- ПОДТИП ХЕЛИЦЕРОВЫЕ (CHELICERATA)
- ПОДТИП ХЕЛИЦЕРОВЫЕ (CHELICERATA)
-
Хелицеровые — обширный и разнообразный подтип членистоногих. В современной фауне они представлены почти исключительно наземными формами, обычно объединяемыми в класс паукообразных (Arachnida). Однако по происхождению хелицеровые — водные членистоногие, первоначально населявшие море. Водные формы, объединяемые в класс меростомовых (Merostomata), вымерли в отдаленные геологические эпохи. Из них до нашего времени дожило только несколько видов мечехвостов (отряд Xiphosura) — удивительным образом сохранившиеся «живые ископаемые». По происхождению хелицеровые связаны с трилобитами.
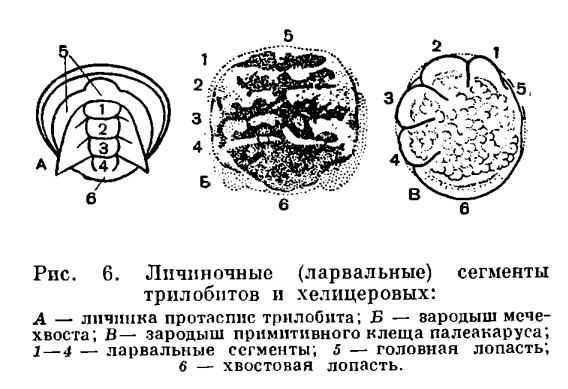
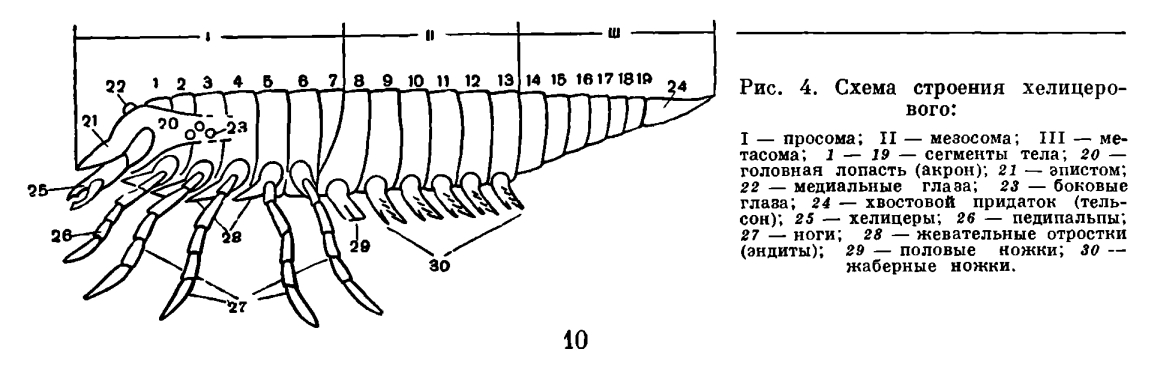 Наиболее характерные признаки хелицеровых — исчезновение усиков (антеннул) и своеобразный способ объединения сегментов тела в отделы, что связано с функциональными особенностями конечностей. Усики у всех хелицеровых отсутствуют, и их место занимает первая пара послеротовых конечностей, которые обычно имеют на конце клешни и получили особое название — хелицеры. Отсюда и название подтипа, что буквально значит «клешнеусые». Сегментальный состав отделов тела здесь иной, чем в других подтипах членистоногих, почему и отделы тела имеют особые названия. Тело делится на головогрудь -просом у — и брюшко — опистосому. Конечности головогруди служат для захвата и пережевывания пищи и для передвижения, а конечности брюшка выполняют дыхательную, половую и другие специальные функции. В состав головогруди входят элементы головной лопасти — акрона — и последующих 6 сегментов. Акрону принадлежат глаза, расположенные на щите, покрывающем головогрудь, и срединный надротовой вырост — э п и ст о м с верхней губой. Первый сегмент несет клешневидные хелицеры, которые, как говорилось, заняли место усиков перед ртом. Однако функционально они не заменяют усики, а служат для захвата пищи. За ними следует вторая пара ротовых конечностей, также имеющих особое название. Это педипальпы, или ногощупальца. Они состоят из основного членика — коксы, снабженного жевательным отростком — коксэндитом, и членистого щупальца. Среди конечностей головогруди функции педипальп наиболее разнообразны. Их основные членики с эндитами почти всегда так или иначе участвуют в приеме пищи, а щупальца в основном служат органами осязания, но могут участвовать в передвижении или захвате добычи. Конечности остальных четырех сегментов головогруди — ноги — служат для передвижения. Но у ряда форм на тазиках первой — третьей пар ног сохраняются жевательные отростки, и они также участвуют в пережевывании пищи.Головогрудь и брюшко у всех хелицеровых соединяются посредством седьмого, так называемого предполового, сегмента, который обычно лишен конечностей и в той или иной степени недоразвит. В составе брюшка объединяются две группы сегментов. Шесть передних, т. е. с восьмого по тринадцатый, имеют в той или иной степени недоразвитые видоизмененные конечности и образуют передний отдел брюшка - мезосому. Следующие за ними шесть сегментов всегда лишены конечностей и образуют задний отдел брюшка — метасому, которая оканчивается хвостовым придатком (тельсон). В пределах подтипа наблюдается постепенная атрофия сегментов метасомы и укорачивание брюшка. Первый брюшной сегмент, по общему счету восьмой, —половой, на нем всегда расположено половое отверстие, а его конечности у ряда форм образуют половые крышечки. Конечности следующих пяти сегментов у первичноводных форм представлены жаберными ножками, а у наземных превращаются в парные легкие, число которых различно и которые могут частично или полностью исчезать, заменяясь дыхательными трубками — трахеями.Во внутреннем строении хелицеровых характерна сложная система выростов кишечника — так называемая печень, клетки которой способны к внутриклеточному перевариванию пищи (фагоцитозу). Сохраняются примитивные выделительные органы — коксальные железы, соответствующие у членистоногих выделительным органам кольчатых червей — целомодуктам. Характерна также атрофия отдела мозга, иннервирующего усики, — так называемого дейтоцеребрума, что связано с их исчезновением.В подавляющем большинстве хелицеровые откладывают яйца, но есть и живородящие формы. Яйца богаты желтком, благодаря чему вылупляющаяся особь по своей организации сходна со взрослой, и послезародышевое развитие, как правило, протекает без превращений — метаморфоза, сопровождаясь ростом и линьками.Хелицеровые очень древняя группа. Первые морские представители подтипа известны уже из кембрийского периода. На родство хелицеровых с трилобитами с определенностью указал в 1924 году Д. М. Федотов, затем Стермер, и эта точка зрения принята теперь большинством зоологов. Однако хелицеровых можно сближать лишь с наиболее примитивными трилобитами, у которых задние сегменты тела не слиты в пигидий. Родство с трилобитами проявляется во многих признаках. Существенно соответствие (гомология) частей двуветвистых конечностей трилобитов и жаберных ножек водных хелицеровых. У тех и других конечность состоит из основания и двух ветвей: внутренней — собственно членистой ножки наружной — членистого придатка с жаберными листочками. У водных хелицеровых, как у трилобитов, рот расположен вентрально, в виде продольной щели между коксальными эндитами ног. Четыре передних сегмента головогруди хелицеровых соответствуют сегментам головы трилобитов и представляют ларвальные сегменты. Как говорилось, они характерны для личинки трилобитов — протасписа — и, в отличие от последующих, образуются все одновременно. Одновременное появление этих сегментов и их анатомические отличия от остальных установлены впервые у зародыша мечехвостов в 1933 году П. П. Ивановым, а недавно обнаружены автором этого раздела у примитивных клещей. У некоторых хелицеровых (сольпуг и др.) и во взрослом состоянии четыре передних сегмента объединены в отдел, соответствующий голове трилобитов, а сегменты третьей и четвертой пар ног расчленены. Постларвальные (туловищные) сегменты у мечехвостов формируются, как у трилобитов, один за другим за счет деятельности околоанальной зоны роста, с тем только различием, что у трилобитов этот процесс происходил в послезародышевом развитии, а у мечехвостов наблюдается в эмбриогенезе.
Наиболее характерные признаки хелицеровых — исчезновение усиков (антеннул) и своеобразный способ объединения сегментов тела в отделы, что связано с функциональными особенностями конечностей. Усики у всех хелицеровых отсутствуют, и их место занимает первая пара послеротовых конечностей, которые обычно имеют на конце клешни и получили особое название — хелицеры. Отсюда и название подтипа, что буквально значит «клешнеусые». Сегментальный состав отделов тела здесь иной, чем в других подтипах членистоногих, почему и отделы тела имеют особые названия. Тело делится на головогрудь -просом у — и брюшко — опистосому. Конечности головогруди служат для захвата и пережевывания пищи и для передвижения, а конечности брюшка выполняют дыхательную, половую и другие специальные функции. В состав головогруди входят элементы головной лопасти — акрона — и последующих 6 сегментов. Акрону принадлежат глаза, расположенные на щите, покрывающем головогрудь, и срединный надротовой вырост — э п и ст о м с верхней губой. Первый сегмент несет клешневидные хелицеры, которые, как говорилось, заняли место усиков перед ртом. Однако функционально они не заменяют усики, а служат для захвата пищи. За ними следует вторая пара ротовых конечностей, также имеющих особое название. Это педипальпы, или ногощупальца. Они состоят из основного членика — коксы, снабженного жевательным отростком — коксэндитом, и членистого щупальца. Среди конечностей головогруди функции педипальп наиболее разнообразны. Их основные членики с эндитами почти всегда так или иначе участвуют в приеме пищи, а щупальца в основном служат органами осязания, но могут участвовать в передвижении или захвате добычи. Конечности остальных четырех сегментов головогруди — ноги — служат для передвижения. Но у ряда форм на тазиках первой — третьей пар ног сохраняются жевательные отростки, и они также участвуют в пережевывании пищи.Головогрудь и брюшко у всех хелицеровых соединяются посредством седьмого, так называемого предполового, сегмента, который обычно лишен конечностей и в той или иной степени недоразвит. В составе брюшка объединяются две группы сегментов. Шесть передних, т. е. с восьмого по тринадцатый, имеют в той или иной степени недоразвитые видоизмененные конечности и образуют передний отдел брюшка - мезосому. Следующие за ними шесть сегментов всегда лишены конечностей и образуют задний отдел брюшка — метасому, которая оканчивается хвостовым придатком (тельсон). В пределах подтипа наблюдается постепенная атрофия сегментов метасомы и укорачивание брюшка. Первый брюшной сегмент, по общему счету восьмой, —половой, на нем всегда расположено половое отверстие, а его конечности у ряда форм образуют половые крышечки. Конечности следующих пяти сегментов у первичноводных форм представлены жаберными ножками, а у наземных превращаются в парные легкие, число которых различно и которые могут частично или полностью исчезать, заменяясь дыхательными трубками — трахеями.Во внутреннем строении хелицеровых характерна сложная система выростов кишечника — так называемая печень, клетки которой способны к внутриклеточному перевариванию пищи (фагоцитозу). Сохраняются примитивные выделительные органы — коксальные железы, соответствующие у членистоногих выделительным органам кольчатых червей — целомодуктам. Характерна также атрофия отдела мозга, иннервирующего усики, — так называемого дейтоцеребрума, что связано с их исчезновением.В подавляющем большинстве хелицеровые откладывают яйца, но есть и живородящие формы. Яйца богаты желтком, благодаря чему вылупляющаяся особь по своей организации сходна со взрослой, и послезародышевое развитие, как правило, протекает без превращений — метаморфоза, сопровождаясь ростом и линьками.Хелицеровые очень древняя группа. Первые морские представители подтипа известны уже из кембрийского периода. На родство хелицеровых с трилобитами с определенностью указал в 1924 году Д. М. Федотов, затем Стермер, и эта точка зрения принята теперь большинством зоологов. Однако хелицеровых можно сближать лишь с наиболее примитивными трилобитами, у которых задние сегменты тела не слиты в пигидий. Родство с трилобитами проявляется во многих признаках. Существенно соответствие (гомология) частей двуветвистых конечностей трилобитов и жаберных ножек водных хелицеровых. У тех и других конечность состоит из основания и двух ветвей: внутренней — собственно членистой ножки наружной — членистого придатка с жаберными листочками. У водных хелицеровых, как у трилобитов, рот расположен вентрально, в виде продольной щели между коксальными эндитами ног. Четыре передних сегмента головогруди хелицеровых соответствуют сегментам головы трилобитов и представляют ларвальные сегменты. Как говорилось, они характерны для личинки трилобитов — протасписа — и, в отличие от последующих, образуются все одновременно. Одновременное появление этих сегментов и их анатомические отличия от остальных установлены впервые у зародыша мечехвостов в 1933 году П. П. Ивановым, а недавно обнаружены автором этого раздела у примитивных клещей. У некоторых хелицеровых (сольпуг и др.) и во взрослом состоянии четыре передних сегмента объединены в отдел, соответствующий голове трилобитов, а сегменты третьей и четвертой пар ног расчленены. Постларвальные (туловищные) сегменты у мечехвостов формируются, как у трилобитов, один за другим за счет деятельности околоанальной зоны роста, с тем только различием, что у трилобитов этот процесс происходил в послезародышевом развитии, а у мечехвостов наблюдается в эмбриогенезе.
, Прототип хелицеровых наиболее полно олицетворяют древние морские меростомовые — эвриптериды (отряд Eurypterida), организация которых может быть выведена из трилобитообразной. При этом имели место две основные группы процессов: во-первых, атрофия органов чувств головы — в первую очередь усиков, а затем и сложных глаз, во-вторых, подразделение однородных трилобитных конечностей на двигательно-жевательные и дыхательные и соответственное объединение сегментов тела в отделы. От усиков, по крайней мере внешне, ни у кого из хелицеровых не осталось и следов. Сложные глаза атрофировались более постепенно, у большинства водных форм они имеются, но становятся меньше, чем у трилобитов, и в конце концов на их месте остается несколько простых глазков. Первая пара конечностей превратилась в хватательные хелицеры, следующие пять пар стали двигательно-жевательными, окружив укоротившуюся ротовую щель своими коксэндитами. В результате всех этих изменений голова как обособленный отдел исчезла, а все 6 передних сегментов и элементы головной лопасти объединились в передний отдел тела — просому. Остальные сегменты образовали брюшко — опистосому, причем конечности его переднего отдела — мезосомы — в большей своей части сохранили дыхательную функцию жабр, а конечности заднего отдела — метасомы — атрофировались. Общее число сегментов сократилось до 19; роль соединения головогруди и брюшка взял на себя седьмой сегмент, за которым всегда следует сегмент с половым отверстием.
Прототип хелицеровых наиболее полно олицетворяют древние морские меростомовые — эвриптериды (отряд Eurypterida), организация которых может быть выведена из трилобитообразной. При этом имели место две основные группы процессов: во-первых, атрофия органов чувств головы — в первую очередь усиков, а затем и сложных глаз, во-вторых, подразделение однородных трилобитных конечностей на двигательно-жевательные и дыхательные и соответственное объединение сегментов тела в отделы. От усиков, по крайней мере внешне, ни у кого из хелицеровых не осталось и следов. Сложные глаза атрофировались более постепенно, у большинства водных форм они имеются, но становятся меньше, чем у трилобитов, и в конце концов на их месте остается несколько простых глазков. Первая пара конечностей превратилась в хватательные хелицеры, следующие пять пар стали двигательно-жевательными, окружив укоротившуюся ротовую щель своими коксэндитами. В результате всех этих изменений голова как обособленный отдел исчезла, а все 6 передних сегментов и элементы головной лопасти объединились в передний отдел тела — просому. Остальные сегменты образовали брюшко — опистосому, причем конечности его переднего отдела — мезосомы — в большей своей части сохранили дыхательную функцию жабр, а конечности заднего отдела — метасомы — атрофировались. Общее число сегментов сократилось до 19; роль соединения головогруди и брюшка взял на себя седьмой сегмент, за которым всегда следует сегмент с половым отверстием. Как мы видим, характернейшая черта организации хелицеровых — это отсутствие головы как обособленного отдела, и дело тут не только в том, что сегменты головы объединились с двумя туловищными в головогрудь. Ведь голова членистоногих, например насекомых, — это подвижный передний отдел тела, на котором расположены основные органы чувств—усики и сложные глаза—и набор ротовых конечностей, специально приспособленных для приема пищи. Ни один из этих компонентов головы у хелицеровых не выражен. Усики у них исчезли, зрение ослабло, а ротовые конечности в собственном смысле (челюсти) так и не образовались — большинство конечностей просомы одновременно жевательные и двигательные. Природу этого своеобразия организации в какой-то мере раскрывает образ жизни первичноводных форм, о чем мы можем судить по ныне живущим мечехвостам и по косвенным данным в отношении вымерших эвриптерид. Это крупные формы с щитовидной головогрудью, тяжелым панцирем и ограниченными возможностями движения, в большинстве донные животные, обитатели мелководных участков моря и опресненных водоемов. По способу добывания пищи большинство водных хелицеровых, по-видимому, было сходно с современными мечехвостами. Это роющиеся бентоядные хищники — потребители малоподвижных беспозвоночных: моллюсков, кольчатых червей и т. п. (эвриптериды и древние мечехвосты, вероятно, питались также трилобитами). Они не настигают добычу в погоне, а отыскивают «на ощупь» у себя под ногами, схватывают хелицерами и измельчают жевательными отростками ног. Ил роют передним острым краем головогрудного щита, а упором при рытье служит крепкая хвостовая игла, характерная для большинства меростомовых. Естественно, что при таком образе жизни необходимость совершенствования глаз как органов предметного зрения отпадает, а длинные нитевидные усики у переднего края тела становятся помехой при описанном способе рытья. Зато развитие клешневидных хелицер, служащих для схватывания добычи, и сохранение набора жевательных эндитов близ рта на брюшной стороне весьма существенны.Значительные отличия от трилобитов возникли в индивидуальном развитии. У хелицеровых произошло обогащение яиц желтком. В результате те стадии, которые у трилобитов были свободными, послезародышевыми и развивались путем длительного первичного метаморфоза, у хелицеровых стали проходить в яйце в сокращенном виде, в порядке эмбриогенеза, и из яйца стала вылупляться особь, сразу похожая на взрослую, т. е. имело место явление, называемое эмбрионизацией развития (понятие, введенное в общей форме И. И. Шмальгаузеном и разработанное А. А. 3ахваткиным).Историческая судьба хелицеровых сложилась иначе, чем судьбы двух других подтипов членистоногих—жабродышащих (ракообразных) и трахейнодышащих. Родина ракообразных—море, здесь они возникли, и вся их длительная эволюция, за немногими исключениями, связана с водной средой. Родина трахейнодышащих, в первую очередь насекомых, — суша, здесь они сформировались, достигли невиданного расцвета и господствуют по сей день. Родиной хелицеровых была водная среда — мелководье морей и, быть может, пресные воды. Здесь сложились основные черты организации этих животных. Однако в дальнейшем, будучи уже водными членистоногими с целым комплексом приспособлений к водной среде обитания и, по-видимому, даже к довольно узкой форме жизни бентоядных хищников, хелицеровые начали выходить на сушу, что, судя по имеющимся данным, происходило не раз и в различные эпохи. В море они не удержались и в большинстве вымерли. Живым свидетельством их прошлой водной жизни осталось несколько видов мечехвостов.Переход к жизни на суше требует многих специальных приспособлений. По типу индивидуального развития хелицеровые были подготовлены к этому. Путем эмбрионизации они избавились от архаического водного метаморфоза — наследия трилобитов, при наличии которого выход на сушу вряд ли возможен. Однако по своей организации хелицеровые были типично водными членистоногими, для которых переход к сухопутной жизни был нелегкой задачей. И хотя она была разрешена наземными хелпцеровыми — паукообразными, печать водного происхождения лежит на всей их эволюционной судьбе. В дальнейшем мы увидим, что по тем формам, которые приняла жизнь хелицеровых на суше, у них наблюдается много общего. В то же время каждый отряд паукообразных имеет свои особенности приспособления к сухопутной жизни и дальнейшей эволюции на суше, что в значительной мере обусловливает разнообразие наземных форм и помогает осветить их природу. Но если общие направления эволюции хелицеровых вырисовываются достаточно отчетливо, то этого нельзя сказать о филогенетических соотношениях группировок внутри подтипа. Накопившиеся результаты сравнительного изучения современных форм и палеонтологические материалы все менее укладываются в традиционную классификацию хелицеровых, и в последнее время не раз предпринимались попытки ее пересмотра. Однако зоологи не пришли к единому мнению, и это вынуждает нас в настоящем издании придерживаться традиционного подразделения хелицеровых на два к л а с с а — меростомовых (Меrostomata) и паукообразных (Arachnida).Вообще надо заметить, что отрасль знания о хелицеровых— арахнология далеко не так разработана, как, например, наука о насекомых — энтомология. Отечественных руководств по общей арахнологии не существует; вопросы происхождения и эволюции хелицеровых затронуты в немногих специальных работах. Иностранные руководства в большинстве также специальны, а в трактовке некоторых общих вопросов не всегда совпадают с нашими представлениями.Желание придать по возможности эволюционное освещение материалу привэло к необходимости научной постановки ряда проблем, при изложении, однако, в достаточно доступной форме, требуемой настоящим изданием. Такими проблемами явились происхождение хелицеровых, выход хелицеровых на сушу и различные пути эволюции паукообразных, сравнительное рассмотрение клещей как трех независимых отрядов и ряд других. При этом особенно существенной для нас была трактовка хелицеровых в трудах наших зоологов В. Н. Беклемишева и А. А. Захват кин а.
Как мы видим, характернейшая черта организации хелицеровых — это отсутствие головы как обособленного отдела, и дело тут не только в том, что сегменты головы объединились с двумя туловищными в головогрудь. Ведь голова членистоногих, например насекомых, — это подвижный передний отдел тела, на котором расположены основные органы чувств—усики и сложные глаза—и набор ротовых конечностей, специально приспособленных для приема пищи. Ни один из этих компонентов головы у хелицеровых не выражен. Усики у них исчезли, зрение ослабло, а ротовые конечности в собственном смысле (челюсти) так и не образовались — большинство конечностей просомы одновременно жевательные и двигательные. Природу этого своеобразия организации в какой-то мере раскрывает образ жизни первичноводных форм, о чем мы можем судить по ныне живущим мечехвостам и по косвенным данным в отношении вымерших эвриптерид. Это крупные формы с щитовидной головогрудью, тяжелым панцирем и ограниченными возможностями движения, в большинстве донные животные, обитатели мелководных участков моря и опресненных водоемов. По способу добывания пищи большинство водных хелицеровых, по-видимому, было сходно с современными мечехвостами. Это роющиеся бентоядные хищники — потребители малоподвижных беспозвоночных: моллюсков, кольчатых червей и т. п. (эвриптериды и древние мечехвосты, вероятно, питались также трилобитами). Они не настигают добычу в погоне, а отыскивают «на ощупь» у себя под ногами, схватывают хелицерами и измельчают жевательными отростками ног. Ил роют передним острым краем головогрудного щита, а упором при рытье служит крепкая хвостовая игла, характерная для большинства меростомовых. Естественно, что при таком образе жизни необходимость совершенствования глаз как органов предметного зрения отпадает, а длинные нитевидные усики у переднего края тела становятся помехой при описанном способе рытья. Зато развитие клешневидных хелицер, служащих для схватывания добычи, и сохранение набора жевательных эндитов близ рта на брюшной стороне весьма существенны.Значительные отличия от трилобитов возникли в индивидуальном развитии. У хелицеровых произошло обогащение яиц желтком. В результате те стадии, которые у трилобитов были свободными, послезародышевыми и развивались путем длительного первичного метаморфоза, у хелицеровых стали проходить в яйце в сокращенном виде, в порядке эмбриогенеза, и из яйца стала вылупляться особь, сразу похожая на взрослую, т. е. имело место явление, называемое эмбрионизацией развития (понятие, введенное в общей форме И. И. Шмальгаузеном и разработанное А. А. 3ахваткиным).Историческая судьба хелицеровых сложилась иначе, чем судьбы двух других подтипов членистоногих—жабродышащих (ракообразных) и трахейнодышащих. Родина ракообразных—море, здесь они возникли, и вся их длительная эволюция, за немногими исключениями, связана с водной средой. Родина трахейнодышащих, в первую очередь насекомых, — суша, здесь они сформировались, достигли невиданного расцвета и господствуют по сей день. Родиной хелицеровых была водная среда — мелководье морей и, быть может, пресные воды. Здесь сложились основные черты организации этих животных. Однако в дальнейшем, будучи уже водными членистоногими с целым комплексом приспособлений к водной среде обитания и, по-видимому, даже к довольно узкой форме жизни бентоядных хищников, хелицеровые начали выходить на сушу, что, судя по имеющимся данным, происходило не раз и в различные эпохи. В море они не удержались и в большинстве вымерли. Живым свидетельством их прошлой водной жизни осталось несколько видов мечехвостов.Переход к жизни на суше требует многих специальных приспособлений. По типу индивидуального развития хелицеровые были подготовлены к этому. Путем эмбрионизации они избавились от архаического водного метаморфоза — наследия трилобитов, при наличии которого выход на сушу вряд ли возможен. Однако по своей организации хелицеровые были типично водными членистоногими, для которых переход к сухопутной жизни был нелегкой задачей. И хотя она была разрешена наземными хелпцеровыми — паукообразными, печать водного происхождения лежит на всей их эволюционной судьбе. В дальнейшем мы увидим, что по тем формам, которые приняла жизнь хелицеровых на суше, у них наблюдается много общего. В то же время каждый отряд паукообразных имеет свои особенности приспособления к сухопутной жизни и дальнейшей эволюции на суше, что в значительной мере обусловливает разнообразие наземных форм и помогает осветить их природу. Но если общие направления эволюции хелицеровых вырисовываются достаточно отчетливо, то этого нельзя сказать о филогенетических соотношениях группировок внутри подтипа. Накопившиеся результаты сравнительного изучения современных форм и палеонтологические материалы все менее укладываются в традиционную классификацию хелицеровых, и в последнее время не раз предпринимались попытки ее пересмотра. Однако зоологи не пришли к единому мнению, и это вынуждает нас в настоящем издании придерживаться традиционного подразделения хелицеровых на два к л а с с а — меростомовых (Меrostomata) и паукообразных (Arachnida).Вообще надо заметить, что отрасль знания о хелицеровых— арахнология далеко не так разработана, как, например, наука о насекомых — энтомология. Отечественных руководств по общей арахнологии не существует; вопросы происхождения и эволюции хелицеровых затронуты в немногих специальных работах. Иностранные руководства в большинстве также специальны, а в трактовке некоторых общих вопросов не всегда совпадают с нашими представлениями.Желание придать по возможности эволюционное освещение материалу привэло к необходимости научной постановки ряда проблем, при изложении, однако, в достаточно доступной форме, требуемой настоящим изданием. Такими проблемами явились происхождение хелицеровых, выход хелицеровых на сушу и различные пути эволюции паукообразных, сравнительное рассмотрение клещей как трех независимых отрядов и ряд других. При этом особенно существенной для нас была трактовка хелицеровых в трудах наших зоологов В. Н. Беклемишева и А. А. Захват кин а.
Жизнь животных: в 6-ти томах. — М.: Просвещение. Под редакцией профессоров Н.А.Гладкова, А.В.Михеева. 1970.
.