- МУТАЦИИ
(от лат. mutatio - изменение), внезапные (скачкообразные) естественные или вызванные искусственно наследуемые изменения генетич. материала (генома), приводящие к изменению тех или иных признаков организма. Различают генеративные М., возникающие в половых клетках и передающиеся по наследству, и соматические М., образующиеся в клетках, не участвующих в репродукции (соматич. клетках). Соматические М. приводят к возникновению генетич. мозаик, т. е. к изменению какой-то части организма, развивающейся из мутантной клетки. У растений соматические М. переносятся потомству в том случае, если растения размножают не семенами, а соматич. частями организма (напр., черенками, почками, клубнями).
Изменение генома клетки могут осуществляться тремя путями: в результате изменения числа хромосом, числа и порядка расположения генов или из-за изменения индивидуальных генов. При изменении числа хромосом (т. наз. геномные М.) может происходить утрата или приобретение одной или неск. хромосом (анеуплоидия), либо меняться число наборов хромосом (полиплоидия). Полиплоидия играет важную роль в эволюции растений и широко используется при их селекции и выведении новых сортов. У животных полиплоидия, как правило, носит летальный характер, т. к. нарушает хромосомный механизм определения пола.
Изменение расположения генов в хромосомах (т. наз. хромосомные М.) происходит в результате дупликации (повторения) гена, инверсии (переворота одного или неск. генов на 180°), транслокации, или транспозиции (переносе участка хромосомы, соизмеримого по длине с геном, в новое положение в той же или в другой хромосоме), а также делеции - выпадения участка генетич. материала (от неск. нуклеотидных пар до фрагментов, содержащих неск. генов; частный случай дефишенси-нехватка генов на конце хромосомы). При транслокации ряда генов наблюдается т. наз. э ф ф е к т п о л о ж е н и я г е н а-изменение проявления активности гена при перемещении его в др. участок хромосомы. Этим объясняется, напр., появление полосковидных глаз у дрозофилы.
Изменение индивидуальных генов (генные М.) осуществляется в результате нарушения последовательности нуклеотидных остатков в цепи ДНК данного гена. М., связанные с заменой одной пары нуклеотидных остатков в ДНК, наз. точковыми. Среди последних обычно принято различать: 1) простые замены (транзиции), когда происходит замена одного пуринового (пиримидинового) основания в нуклеотиде на другое пуриновое (пиримидиновое) основание (напр., А
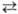 Г или Т
Г или Т 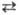 Ц; А, Г, Т и Ц-азотистые основания, соотв. аденин, гуанин, тимин и цитозин); 2) сложные, или перекрестные замены (трансверсии), когда пуриновое основание замещается на пиримидиновое и обратно (напр., А
Ц; А, Г, Т и Ц-азотистые основания, соотв. аденин, гуанин, тимин и цитозин); 2) сложные, или перекрестные замены (трансверсии), когда пуриновое основание замещается на пиримидиновое и обратно (напр., А 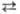 Ц, А
Ц, А 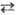 Т, Г
Т, Г 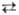 Ц, Г
Ц, Г 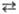 Т). При простых заменах в двойной спирали ДНК комплементарная пара А и Т замещается на пару Г и Ц, и обратно. При сложных заменах пара Г и Ц замещается на пару Ц и Г или Т и А, а пара А и Т-на пару Т и А или Ц и Г. Кроме замен, в ДНК могут происходить выпадения (делеции) или вставки одного или неск. нуклеотидных остатков. В этом случае возникают т. наз. М. со сдвигом рамки.
Т). При простых заменах в двойной спирали ДНК комплементарная пара А и Т замещается на пару Г и Ц, и обратно. При сложных заменах пара Г и Ц замещается на пару Ц и Г или Т и А, а пара А и Т-на пару Т и А или Ц и Г. Кроме замен, в ДНК могут происходить выпадения (делеции) или вставки одного или неск. нуклеотидных остатков. В этом случае возникают т. наз. М. со сдвигом рамки.
Изменения в последовательности ДНК приводят к изменению нуклеотидной последовательности в матричной РНК (мРНК; синтезируется на ДНК-матрице при транскрипции), что приводит к изменению в последовательности аминокислотных остатков в полипептидной цепи белковой молекулы, определяемой данным геном. Если в результате точко-вой М. в полипептидной цепи происходит замена одного остатка аминокислоты на другой, то имеет место т. наз. миссенс-М., или М. с изменением смысла. Если М. в ДНК приводит к образованию кодона-терминатора (и соотв. кодона-терминатора в мРНК, сигнализирующего об окончании трансляции, т. е. синтеза белковой молекулы на РНК-матрице), то процесс трансляции в данной точке останавливается. Подобная М. носит назв. нонсенс-М. или бессмысленной и, как правило, сопровождается полным выключением ф-ции фермента. При миссенс-М. не всякая замена аминокислотного остатка отражается на функцион. активности белка. Примером серьезных последствий для организма миссенс-М. может служить наследств. болезнь у человека-серповидноклеточная анемия (см. Гемоглобин).
При М. со сдвигом рамки, начиная с кодона (см. Генетический код), в к-ром потерян или приобретен нуклеотид, вся послед. аминокислотная последовательность белка при трансляции полностью меняется, что приводит к полному выключению ф-ции фермента.
Если в результате М. возникает мутантный фенотип (совокупность всех признаков и св-в организма, сформировавшихся в процессе его индивидуального развития), отличный от исходного "дикого" (наиб. распространенного в природе), то М. наз. прямой. В тех случаях, когда благодаря М. исходный дикий фенотип организма восстанавливается (полностью или частично), то М. наз. обратной или реверсией. Различают истинные реверсии, когда в результате повторной М. исходная последовательность цепи ДНК восстанавливается, и супрессорные М., к-рые локализованы в др. месте генома, но тем или иным образом компенсируют дефект, обусловленный исходной М. Наиб. интерес в этом классе М. представляют т. наз. супрессоры нонсенс-М. Мол. природа подобных обратных М. состоит в мутации гена, определяющего синтез транспортной РНК (тРНК). В этом случае в тРНК меняется последовательность нуклеотидных остатков в антикодоне [участок молекулы тРНК, состоящий из трех нуклеотидов и узнающий соответствующий ему участок из трех нуклеотидов (кодон) в молекуле мРНК] таким образом, что он приобретает способность взаимод. ("узнавать") с кодоном-терминатором (или нонсенс-кодо-ном). Благодаря этому нонсенс-кодон прочитывается как значащий, т. е. в соответствующем месте полипептидной цепи устанавливается аминокислота, и синтез белка продолжается.
По происхождению М. можно разделить на две группы: спонтанные и индуцированные. При индуцир. мутагенезе (искусств. получение М.) М. возникают в результате воздействия на организм мутагенов. Один из ключевых ферментов, определяющих частоту спонтанных М.-ДНК-поли-мераза (катализирует синтез ДНК из нуклеотидов на ДНК-матрице; см. Полидезоксирибонуклеотид-синтетазы). Частота неправильных включений нуклеотидов при репликации (самовоспроизведении) ДНК с использованием этого фермента из разл. источников (микроорганизмы, дрожжи, клетки млекопитающих) достигает довольно высоких значений-10 -4 (отношение числа мутированных нуклеотидных звеньев к общему числу мономерных звеньев ДНК). Однако в клетке существует спец. система коррекции, а также разл. системы репарации (восстановление нативной структуры) ДНК, к-рые понижают эту величину до 10-9.
Значит. роль в спонтанной М. играют специфич. мигрирующие генетические элементы. Частота М. с их участием составляет у простейших организмов (бактерий, дрожжей) ок. 10-5 на поколение, а при определенных условиях может значительно увеличиваться. В результате встраивания подобных элементов в гены может нарушаться их активность, изменяться система регуляции и т. п.
Осн. доля всех М. в природе обусловлена генными М. Они вызывают разнообразные изменения признаков. Большинство из М. вредны для организмов (могут вызывать уродство и даже гибель). Очень редко возникают М., улучшающие св-ва организма. Эти М. дают осн. материал для естеств. и искусств. отбора, являясь необходимым условием эволюции в природе и селекции полезных форм растений, животных и микроорганизмов. Частота спонтанных мутаций у каждого вида генетически обусловлена и поддерживается на оптим. уровне.
Осн. представления о М. впервые получили развитие в работах X. Де Фриза (1901), а в дальнейшем (1925-1928) в работах Г. Мёллера, Л. Стадлера, Г. Надсона, Г. С. Филиппова и др. в связи с исследованием мутагенного действия радиоактивности, рентгеновских и УФ лучей. Механизмы М. на мол. уровне стали выясняться с сер. 20 в. в связи с открытием двойной спирали ДНК Дж. Уотсоном и Ф. Криком (1953).
Лит.: АуэрбахШ., Проблемы мутагенеза, пер. с нем., М., 1978; Томилин Н. В., Генетическая стабильность клетки,. Л., 1983; Хесин Р. Б., Непостоянство генома, М., 1984; Алиханян С. И., Акифьев А. П., Чернив Л. С, Общая генетика, М., 1985. Г. Б. Завильгелъский.
Химическая энциклопедия. — М.: Советская энциклопедия. Под ред. И. Л. Кнунянца. 1988.