». Nat Prod Rep 20 (2): 171–83. DOI:10.1039/b109860h. PMID 12735695.
- ↑ Grochowski L, Xu H, White R (May 2006). «Methanocaldococcus jannaschii uses a modified mevalonate pathway for biosynthesis of isopentenyl diphosphate». J Bacteriol 188 (9): 3192–8. DOI:10.1128/JB.188.9.3192-3198.2006. PMID 16621811.
- ↑ Lichtenthaler H (1999). «The 1-Ddeoxy-D-xylulose-5-phosphate pathway of isoprenoid biosynthesis in plants». Annu Rev Plant Physiol Plant Mol Biol 50: 47–65. DOI:10.1146/annurev.arplant.50.1.47. PMID 15012203.
- ↑ 1 2 Schroepfer G (1981). «Sterol biosynthesis». Annu Rev Biochem 50: 585–621. DOI:10.1146/annurev.bi.50.070181.003101. PMID 7023367.
- ↑ Lees N, Skaggs B, Kirsch D, Bard M (1995). «Cloning of the late genes in the ergosterol biosynthetic pathway of Saccharomyces cerevisiae—a review». Lipids 30 (3): 221–6. DOI:10.1007/BF02537824. PMID 7791529.
- ↑ Guyton Arthur C. Textbook of Medical Physiology. — Philadelphia: Elsevier, 2006. — P. 855–6. — ISBN 0-7216-0240-1
- ↑ Ibba M, Söll D (2001). «The renaissance of aminoacyl-tRNA synthesis». EMBO Rep 2 (5): 382–7. PMID 11375928.
- ↑ Lengyel P, Söll D (1969). «Mechanism of protein biosynthesis». Bacteriol Rev 33 (2): 264–301. PMID 4896351.
- ↑ 1 2 Rudolph F (1994). «The biochemistry and physiology of nucleotides». J Nutr 124 (1 Suppl): 124S–127S. PMID 8283301.
- ↑ Zrenner R, Stitt M, Sonnewald U, Boldt R (2006). «Pyrimidine and purine biosynthesis and degradation in plants». Annu Rev Plant Biol 57: 805–36. DOI:10.1146/annurev.arplant.57.032905.105421. PMID 16669783.
- ↑ Stasolla C, Katahira R, Thorpe T, Ashihara H (2003). «Purine and pyrimidine nucleotide metabolism in higher plants». J Plant Physiol 160 (11): 1271–95. DOI:10.1078/0176-1617-01169. PMID 14658380.
- ↑ Smith J (1995). «Enzymes of nucleotide synthesis». Curr Opin Struct Biol 5 (6): 752–7. DOI:10.1016/0959-440X(95)80007-7. PMID 8749362.
- ↑ Testa B, Krämer S (2006). «The biochemistry of drug metabolism—an introduction: part 1. Principles and overview». Chem Biodivers 3 (10): 1053–101. DOI:10.1002/cbdv.200690111. PMID 17193224.
- ↑ Danielson P (2002). «The cytochrome P450 superfamily: biochemistry, evolution and drug metabolism in humans». Curr Drug Metab 3 (6): 561–97. DOI:10.2174/1389200023337054. PMID 12369887.
- ↑ King C, Rios G, Green M, Tephly T (2000). «UDP-glucuronosyltransferases». Curr Drug Metab 1 (2): 143–61. DOI:10.2174/1389200003339171. PMID 11465080.
- ↑ Sheehan D, Meade G, Foley V, Dowd C (November 2001). «Structure, function and evolution of glutathione transferases: implications for classification of non-mammalian members of an ancient enzyme superfamily». Biochem J 360 (Pt 1): 1–16. DOI:10.1042/0264-6021:3600001. PMID 11695986.
- ↑ Galvão T, Mohn W, de Lorenzo V (2005). «Exploring the microbial biodegradation and biotransformation gene pool». Trends Biotechnol 23 (10): 497–506. DOI:10.1016/j.tibtech.2005.08.002. PMID 16125262.
- ↑ Janssen D, Dinkla I, Poelarends G, Terpstra P (2005). «Bacterial degradation of xenobiotic compounds: evolution and distribution of novel enzyme activities». Environ Microbiol 7 (12): 1868–82. DOI:10.1111/j.1462-2920.2005.00966.x. PMID 16309386.
- ↑ Davies K (1995). «Oxidative stress: the paradox of aerobic life». Biochem Soc Symp 61: 1–31. PMID 8660387.
- ↑ Tu B, Weissman J (2004). «Oxidative protein folding in eukaryotes: mechanisms and consequences». J Cell Biol 164 (3): 341–6. DOI:10.1083/jcb.200311055. PMID 14757749.
- ↑ Sies H (1997). «Oxidative stress: oxidants and antioxidants» (PDF). Exp Physiol 82 (2): 291–5. PMID 9129943.
- ↑ Vertuani S, Angusti A, Manfredini S (2004). «The antioxidants and pro-antioxidants network: an overview». Curr Pharm Des 10 (14): 1677–94. DOI:10.2174/1381612043384655. PMID 15134565.
- ↑ von Stockar U, Liu J (1999). «Does microbial life always feed on negative entropy? Thermodynamic analysis of microbial growth». Biochim Biophys Acta 1412 (3): 191–211. DOI:10.1016/S0005-2728(99)00065-1. PMID 10482783.
- ↑ Demirel Y, Sandler S (2002). «Thermodynamics and bioenergetics». Biophys Chem 97 (2–3): 87–111. DOI:10.1016/S0301-4622(02)00069-8. PMID 12050002.
- ↑ Albert R (2005). «Scale-free networks in cell biology». J Cell Sci 118 (Pt 21): 4947–57. DOI:10.1242/jcs.02714. PMID 16254242.
- ↑ Brand M (1997). «Regulation analysis of energy metabolism». J Exp Biol 200 (Pt 2): 193–202. PMID 9050227.
- ↑ Soyer O, Salathé M, Bonhoeffer S (2006). «Signal transduction networks: topology, response and biochemical processes». J Theor Biol 238 (2): 416–25. DOI:10.1016/j.jtbi.2005.05.030. PMID 16045939.
- ↑ Westerhoff H, Groen A, Wanders R (1984). «Modern theories of metabolic control and their applications (review)». Biosci Rep 4 (1): 1–22. DOI:10.1007/BF01120819. PMID 6365197.
- ↑ Salter M, Knowles R, Pogson C (1994). «Metabolic control». Essays Biochem 28: 1–12. PMID 7925313.
- ↑ Fell D, Thomas S (1995). «Physiological control of metabolic flux: the requirement for multisite modulation». Biochem J 311 (Pt 1): 35–9. PMID 7575476.
- ↑ Hendrickson W (2005). «Transduction of biochemical signals across cell membranes». Q Rev Biophys 38 (4): 321–30. DOI:10.1017/S0033583506004136. PMID 16600054.
- ↑ Cohen P (2000). «The regulation of protein function by multisite phosphorylation—a 25 year update». Trends Biochem Sci 25 (12): 596–601. DOI:10.1016/S0968-0004(00)01712-6. PMID 11116185.
- ↑ Lienhard G, Slot J, James D, Mueckler M (1992). «How cells absorb glucose». Sci Am 266 (1): 86–91. DOI:10.1038/scientificamerican0192-86. PMID 1734513.
- ↑ Roach P (2002). «Glycogen and its metabolism». Curr Mol Med 2 (2): 101–20. DOI:10.2174/1566524024605761. PMID 11949930.
- ↑ Newgard C, Brady M, O'Doherty R, Saltiel A (2000). «Organizing glucose disposal: emerging roles of the glycogen targeting subunits of protein phosphatase-1» (PDF). Diabetes 49 (12): 1967–77. DOI:10.2337/diabetes.49.12.1967. PMID 11117996.
- ↑ Romano A, Conway T (1996). «Evolution of carbohydrate metabolic pathways». Res Microbiol 147 (6–7): 448–55. DOI:10.1016/0923-2508(96)83998-2. PMID 9084754.
- ↑ Koch A (1998). «How did bacteria come to be?». Adv Microb Physiol 40: 353–99. DOI:10.1016/S0065-2911(08)60135-6. PMID 9889982.
- ↑ Ouzounis C, Kyrpides N (1996). «The emergence of major cellular processes in evolution». FEBS Lett 390 (2): 119–23. DOI:10.1016/0014-5793(96)00631-X. PMID 8706840.
- ↑ Caetano-Anolles G, Kim HS, Mittenthal JE (2007). «The origin of modern metabolic networks inferred from phylogenomic analysis of protein architecture». Proc Natl Acad Sci USA 104 (22): 9358–63. DOI:10.1073/pnas.0701214104. PMID 17517598.
- ↑ Schmidt S, Sunyaev S, Bork P, Dandekar T (2003). «Metabolites: a helping hand for pathway evolution?». Trends Biochem Sci 28 (6): 336–41. DOI:10.1016/S0968-0004(03)00114-2. PMID 12826406.
- ↑ Light S, Kraulis P (2004). «Network analysis of metabolic enzyme evolution in Escherichia coli». BMC Bioinformatics 5: 15. DOI:10.1186/1471-2105-5-15. PMID 15113413. Alves R, Chaleil R, Sternberg M (2002). «Evolution of enzymes in metabolism: a network perspective». J Mol Biol 320 (4): 751–70. DOI:10.1016/S0022-2836(02)00546-6. PMID 12095253.
- ↑ Kim HS, Mittenthal JE, Caetano-Anolles G (2006). «MANET: tracing evolution of protein architecture in metabolic networks». BMC Bioinformatics 19 (7): 351. DOI:10.1186/1471-2105-7-351. PMID 16854231.
- ↑ Teichmann SA, Rison SC, Thornton JM, Riley M, Gough J, Chothia C (2001). «Small-molecule metabolsim: an enzyme mosaic». Trends Biotechnol 19 (12): 482–6. DOI:10.1016/S0167-7799(01)01813-3. PMID 11711174.
- ↑ Spirin V, Gelfand M, Mironov A, Mirny L (June 2006). «A metabolic network in the evolutionary context: multiscale structure and modularity». Proc Natl Acad Sci USA 103 (23): 8774–9. DOI:10.1073/pnas.0510258103. PMID 16731630.
- ↑ Lawrence J (2005). «Common themes in the genome strategies of pathogens». Curr Opin Genet Dev 15 (6): 584–8. DOI:10.1016/j.gde.2005.09.007. PMID 16188434. Wernegreen J (2005). «For better or worse: genomic consequences of intracellular mutualism and parasitism». Curr Opin Genet Dev 15 (6): 572–83. DOI:10.1016/j.gde.2005.09.013. PMID 16230003.
- ↑ Pál C, Papp B, Lercher M, Csermely P, Oliver S, Hurst L (2006). «Chance and necessity in the evolution of minimal metabolic networks». Nature 440 (7084): 667–70. DOI:10.1038/nature04568. PMID 16572170.
- ↑ Rennie M (1999). «An introduction to the use of tracers in nutrition and metabolism». Proc Nutr Soc 58 (4): 935–44. DOI:10.1017/S002966519900124X. PMID 10817161.
- ↑ Phair R (1997). «Development of kinetic models in the nonlinear world of molecular cell biology». Metabolism 46 (12): 1489–95. DOI:10.1016/S0026-0495(97)90154-2. PMID 9439549.
- ↑ Sterck L, Rombauts S, Vandepoele K, Rouzé P, Van de Peer Y (2007). «How many genes are there in plants (... and why are they there)?». Curr Opin Plant Biol 10 (2): 199–203. DOI:10.1016/j.pbi.2007.01.004. PMID 17289424.
- ↑ Borodina I, Nielsen J (2005). «From genomes to in silico cells via metabolic networks». Curr Opin Biotechnol 16 (3): 350–5. DOI:10.1016/j.copbio.2005.04.008. PMID 15961036.
- ↑ Gianchandani E, Brautigan D, Papin J (2006). «Systems analyses characterize integrated functions of biochemical networks». Trends Biochem Sci 31 (5): 284–91. DOI:10.1016/j.tibs.2006.03.007. PMID 16616498.
- ↑ Duarte NC, Becker SA, Jamshidi N, et al. (February 2007). «Global reconstruction of the human metabolic network based on genomic and bibliomic data». Proc. Natl. Acad. Sci. U.S.A. 104 (6): 1777–82. DOI:10.1073/pnas.0610772104. PMID 17267599.
- ↑ Goh KI, Cusick ME, Valle D, Childs B, Vidal M, Barabási AL (May 2007). «The human disease network». Proc. Natl. Acad. Sci. U.S.A. 104 (21): 8685–90. DOI:10.1073/pnas.0701361104. PMID 17502601.
- ↑ Lee DS, Park J, Kay KA, Christakis NA, Oltvai ZN, Barabási AL (July 2008). «The implications of human metabolic network topology for disease comorbidity». Proc. Natl. Acad. Sci. U.S.A. 105 (29): 9880–9885. DOI:10.1073/pnas.0802208105. PMID 18599447.
- ↑ Csete M, Doyle J (2004). «Bow ties, metabolism and disease». Trends Biotechnol. 22 (9): 446–50. DOI:10.1016/j.tibtech.2004.07.007. PMID 5249808.
- ↑ Ma HW, Zeng AP (2003). «The connectivity structure, giant strong component and centrality of metabolic networks». Bioinformatics 19 (11): 1423–30. DOI:10.1093/bioinformatics/btg177. PMID 12874056.
- ↑ Zhao J, Yu H, Luo JH, Cao ZW, Li YX (2006). «Hierarchical modularity of nested bow-ties in metabolic networks». BMC Bioinformatics 7: 386. DOI:10.1186/1471-2105-7-386. PMID 16916470.
- ↑ Thykaer J, Nielsen J (2003). «Metabolic engineering of beta-lactam production». Metab Eng 5 (1): 56–69. DOI:10.1016/S1096-7176(03)00003-X. PMID 12749845. González-Pajuelo M, Meynial-Salles I, Mendes F, Andrade J, Vasconcelos I, Soucaille P (2005). «Metabolic engineering of Clostridium acetobutylicum for the industrial production of 1,3-propanediol from glycerol». Metab Eng 7 (5–6): 329–36. DOI:10.1016/j.ymben.2005.06.001. PMID 16095939. Krämer M, Bongaerts J, Bovenberg R, Kremer S, Müller U, Orf S, Wubbolts M, Raeven L (2003). «Metabolic engineering for microbial production of shikimic acid». Metab Eng 5 (4): 277–83. DOI:10.1016/j.ymben.2003.09.001. PMID 14642355.
- ↑ Koffas M, Roberge C, Lee K, Stephanopoulos G (1999). «Metabolic engineering». Annu Rev Biomed Eng 1: 535–57. DOI:10.1146/annurev.bioeng.1.1.535. PMID 11701499.
- ↑ Dr. Abu Shadi Al-Roubi (1982), «Ibn Al-Nafis as a philosopher», Symposium on Ibn al Nafis, Second International Conference on Islamic Medicine: Islamic Medical Organization, Kuwait (cf. Ibnul-Nafees As a Philosopher, Encyclopedia of Islamic World [1]).
- ↑ Eknoyan G (1999). «Santorio Sanctorius (1561–1636) - founding father of metabolic balance studies». Am J Nephrol 19 (2): 226–33. DOI:10.1159/000013455. PMID 10213823.
- ↑ Williams, H. S. (1904) A History of Science: in Five Volumes. Volume IV: Modern Development of the Chemical and Biological Sciences Harper and Brothers (New York) Retrieved on 2007-03-26
- ↑ Dubos J. (1951). «Louis Pasteur: Free Lance of Science, Gollancz. Quoted in Manchester K. L. (1995) Louis Pasteur (1822–1895)—chance and the prepared mind». Trends Biotechnol 13 (12): 511–515. DOI:10.1016/S0167-7799(00)89014-9. PMID 8595136.
- ↑ Kinne-Saffran E, Kinne R (1999). «Vitalism and synthesis of urea. From Friedrich Wöhler to Hans A. Krebs». Am J Nephrol 19 (2): 290–4. DOI:10.1159/000013463. PMID 10213830.
- ↑ Eduard Buchner’s 1907 Nobel lecture at http://nobelprize.org Accessed 2007-03-20
- ↑ Kornberg H (2000). «Krebs and his trinity of cycles». Nat Rev Mol Cell Biol 1 (3): 225–8. DOI:10.1038/35043073. PMID 11252898.
- ↑ Krebs HA, Henseleit K (1932). «Untersuchungen über die Harnstoffbildung im tierkorper». Z. Physiol. Chem. 210: 33–66.Krebs H, Johnson W (April 1937). «Metabolism of ketonic acids in animal tissues». Biochem J 31 (4): 645–60. PMID 16746382.
Ссылки
- Обмен веществ (рус.)
- Метаболизм (англ.)
- Микробный метаболизм Школьный уровень (англ.).
Категории:- Физиология
- Метаболизм








Wikimedia Foundation. 2010.