- Гидра (род)
-
Гидра (род) 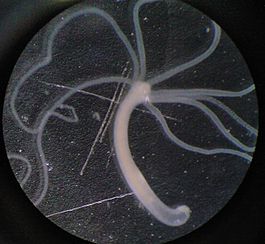
Гидра (Hydra)Научная классификация промежуточные рангиДомен: Эукариоты Царство: Животные Подцарство: Эуметазои Тип: Стрекающие Подтип: Medusozoa Класс: Гидроидные Подкласс: Hydroidolina Отряд: Anthoathecata Семейство: Hydridae Род: Гидры Международное научное название 
Систематика
на Викивидах
Изображения
на ВикискладеITIS 50845 NCBI 6083 EOL 99065 Гидра (лат. Hydra) — род пресноводных сидячих кишечнополостных.
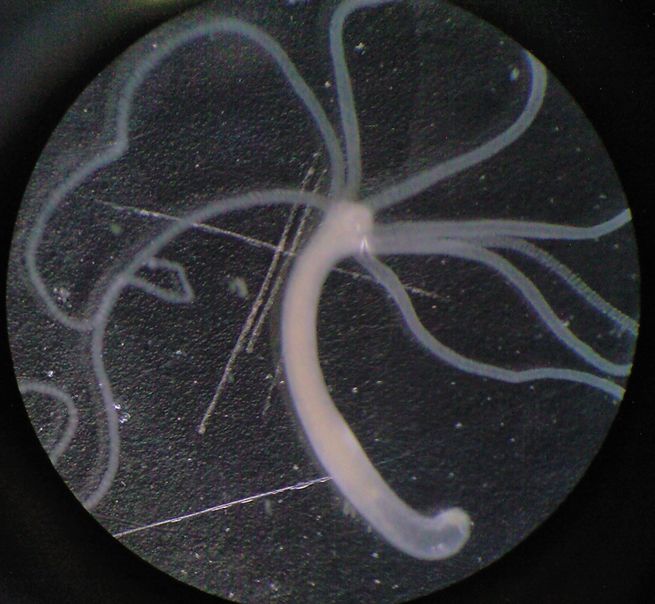
Гидра обитает в стоячих водоёмах и реках с медленным течением. Она прикрепляется к водным растениям или грунту. Длина тела гидры составляет 1—20 мм. Это одиночный малоподвижный полип.План строения
Тело гидры цилиндрической формы, на переднем конце тела на околоротовом конусе расположен рот, окружённый венчиком из 5—12 щупалец. У некоторых видов тело разделено на туловище и стебелёк. На заднем конце тела (стебелька) расположена подошва, с её помощью гидра передвигается и прикрепляется. Гидра обладает радиальной (одноосно-гетеропольной) симметрией. Ось симметрии соединяет два полюса — оральный, на котором находится рот, и аборальный, на котором находится подошва. Через ось симметрии можно провести несколько плоскостей симметрии, разделяющих тело на две зеркально симметричных половины.
Тело гидры — мешок со стенкой из двух слоёв клеток (эктодермы и энтодермы), между которыми находится тонкий слой межклеточного вещества (мезоглея). Полость тела гидры — гастральная полость — образует выросты, заходящие внутрь щупалец. Хотя обычно считают, что у гидры есть только одно ведущее в гастральную полость отверстие (ротовое), на самом деле на подошве гидры имеется узкая аборальная пора. Через неё может выделяться жидкость из кишечной полости, а также пузырёк газа. При этом гидра вместе с пузырьком открепляется от субстрата и всплывает, удерживаясь вниз головой в толще воды. Таким способом она может расселяться по водоёму. Что касается ротового отверстия, то у не питающейся гидры оно фактически отсутствует — клетки эктодермы ротового конуса смыкаются и образуют плотные контакты, такие же, как и на других участках тела[1]. Поэтому при питании гидре каждый раз приходится «прорывать» рот заново.
Клеточный состав тела
Эпителиально-мускульные клетки
Эпителиально-мускульные клетки эктодермы и энтодермы образуют основную массу тела гидры. У гидры около 20 000 эпителиально-мускульных клеток.
Клетки эктодермы имеют цилиндрическую форму эпителиальных частей и формируют однослойный покровный эпителий. К мезоглее прилегают сократимые отростки данных клеток, образующие продольную мускулатуру гидры.
Эпителиально-мускульные клетки энтодермы направлены эпителиальными частями в полость кишки и несут по 2—5 жгутиков, которые перемешивают пищу. Эти клетки могут образовывать ложноножки, с помощью которых захватывают частицы пищи. В клетках формируются пищеварительные вакуоли.
Эпителиально-мускульные клетки эктодермы и энтодермы представляют собой две независимые клеточные линии. В верхней трети туловища гидры они делятся митотически, а их потомки постепенно смещаются либо в сторону гипостома и щупалец, либо в сторону подошвы. По мере перемещения происходит дифференцировка клеток: так, клетки эктодермы на щупальцах дают клетки стрекательных батарей, а на подошве — железистые клетки, выделяющие слизь.
Железистые клетки энтодермы
Железистые клетки энтодермы выделяют в полость кишки пищеварительные ферменты, которые расщепляют пищу. Эти клетки образуются из интерстициальных клеток. У гидры около 5000 железистых клеток.
Интерстициальные клетки
Между эпителиально-мускульными клетками находятся группы мелких, округлых клеток, называемых промежуточными, или интерстициальными (i-клетки). У гидры их около 15 000. Это недифференцированные клетки. Они могут превращаться в остальные типы клеток тела гидры, кроме эпителиально-мускульных. Промежуточные клетки обладают всеми свойствами мультипотентных стволовых клеток. Доказано, что каждая промежуточная клетка потенциально способна дать как половые, так и соматические клетки. Стволовые промежуточные клетки не мигрируют, однако их дифференцирующиеся клетки-потомки способны к быстрым миграциям.
Нервные клетки и нервная система
Нервные клетки образуют в эктодерме примитивную диффузную нервную систему — рассеянное нервное сплетение (диффузный плексус). В энтодерме есть отдельные нервные клетки. Всего у гидры около 5000 нейронов. У гидры имеются сгущения диффузного плексуса на подошве, вокруг рта и на щупальцах. По новым данным, у гидры имеется околоротовое нервное кольцо, сходное с нервным кольцом, расположенным на крае зонтика у гидромедуз.
У гидры нет четкого деления на чувствительные, вставочные и моторные нейроны. Одна и та же клетка может воспринимать раздражение и передавать сигнал эпителиально-мускульным клеткам. Тем не менее, есть два основных типа нервных клеток — чувствительные и ганглиозные. Тела чувствительных клеток расположены поперек эпителиального пласта, они имеют неподвижный жгутик, окруженный воротничком из микроворсинок, который торчит во внешнюю среду и способен воспринимать раздражение. Ганглиозные клетки расположены в основании эпителиально-мускульных, их отростки не выходят во внешнюю среду. По морфологии большинство нейронов гидры — биполярные или мультиполярные.
В нервной системе гидры присутствуют как электрические, так и химические синапсы. Из нейромедиаторов у гидры обнаружены дофамин, серотонин, норадреналин, гамма-аминомасляная кислота, глютамат, глицин и многие нейропептиды (вазопрессин, вещество Р и др.).
Гидра — наиболее примитивное животное, в нервных клетках которого обнаружены чувствительные к свету белки опсины. Анализ гена опсина гидры позволяет предположить, что опсины гидры и человека имеют общее происхождение[2].
Стрекательные клетки
Стрекательные клетки образуются из промежуточных только в области туловища. Сначала промежуточная клетка делится 3–5 раз, образуя кластер (гнездо) из предшественников стрекательных клеток (книдобластов), соединенных цитоплазматическими мостиками. Затем начинается дифференцировка, в ходе которой мостики исчезают. Дифференцирующиеся книдоциты мигрируют в щупальца. Стрекательные клетки наиболее многочисленные из всех клеточных типов, их у гидры около 55 000.
Стрекательная клетка имеет стрекательную капсулу, заполненную ядовитым веществом. Внутрь капсулы ввёрнута стрекательная нить. На поверхности клетки находится чувствительный волосок, при его раздражении нить выбрасывается и поражает жертву. После выстреливания нити клетки погибают, а из промежуточных клеток образуются новые.
У гидры есть четыре типа стрекательных клеток — стенотелы (пенетранты), десмонемы (вольвенты), голотрихи изоризы (большие глютинанты) и атрихи изоризы (малые глютинанты). При охоте первыми выстреливают вольвенты. Их спиральные стрекательные нити опутывают выросты тела жертвы и обеспечивают её удержание. Под действием рывков жертвы и вызванной ими вибрации срабатывают имеющие более высокий порог раздражения пенетранты. Шипы, имеющиеся у основания их стрекательных нитей, заякориваются в теле добычи, а через полую стрекательную нить в её тело вводится яд.
Большое количество стрекательных клеток находится на щупальцах, где они образуют стрекательные батареи. Обычно в состав батареи входит одна крупная эпителиально-мускульная клетка, в которую погружены стрекательные клетки. В центре батареи находится крупная пенетранта, вокруг неё — более мелкие вольвенты и глютинанты. Книдоциты соединены десмосомами с мускульными волокнами эпителиально-мускульной клетки. Большие глютинанты (их стрекательная нить имеет шипы, но не имеет, как и у вольвент, отверстия на вершине), видимо, в основном используются для защиты. Малые глютинанты используются только при передвижении гидры для прочного прикрепления щупальцами к субстрату. Их выстреливание блокируется экстрактами из тканей жертв гидры.
Выстреливание пенетрант гидры было изучено с помощью сверхвысокоскоростной киносъемки. Оказалось, что весь процесс выстреливания занимает около 3 мс. В его начальной фазе (до выворачивания шипов) скорость его достигает 2 м/c, а ускорение составляет около 40 000 g (данные 1984 года[3][4]); видимо, это один из самых быстрых клеточных процессов, известных в природе. Первым видимым изменением (менее чем через 10 мкс после стимуляции) было увеличение объёма стрекательной капсулы примерно на 10 %, затем объём снижается почти до 50 % от исходного. В дальнейшем выяснилось, что и скорость, и ускорение при выстреливании нематоцист были сильно недооценены; по данным 2006 года[5], на ранней фазе выстреливания (выбрасывание шипов) скорость этого процесса составляет 9-18 м/с, а ускорение составляет от 1 000 000 до 5 400 000 g. Это позволяет нематоцисте массой около 1 нг развивать на кончиках шипов (диаметр которых составляет около 15 нм) давление порядка 7 гПа, что сравнимо с давлением пули на мишень и позволяет пробивать достаточно толстую кутикулу жертв.
Половые клетки и гаметогенез
Как и всем животным, гидрам свойственна оогамия. Большинство гидр раздельнополы, но встречаются гермафродитные линии гидр. И яйцеклетки, и сперматозоиды образуются из I-клеток . Считается, что это особые субпопуляции i-клеток, которые можно отличить по клеточным маркерам и которые в небольшом количестве присутствуют у гидр и в период бесполого размножения.
При оогенезе ооциты фагоцитируют целые оогонии, а затем несколько ооцитов сливаются, после чего ядро одного из них превращается в ядро яйцеклетки, а остальные ядра дегенерируют. Эти процессы обеспечивают быстрый рост яйцеклетки.
Как недавно показано, при сперматогенезе имеет место программированная клеточная смерть части клеток-предшественников сперматозоидов и их фагоцитирование окружающими клетками эктодермы[6]
Дыхание и выделение
Дыхание и выделение продуктов обмена происходит через всю поверхность тела животного. Вероятно, в выделении некоторую роль играют вакуоли, которые есть в клетках гидры. Главная функция вакуолей, вероятно, осморегуляторная; они выводят излишки воды, которые постоянно поступают в клетки гидры путем осмоса.
Раздражимость и рефлексы
Наличие нервной системы позволяет гидре осуществлять простые рефлексы. Гидра реагирует на механическое раздражение, температуру, освещённость[2], наличие в воде химических веществ и на ряд других факторов внешней среды.
Питание и пищеварение
Гидра питается мелкими беспозвоночными — дафниями и другими ветвистоусыми, циклопами, а также олигохетами-наидидами. Есть данные о потреблении гидрами коловраток и церкарий трематод. Добыча захватывается щупальцами с помощью стрекательных клеток, яд которых быстро парализует мелких жертв. Координированными движениями щупалец добыча подносится ко рту, а затем с помощью сокращений тела гидра «надевается» на жертву. Пищеварение начинается в кишечной полости (полостное пищеварение), заканчивается внутри пищеварительных вакуолей эпителиально-мускульных клеток энтодермы (внутриклеточное пищеварение). Непереваренные остатки пищи выбрасываются через рот.
Так как у гидры нет транспортной системы, а мезоглея (слой межклеточного вещества между экто-и энтодермой) достаточно плотная, возникает проблема транспорта питательных веществ к клеткам эктодермы. Эта проблема решается за счет образования выростов клеток обоих слоёв, которые пересекают мезоглею и соединяются через щелевые контакты. Через них могут проходить мелкие органические молекулы (моносахариды, аминокислоты), что обеспечивает питание клеток эктодермы.Размножение и развитие
При благоприятных условиях гидра размножается бесполым путём. На теле животного (обычно в нижней трети туловища) образуется почка, она растет, затем формируются щупальца и прорывается рот. Молодая гидра отпочковывается от материнского организма (при этом материнский и дочерний полипы прикрепляются щупальцами к субстрату и тянут в разные стороны) и ведет самостоятельный образ жизни. Осенью гидра переходит к половому размножению. На теле, в эктодерме закладываются гонады — половые железы, а в них из промежуточных клеток развиваются половые клетки. При образовании гонад гидр формируется медузоидный узелок. Это позволяет предполагать, что гонады гидры — сильно упрощенные споросаки, последний этап в ряду преобразования утраченного медузоидного поколения в орган. Большинство видов гидр раздельнополы, реже встречается гермафродитизм. Яйцеклетки гидр быстро растут, фагоцитируя окружающие клетки. Зрелые яйцеклетки достигают диаметра 0,5—1 мм. Оплодотворение происходит в теле гидры: через специальное отверстие в гонаде сперматозоид проникает к яйцеклетке и сливается с ней. Зигота претерпевает полное равномерное дробление, в результате которого образуется целобластула. Затем в результате смешанной деламинации (сочетание иммиграции и деламинации) осуществляется гаструляция. Вокруг зародыша формируется плотная защитная оболочка (эмбриотека) с выростами-шипиками. На стадии гаструлы зародыши впадают в анабиоз. Взрослые гидры погибают, а зародыши опускаются на дно и зимуют. Весной продолжается развитие, в паренхиме энтодермы путем расхождения клеток образуется кишечная полость, затем формируются зачатки щупалец, и из-под оболочки выходит молодая гидра. Таким образом, в отличие от большинства морских гидроидных, у гидры отсутствуют свободноплавающие личинки, развитие у неё прямое.
Рост и регенерация
Миграция и обновление клеток
В норме у взрослой гидры клетки всех трех клеточных линий интенсивно делятся в средней части тела и мигрируют к подошве, гипостому и кончикам щупалец. Там происходит гибель и слущивание клеток. Таким образом, все клетки тела гидры постоянно обновляются. При нормальном питании «избыток» делящихся клеток перемещается в почки, которые обычно образуются в нижней трети туловища.
Регенерационная способность
Гидра обладает очень высокой способностью к регенерации. При разрезании поперек на несколько частей каждая часть восстанавливает «голову» и «ногу», сохраняя исходную полярность — рот и щупальца развиваются на той стороне, которая была ближе к оральному концу тела, а стебелек и подошва — на аборальной стороне фрагмента. Целый организм может восстанавливаться из отдельных небольших кусочков тела (менее 1/200 объёма), из кусочков щупалец, а также из взвеси клеток. При этом сам процесс регенерации не сопровождается усилением клеточных делений и представляет собой типичный пример морфаллаксиса.
Гидра может регенерировать из взвеси клеток, полученных путем мацерации (например, при протирании гидры через мельничный газ). В экспериментах показано, что для восстановления головного конца достаточно образования агрегата из примерно 300 эпителиально-мускульных клеток. Показано, что регенерация нормального организма возможна из клеток одного слоя (только эктодермы или только энтодермы).
Опыты по изучению регенерации и модели регенерации
Уже ранние опыты Трамбле показали, что при регенерации сохраняется полярность фрагмента. Если разрезать тело гидры поперек на несколько цилиндрических фрагментов, то на каждом из них ближе к бывшему оральному концу регенерируют гипостом и щупальца (в экспериментальной эмбриологии гидры закрепился термин «голова» для обозначения орального конца тела), а ближе к бывшему аборальному полюсу — подошва («нога»). При этом у тех фрагментов, которые располагались ближе к «голове», быстрее регенерирует «голова», а у располагавшихся ближе к «ноге» — «нога».
Позднее опыты по изучению регенерации были усовершенствованы в результате применения методики сращивания фрагментов разных особей. Если вырезать из боковой стороны туловища гидры фрагмент и срастить его с телом другой гидры, то возможны три исхода опыта: 1) фрагмент полностью сливается с телом реципиента; 2) фрагмент образует выступ, на конце которого развивается «голова» (то есть превращается в почку); 3) фрагмент образует выступ, на конце которого образуется «нога». Выяснилось, что процент образования «голов» тем выше, чем ближе к «голове» донора взят фрагмент для пересадки и чем дальше от «головы» реципиента он помещен. Эти и аналогичные опыты привели к постулированию существования четырёх веществ-морфогенов, регулирующих регенерацию — активатора и ингибитора «головы» и активатора и ингибитора «ноги». Эти вещества, согласно данной модели регенерации, образуют концентрационные градиенты: в районе «головы» у нормального полипа максимальна концентрация как активатора, так и ингибитора головы, а в районе «ноги» — максимальна концентрация и активатора, и ингибитора ноги.
Эти вещества действительно были обнаружены. Активатор головы — пептид из 11 аминокислот (pGlu-Pro-Pro-Gly-Gly-Ser-Lys-Val-Ile-Leu-Phe), активный в пикомолярной концентрации (первоначально 20 мкг активатора были получены из 200 кг морских актиний). У человека он присутствует в гипоталамусе и кишечнике и в той же концентрации обладает нейротрофическим действием. У гидры и млекопитающих этот пептид обладает также митогенным действием и влияет на клеточную дифференцировку.
Активатор ноги — тоже пептид с молекулярной массой, близкой к 1000 Да. Ингибиторы головы и ноги — низкомолекулярные гидрофильные вещества небелковой природы. В норме все четыре вещества выделяются нервными клетками гидры. Активатор головы имеет большее время полужизни (около 4 ч), чем ингибитор (30 мин) и медленнее диффундирует, так как связан с белком-носителем. Ингибитор головы в очень низкой концентрации подавляет выделение активатора, а в 20 раз большей концентрации — своё собственное выделение. Ингибитор ноги также ингибирует выделение активатора ноги.
Молекулярные механизмы регенерации

Этот раздел статьи ещё не написан. Согласно замыслу одного из участников Википедии, на этом месте должен располагаться специальный раздел.
Вы можете помочь проекту, написав этот раздел.Получение «безнервных» гидр
При регенерации, как и при росте и бесполом размножении, эпителиально-мускульные клетки делятся самостоятельно, причем клетки эктодермы и энтодермы — две независимые клеточные линии. Остальные типы клеток (нервные, стрекательные и железистые) развиваются из промежуточных. Убив делящиеся промежуточные клетки высокой дозой радиации или колхицином, можно получить «безнервных», или эпителиальных гидр — они продолжают расти и почковаться, но отделяющиеся почки лишены нервных и стрекательных клеток. Культуру таких гидр удается поддерживать в лаборатории с помощью «насильственного» кормления.
Известны также мутантные линии «безнервных» гидр, у которых нет промежуточных клеток и у которых промежуточные клетки могут давать только сперматозоиды, но не соматические клетки, а также мутантные линии, у которых промежуточные клетки погибают при повышенной температуре.
«Безнервные» гидры сохраняют способность к регенерации.
Продолжительность жизни
Ещё в конце XIX века была выдвинута гипотеза о теоретическом бессмертии гидры, которую пытались научно доказать или опровергнуть на протяжении всего XX века. В 1997 году гипотеза была доказана экспериментальным путём Даниэлем Мартинесом[7]. Эксперимент продолжался порядка четырёх лет и показал отсутствие смертности среди трёх групп гидр вследствие старения. Считается, что «бессмертность» гидр напрямую связана с их высокой регенерационной способностью.
Перед наступлением зимы, после перехода к половому размножению и созреванию покоящихся стадий, гидры в водоёмах средней полосы погибают. Видимо, это происходит не из-за нехватки пищи или непосредственного воздействия иных неблагоприятных факторов. Это говорит о наличии у гидр механизмов старения[8].
Местные виды
В водоёмах России и Украины наиболее часто встречаются следующие виды гидр (в настоящее время многие зоологи выделяют кроме рода Hydra ещё 2 рода — Pelmatohydra и Chlorohydra):
- гидра длинностебельчатая (Hydra (Pelmatohydra) oligactis) — крупная, с пучком очень длинных нитевидных щупалец, в 2—5 раз превышающих длину её тела;
- гидра обыкновенная (Hydra vulgaris) — щупальца приблизительно вдвое длиннее тела, а само тело, как и у предыдущего вида сужается ближе к подошве;
- гидра тонкая (Hydra attennata) — тело этой гидры имеет вид тонкой трубочки равномерной толщины, а щупальца только слегка длиннее тела;
- гидра зелёная (Hydra (Chlorohydra) viridissima) с короткими, но многочисленными щупальцами, травянистого зелёного цвета.
 Зелёные гидры
Зелёные гидры
Симбионты
У так называемых «зеленых» гидр Hydra (Chlorohydra) viridissima в клетках энтодермы живут эндосимбиотические водоросли рода Chlorella — зоохлореллы. На свету такие гидры могут длительное время (более четырёх месяцев) обходиться без пищи, в то время как искусственно лишенные симбионтов гидры без кормления погибают через два месяца. Зоохлореллы проникают в яйцеклетки и передаются потомству трансовариально. Другие виды гидр в лабораторных условиях иногда удается заразить зоохлореллами, однако устойчивого симбиоза при этом не возникает.
Именно с наблюдений за зелеными гидрами начал свои исследования А. Трамбле.
Хищники и паразиты
На гидр могут нападать мальки рыб, для которых ожоги стрекательных клеток, видимо, довольно чувствительны: схватив гидру, малёк обычно выплёвывает её и отказывается от дальнейших попыток съесть.
На поверхности тела гидр в качестве паразитов или комменсалов часто обитают Kerona polyporum, триходина и другие инфузории.
К питанию тканями гидр приспособлен ветвистоусый рачок из семейства хидорид Anchistropus emarginatus.
Тканями гидр могут также питаться турбеллярии микростомулы, которые способны использовать непереваренные молодые стрекательные клетки гидр в качестве защитных клеток — клептокнид.
История открытия и изучения
Видимо, впервые описал гидру Антонио ван Левенгук. Подробно изучил питание, движение и бесполое размножение, а также регенерацию гидры Авраам Трамбле, который описал результаты своих опытов и наблюдений в книге «Мемуары к истории одного рода пресноводных полипов с руками в форме рогов» (первое издание вышло на французском языке в 1744 г.). Открытие Трамбле приобрело громкую славу, его опыты обсуждались в светских салонах и при французском королевском дворе. Эти опыты опровергли господствовавшее тогда убеждение, что отсутствие бесполого размножения и развитой регенерации у животных — одно из важнейших их отличий от растений. Считается, что изучении регенерации гидры (опыты А. Трамбле) положили начало экспериментальной зоологии. Научное название роду в соответствии с правилами зоологической номенклатуры присвоил Карл Линней.
Гидра как модельный объект
 Трансгенная гидра Hydra vulgaris линии AEP с энтодермальными клетками, в которых экспрессируется зелёный флуоресцентный белок
Трансгенная гидра Hydra vulgaris линии AEP с энтодермальными клетками, в которых экспрессируется зелёный флуоресцентный белокВ последние десятилетия гидра используется как модельный объект для изучения регенерации и процессов морфогенеза. Геном гидры (североамериканский вид Hydra magnipapillata) частично расшифрован. Имеются коллекции мутантных линий гидры в Японии и Германии. Разработана методика получения трансгенных гидр.
Источники
- Степаньянц С. Д., Кузнецова В. Г., Анохин Б. А. Гидра: от Абраама Трамбле до наших дней. — Москва-Санкт-Петербург: Товарищество научных изданий КМК, 2003. — (Разнообразие животных).
- Трамбле А. Мемуары к истории одного рода преноводных полипов с руками в форме рогов. — М.-Л.: Государственное издательство биологической и медицинской литературы, 1937.
- Иофф Н. А. Курс эмбриологии беспозвоночных. — М.: Высшая школа, 1962.
- G. Kass-Simon, A.A. Scappaticci, Jr. The behavioral and developmental physiology of nematocysts. — 2002. — С. 1772—1794. — (Can. J. Zool. Vol. 80).
Ссылки
- [2]"Бесхозные" гены отвечают за морфологию гидр
- [3] Н. Ю. Зотова. История гидры от Антона Левенгука до наших дней.
- [4] Степаньянц С. Д., Кузнецова В. Г., Анохин Б. А. Гидра: от Абраама Трамбле до наших дней
- [5] Некоммерческая инициатива лаборатории Кильского университета по получению и использованию трансгенных гидр
- [6] The innate immune repertoire in Cnidaria — ancestral complexity and stochastic gene loss. David J Miller et al. Genome Biology 2007, 8:R59
Примечания
- ↑ Campbell Richard D. Structure of the mouth of Hydra spp. A breach in the epithelium that disappears when it closes. Cell and Tissue Research,1987, Volume 249, Number 1, p.189-197
- ↑ 1 2 Debora MacKenzie Eyeless hydra sheds light on evolution of the eye — на сайте «NewScientist», 10 марта 2010
- ↑ Holstein T., Tardent P. (1984). «An ultrahigh-speed analysis of exocytosis: nematocyst discharge». Science 223 (4638): 830–833. DOI:10.1126/science.6695186. PMID 6695186. Проверено 2012-11-11.
- ↑ Kass-Simon G., Scappaticci A. A. Jr (2002). «The behavioral and developmental physiology of nematocysts». Canadian Journal of Zoology 80: 1772–1794. DOI:10.1139/Z02-135. Проверено 2012-10-25.
- ↑ Nüchter Timm, Benoit Martin, Engel Ulrike, Özbek Suat, Holstein Thomas W. (2006). «Nanosecond-scale kinetics of nematocyst discharge». Current Biology 16 (9): R316–R318. DOI:10.1016/j.cub.2006.03.089. Проверено 2012-10-25.
- ↑ Sergey Kuznetsov, Maria Lyanguzowa, Thomas C.G. Bosch/ Role of epithelial cells and programmed cell death in Hydra spermatogenesis/ Zoology, Vol. 104, Issue 1, 2001, Pages 25-31
- ↑ Martinez, D.E. (1998) «Mortality patterns suggest lack of senescence in hydra.» Experimental Gerontology 1998 May;33(3):217-225. Full text.
- ↑ [1] Yoshida K, Fujisawa T, Hwang JS, Ikeo K, Gojobori T. Degeneration after sexual differentiation in hydra and its relevance to the evolution of aging. Gene, 2006, 385:64-70.
Категории:- Животные по алфавиту
- Гидроидные
- Модельные организмы

Wikimedia Foundation. 2010.