- Семейство злаки (Poaceae)
- Семейство злаки (Poaceae)
-
Среди всех семейств цветковых растений злаки занимают особое положение. Оно определяется не только их высокой хозяйственной ценностью, но и той большой ролью, которую они играют в сложении травянистых группировок растительности —- лугов, степей, прерий и пампасов, а также саванн. К злакам принадлежат основные пищевые растения человечества —- пшеница мягкая (Triticum aestivum), рис посевной (Oryza sativa) и кукуруза (Zea mays), а также многие другие зерновые культуры, снабжающие нас такими необходимыми продуктами, как мука и крупа. Пожалуй, не менее важным является и использование злаков в качестве кормовых растений для домашних животных. Многообразно хозяйственное значение злаков и во многих других отношениях.Известно 650 родов и: от 9000 до 10 000 видов злаков. Ареал этого семейства охватывает всю сушу земного шара, исключая территории, покрытые льдами. Мятлик (Роа), овсяница (Festuca), щучка (Deschampsia), лисохвост (Alopecurus) и некоторые другие роды злаков доходят до северного (в Арктике) и до южного (в Антарктике) пределов существования цветковых растений. Среди поднимающихся наиболее высоко в горы цветковых растений злаки также занимают одно из первых мест.Для злаков характерна относительная равномерность их распространения на Земле. В тропических странах это семейство примерно так же богато видами, как и в странах с умеренным климатом, а в Арктике злаки занимают первое место среди других семейств по количеству видов. Среди злаков относительно мало узких эндемиков, однако их приводят для Австралии 632, для Индии —- 143, для Мадагаскара —- 106, для Капской области —- 102. В СССР богаты злаками-эндемиками Средняя Азия (около 80) и Кавказ (около 60 видов). Злаки обычно нетрудно узнать уже по внешнему облику. Они обычно имеют членистые стебли с хорошо развитыми узлами и двурядно расположенные очередные листья, разделенные на охватывающее стебель влагалище, линейную или ланцетную пластинку с параллельным жилкованием и расположенный у основания пластинки перепончатый вырост, называемый язычком или лигулой. Значительное большинство злаков —- травянистые растения, однако у многих представителей подсемейства бамбуковых (Bambusoideae) высокие, сильно разветвленные в верхней части, с многочисленными узлами стебли сильно одревесневают, сохраняя, однако, типичное для злаков строение. У южноамериканских видов бамбука (Bambusa) они бывают высотой до 30 м и диаметром 20 см. У южноазиатского дендрокаламуса гигантского (Dendrocalamus giganteus) стебель высотой 40 м не уступает по росту многим деревьям. Среди бамбуковых известны также лазающие или вьющиеся, иногда колючие лианообразные формы (например, азиатская динохлоа —- Dinochloa). Жизненные формы травянистых злаков также довольно разнообразны, хотя внешне они выглядят как будто одинаково. Среди злаков много однолетников, однако значительно преобладают многолетние виды, которые могут быть дерновинными или иметь длинные ползучие корневища.Как и большинству других однодольных, злакам свойственна мочковатая корневая система, образующаяся в результате недоразвития главного корня и очень ранней замены его придаточными корнями. Уже при прорастании семени развиваются 1 —- 7 таких придаточных корешков, образующих первичную корневую систему, но уже через несколько дней из нижних сближенных узлов проростка начинают развиваться вторичные придаточные корни, из которых обычно и слагается корневая система взрослого растения. У злаков с высокими прямостоячими стеблями (например, у кукурузы) придаточные корни могут развиваться и из узлов над поверхностью почвы, выполняя, роль опорных корней.
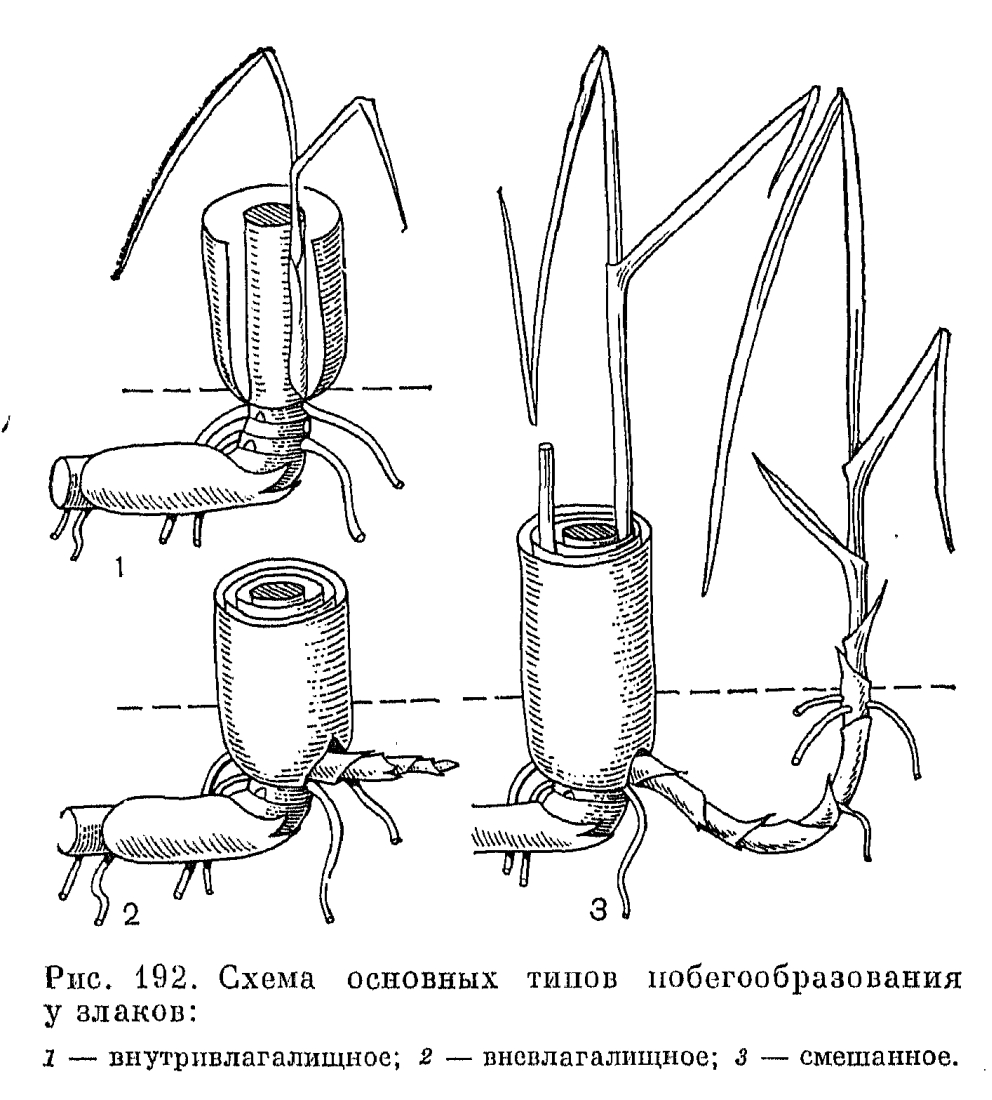 У большинства злаков ветвление побегов осуществляется лишь у их основания, где находится так называемая зона кущения, состоящая из тесно сближенных узлов. В пазухах отходящих от этих узлов листьев образуются почки, дающие начало боковым побегам. По направлению роста последние делятся на внутривлагалищные (интравагинальные) и вневлагалищные (экстравагинальные). При формировании внутривлагалищного побега (рис. 192, 1) пазушная почка растет вертикально вверх внутри влагалища своего кроющего листа. При таком способе побегообразования образуются очень густые дерновины, как у многих видов ковыля (Stipa) или у овсяницы-типчака (Festuca valesiaca). Почка вневлагалищного побега начинает расти горизонтально и пробивает своей верхушкой влагалище кроющего листа (рис. 192, 2). Такой способ побегообразования особенно характерен для видов с длинными ползучими подземными побегами-корневищами, например для пырея ползучего (Elytrigia repens). Однако нередки случаи, когда вневлагалищные побеги быстро изменяют направление своего роста на вертикальный, вследствие чего образуются дерновины, не менее густые, чем при внутривлагалищном способе побегообразования. У многих злаков известно и смешанное побегообразование, когда каждое растение образует побеги обоих типов (рис. 192).
У большинства злаков ветвление побегов осуществляется лишь у их основания, где находится так называемая зона кущения, состоящая из тесно сближенных узлов. В пазухах отходящих от этих узлов листьев образуются почки, дающие начало боковым побегам. По направлению роста последние делятся на внутривлагалищные (интравагинальные) и вневлагалищные (экстравагинальные). При формировании внутривлагалищного побега (рис. 192, 1) пазушная почка растет вертикально вверх внутри влагалища своего кроющего листа. При таком способе побегообразования образуются очень густые дерновины, как у многих видов ковыля (Stipa) или у овсяницы-типчака (Festuca valesiaca). Почка вневлагалищного побега начинает расти горизонтально и пробивает своей верхушкой влагалище кроющего листа (рис. 192, 2). Такой способ побегообразования особенно характерен для видов с длинными ползучими подземными побегами-корневищами, например для пырея ползучего (Elytrigia repens). Однако нередки случаи, когда вневлагалищные побеги быстро изменяют направление своего роста на вертикальный, вследствие чего образуются дерновины, не менее густые, чем при внутривлагалищном способе побегообразования. У многих злаков известно и смешанное побегообразование, когда каждое растение образует побеги обоих типов (рис. 192).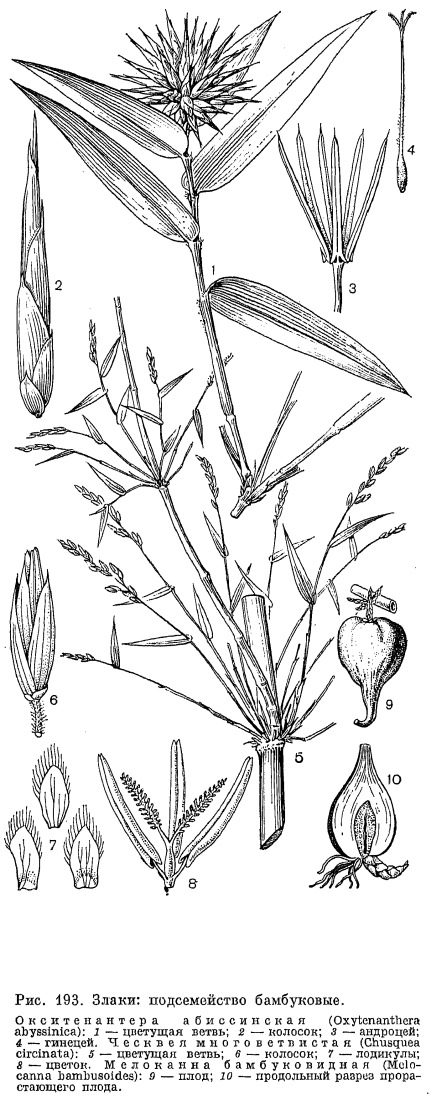
, Ветвление стеблей в их средней и верхней части у злаков внетропических стран встречается редко и обычно только у видов со стелющимися по земле стеблями (например, у прибрежницы —- Aeluropus). Значительно чаще его можно видеть у злаков тропиков, причем их боковые побеги обычно заканчиваются соцветиями. Дерновины таких злаков нередко напоминают по облику букеты или метлы. Особенно сильно разветвленные в верхней части стебли свойственны крупным бамбуковым, причем у них встречается даже мутовчатое расположение боковых ветвей, например у некоторых центральноамериканских видов ческвеи —- Chusquea (рис. 193, 5). Многие злаки со стелющимися и укореняющимися в узлах надземными побегами, например трава бизонов (Buchlоё dactyloides) североамериканских прерий (рис. 194, 6), могут формировать большие клоны, покрывающие почву густым ковром. У также североамериканской муленбергии Торрея (Muhlenbergia torreyi) и некоторых других видов такие клоны разрастаются по периферии и отмирают в середине, образуя подобие «ведьминых колец» у некоторых видов грибов.Для многолетних злаков внетропических стран весьма характерно образование нередко очень многочисленных укороченных вегетативных побегов с тесно сближенными у их основания узлами. Такие побеги могут существовать в течение одного или нескольких лет, а затем переходить к цветению. Удлиненные репродуктивные побеги формируются из них после возникновения зачатка общего соцветия за счет быстрого вставочного роста междоузлий. При этом каждый членик побега злака растет самостоятельно под защитой листового влагалища, имея свою зону вставочной меристемы. Сердцевина в растущих междоузлиях обычно быстро отмирает, и они становятся полыми, но у многих злаков тропического происхождения (например, у кукурузы) сердцевина не только сохраняется во всем стебле, но и имеет рассеянные проводящие пучки. Заполненные сердцевиной междоузлия имеются и у многих лианоподобных бамбуковых. Иногда при переходе к удлиненному репродуктивному побегу удлиняется только самое верхнее, расположенное под соцветием междоузлие, например у молинии голубой (Molinia coerulea).Как правило, стебли злаков имеют цилиндрическую форму, однако имеются и виды с сильно сплюснутыми стеблями, например широко распространенный в европейской части СССР мятлик сплюснутый (Роа compressa). Некоторые из нижних укороченных междоузлий стебля могут клубнеобразно утолщаться, выполняя функцию хранилища питательных веществ или воды. Эта особенность имеется у некоторых злаков-эфемероидов (например, у ячменя луковичного —- Hordeum bulbosum), но встречается и у мезофильных луговых видов. У мятлика дубравного (Роа sylvicola) клубневидно утолщенными становятся укороченные междоузлия ползучих подземных побегов.Признаки анатомического строения стебля используются в систематике злаков. Так, для большинства внетропических злаков, обычно называемых фестукоидными (от Festuca —- овсяница), характерны междоузлия стеблей с широкой полостью и расположением пучков проводящей ткани в 2 круга (наружный из более мелких пучков), а для преимущественно тропических —- паникоидных (от Panicum —- просо) —- междоузлия с узкой полостью или без нее и с расположением проводящих пучков многими кругами.Листья злаков располагаются всегда очередно и почти всегда двурядно. Лишь у австралийского рода микрайра (Micraira) встречается спиральное листорасположение. Листья в виде более или менее кожистых чешуи, гомологичных листовым влагалищам, обычно имеются на корневищах, а нередко также у основания надземных побегов. У многих бамбуковых опадающие чешуевидные листья без пластинок или с очень мелкими пластинками часто расположены почти по всей длине основного побега. Чешуи имеют преимущественно защитное значение и обычно следуют за самым первым листовидным органом побега —- всегда чешуевидным и обычно двукилевым предлистом.
Ветвление стеблей в их средней и верхней части у злаков внетропических стран встречается редко и обычно только у видов со стелющимися по земле стеблями (например, у прибрежницы —- Aeluropus). Значительно чаще его можно видеть у злаков тропиков, причем их боковые побеги обычно заканчиваются соцветиями. Дерновины таких злаков нередко напоминают по облику букеты или метлы. Особенно сильно разветвленные в верхней части стебли свойственны крупным бамбуковым, причем у них встречается даже мутовчатое расположение боковых ветвей, например у некоторых центральноамериканских видов ческвеи —- Chusquea (рис. 193, 5). Многие злаки со стелющимися и укореняющимися в узлах надземными побегами, например трава бизонов (Buchlоё dactyloides) североамериканских прерий (рис. 194, 6), могут формировать большие клоны, покрывающие почву густым ковром. У также североамериканской муленбергии Торрея (Muhlenbergia torreyi) и некоторых других видов такие клоны разрастаются по периферии и отмирают в середине, образуя подобие «ведьминых колец» у некоторых видов грибов.Для многолетних злаков внетропических стран весьма характерно образование нередко очень многочисленных укороченных вегетативных побегов с тесно сближенными у их основания узлами. Такие побеги могут существовать в течение одного или нескольких лет, а затем переходить к цветению. Удлиненные репродуктивные побеги формируются из них после возникновения зачатка общего соцветия за счет быстрого вставочного роста междоузлий. При этом каждый членик побега злака растет самостоятельно под защитой листового влагалища, имея свою зону вставочной меристемы. Сердцевина в растущих междоузлиях обычно быстро отмирает, и они становятся полыми, но у многих злаков тропического происхождения (например, у кукурузы) сердцевина не только сохраняется во всем стебле, но и имеет рассеянные проводящие пучки. Заполненные сердцевиной междоузлия имеются и у многих лианоподобных бамбуковых. Иногда при переходе к удлиненному репродуктивному побегу удлиняется только самое верхнее, расположенное под соцветием междоузлие, например у молинии голубой (Molinia coerulea).Как правило, стебли злаков имеют цилиндрическую форму, однако имеются и виды с сильно сплюснутыми стеблями, например широко распространенный в европейской части СССР мятлик сплюснутый (Роа compressa). Некоторые из нижних укороченных междоузлий стебля могут клубнеобразно утолщаться, выполняя функцию хранилища питательных веществ или воды. Эта особенность имеется у некоторых злаков-эфемероидов (например, у ячменя луковичного —- Hordeum bulbosum), но встречается и у мезофильных луговых видов. У мятлика дубравного (Роа sylvicola) клубневидно утолщенными становятся укороченные междоузлия ползучих подземных побегов.Признаки анатомического строения стебля используются в систематике злаков. Так, для большинства внетропических злаков, обычно называемых фестукоидными (от Festuca —- овсяница), характерны междоузлия стеблей с широкой полостью и расположением пучков проводящей ткани в 2 круга (наружный из более мелких пучков), а для преимущественно тропических —- паникоидных (от Panicum —- просо) —- междоузлия с узкой полостью или без нее и с расположением проводящих пучков многими кругами.Листья злаков располагаются всегда очередно и почти всегда двурядно. Лишь у австралийского рода микрайра (Micraira) встречается спиральное листорасположение. Листья в виде более или менее кожистых чешуи, гомологичных листовым влагалищам, обычно имеются на корневищах, а нередко также у основания надземных побегов. У многих бамбуковых опадающие чешуевидные листья без пластинок или с очень мелкими пластинками часто расположены почти по всей длине основного побега. Чешуи имеют преимущественно защитное значение и обычно следуют за самым первым листовидным органом побега —- всегда чешуевидным и обычно двукилевым предлистом. У обычных, ассимилирующих листьев влагалище образовано разросшимся в виде охватывающего стебель футляра основанием листа и служит защитой для растущего междоузлия. Влагалища злаков могут быть как до основания расщепленными (например, у преимущественно тропических триб просовых -Paniceae и сорговых —- Andropogoneae), так и сросшимися краями в трубку (у триб костровых —- Bromeae и перловниковых —- Meliceae). У некоторых видов степей и полупустынь (например, у мятлика луковичного —- Роа bulbosa, рис. 195, 4) влагалища листьев вегетативных побегов становятся запасающим органом, и побег в целом напоминает луковичку. У многих злаков отмершие влагалища нижних листьев защищают основания побегов от чрезмерного испарения или перегрева. Когда проводящие пучки влагалищ соединены между собой прочными анастомозами, у основания побегов образуется сетчато-волокнистый чехлик, характерный, например, для обычного в степях европейской части СССР костреца берегового (Bromopsis riparia).Расположенный у основания листовой пластинки и направленный вертикально вверх перепончатый или тонкокожистый вырост —- язычок, или лигула, по-видимому, препятствует проникновению воды, а с ней бактерий и спор грибков внутрь влагалища. Не случайно он хорошо развит у мезофильных и гидрофильных злаков, а у многих ксерофильных групп, особенно в подсемействе полевичковых (Eragrostoideae), видоизменен в ряд густо расположенных волосков. У большинства видов широко распространенного рода ежовник (Echinochloa) и у североамериканского рода неостапфия (Neostapfia) язычок совершенно отсутствует и влагалище переходит в пластинку без ясно выраженной границы между ними. Напротив, очень длинные (2-4 см) язычки имеются у мексиканской муленбергии хвостатой (Muhlenbergia macroura). На верхушке влагалища по бокам: от язычка у некоторых злаков (особенно у бамбуковых) имеются 2 ланцетных, часто серповидно изогнутых выроста, называемых ушками.
У обычных, ассимилирующих листьев влагалище образовано разросшимся в виде охватывающего стебель футляра основанием листа и служит защитой для растущего междоузлия. Влагалища злаков могут быть как до основания расщепленными (например, у преимущественно тропических триб просовых -Paniceae и сорговых —- Andropogoneae), так и сросшимися краями в трубку (у триб костровых —- Bromeae и перловниковых —- Meliceae). У некоторых видов степей и полупустынь (например, у мятлика луковичного —- Роа bulbosa, рис. 195, 4) влагалища листьев вегетативных побегов становятся запасающим органом, и побег в целом напоминает луковичку. У многих злаков отмершие влагалища нижних листьев защищают основания побегов от чрезмерного испарения или перегрева. Когда проводящие пучки влагалищ соединены между собой прочными анастомозами, у основания побегов образуется сетчато-волокнистый чехлик, характерный, например, для обычного в степях европейской части СССР костреца берегового (Bromopsis riparia).Расположенный у основания листовой пластинки и направленный вертикально вверх перепончатый или тонкокожистый вырост —- язычок, или лигула, по-видимому, препятствует проникновению воды, а с ней бактерий и спор грибков внутрь влагалища. Не случайно он хорошо развит у мезофильных и гидрофильных злаков, а у многих ксерофильных групп, особенно в подсемействе полевичковых (Eragrostoideae), видоизменен в ряд густо расположенных волосков. У большинства видов широко распространенного рода ежовник (Echinochloa) и у североамериканского рода неостапфия (Neostapfia) язычок совершенно отсутствует и влагалище переходит в пластинку без ясно выраженной границы между ними. Напротив, очень длинные (2-4 см) язычки имеются у мексиканской муленбергии хвостатой (Muhlenbergia macroura). На верхушке влагалища по бокам: от язычка у некоторых злаков (особенно у бамбуковых) имеются 2 ланцетных, часто серповидно изогнутых выроста, называемых ушками.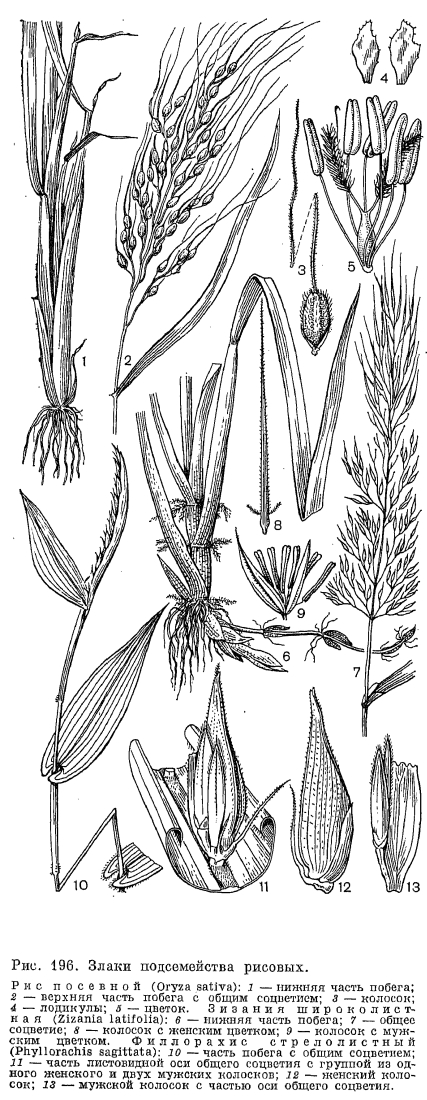
,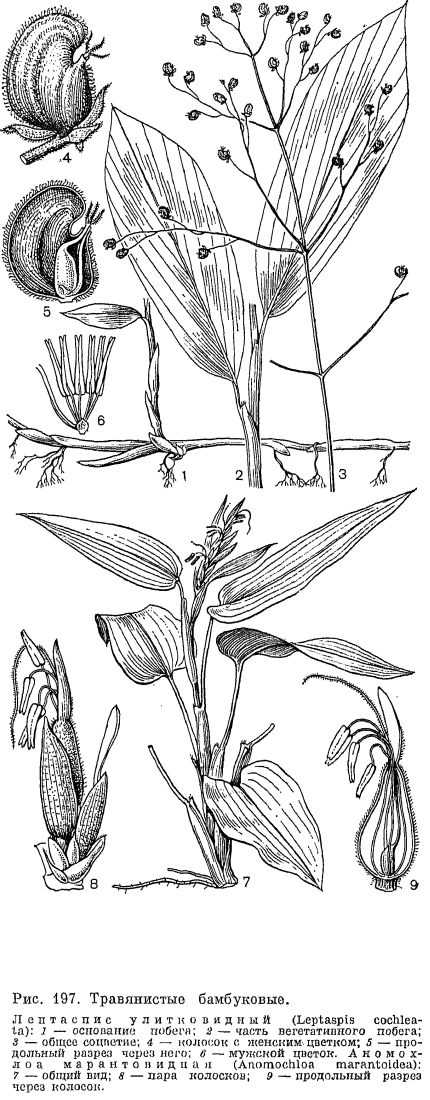 У значительного большинства злаков листовые пластинки имеют параллельное жилкование, линейную или линейно-ланцетную форму и соединены с влагалищем широким или лишь немного суженным основанием. Однако у рода артраксон (Arthraxon) и у ряда других, преимущественно тропических, родов они ланцетно-яйцевидные, а у 2 африканских родов —- филлорахиса (Phyllorachis) и умбертохлоа (Umbertochloa) —- даже стреловидные у основания (рис. 196, 10). В подсемействе бамбуковых листовые пластинки, как правило, ланцетные и у основания сужены в более пли менее развитый черешок. У бразильского травянистого бамбукового аномохлоа (Anomochloa) листовые пластинки сердцевидные и соединены с влагалищами черешком, длиной до 25 см (рис. 197, 7). Очень длинные черешки имеют также листья другого американского рода —- фарус (Pharus), имеющие еще одну, не свойственную другим злакам особенность —- перистое жилкование пластинок. У большинства бамбуковых, как и у некоторых широколистных злаков из других подсемейств, листовые пластинки имеют хорошо развитые поперечные анастомозы между параллельно проходящими основными жилками. Сильно варьируют и общие размеры листовых пластинок. У североамериканского литорального вида монантохлое прибрежного (Monanthochloe littoralis) пластинки густо расположенных листьев редко превышают в длину 1 см, а у южноамериканского бамбукового невролеписа высокого (Neurolepis elata) они длиной до 5 м и шириной 0,6 м. Очень узкие, щетиновидно вдоль сложенные или свернутые листовые пластинки имеют многие виды ковыля, овсяницы: и других, обычно ксерофильных злаков. У африканского мискантидиума щетинолистного (Miscanthidium teretifolium) очень узкие пластинки представлены почти одной только средней жилкой.
У значительного большинства злаков листовые пластинки имеют параллельное жилкование, линейную или линейно-ланцетную форму и соединены с влагалищем широким или лишь немного суженным основанием. Однако у рода артраксон (Arthraxon) и у ряда других, преимущественно тропических, родов они ланцетно-яйцевидные, а у 2 африканских родов —- филлорахиса (Phyllorachis) и умбертохлоа (Umbertochloa) —- даже стреловидные у основания (рис. 196, 10). В подсемействе бамбуковых листовые пластинки, как правило, ланцетные и у основания сужены в более пли менее развитый черешок. У бразильского травянистого бамбукового аномохлоа (Anomochloa) листовые пластинки сердцевидные и соединены с влагалищами черешком, длиной до 25 см (рис. 197, 7). Очень длинные черешки имеют также листья другого американского рода —- фарус (Pharus), имеющие еще одну, не свойственную другим злакам особенность —- перистое жилкование пластинок. У большинства бамбуковых, как и у некоторых широколистных злаков из других подсемейств, листовые пластинки имеют хорошо развитые поперечные анастомозы между параллельно проходящими основными жилками. Сильно варьируют и общие размеры листовых пластинок. У североамериканского литорального вида монантохлое прибрежного (Monanthochloe littoralis) пластинки густо расположенных листьев редко превышают в длину 1 см, а у южноамериканского бамбукового невролеписа высокого (Neurolepis elata) они длиной до 5 м и шириной 0,6 м. Очень узкие, щетиновидно вдоль сложенные или свернутые листовые пластинки имеют многие виды ковыля, овсяницы: и других, обычно ксерофильных злаков. У африканского мискантидиума щетинолистного (Miscanthidium teretifolium) очень узкие пластинки представлены почти одной только средней жилкой.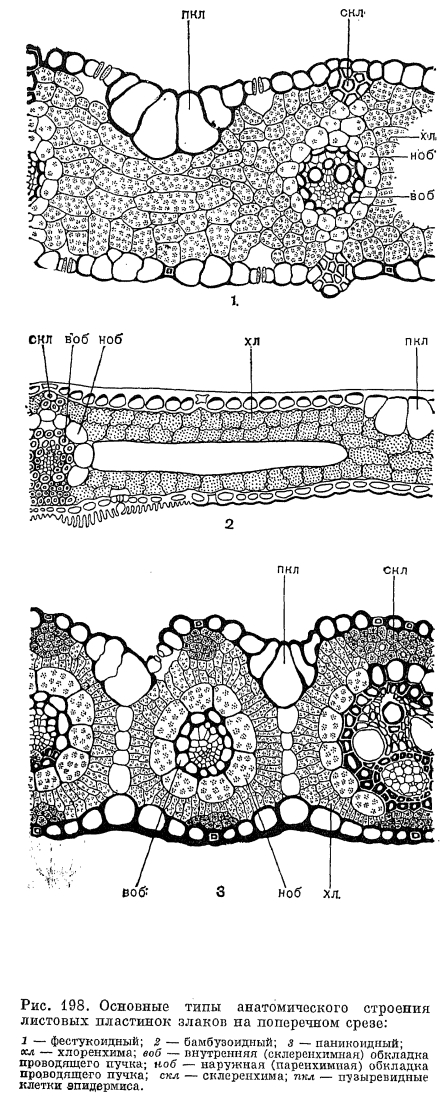 Анатомическое строение листовых пластинок как систематический признак имеет у злаков еще большую ценность, чем анатомическое строение стеблей, и обычно бывает характерным для подсемейств и триб. В настоящее время выделяют 6 основных типов анатомического строения листовых пластинок: фестукоидный, бамбузоидный (от Bambusa —- бамбук), арундиноидный (от Arundo —- арундо), паникоидный, аристидоидный (от Aristida —- триостренница) и хлоридоидный или эрагростоидный (от Chloris —- хлорис и Eragrostis —- полевичка). Для фестукоидного типа (преимущественно внетропические трибы злаков) характерно неупорядоченное расположение хлоренхимы, хорошо развитая внутренняя (склеренхимная) и относительно слабо отграниченная от хлоренхимы наружная (паренхимная) обкладки проводящих пучков (рис. 198, 1). Бамбузоидный тип, свойственный подсемейству бамбуковых, во многом сходен с фестукоидным, но отличается хлоренхимой, состоящей из своеобразных лопастных клеток, расположенных параллельными эпидермису рядами, а также более обособленной от хлоренхимы наружной обкладкой проводящих пучков (рис. 198, 2). При арундиноидном типе, свойственном подсемейству тростниковых (Arundinoideae), внутренняя обкладка пучков слабо развита, а наружная —- хорошо развита и состоит из крупных клеток без хлоропластов, клетки хлоренхимы расположены плотно и отчасти радиально вокруг пучков. Для остальных типов (преимущественно тропические подсемейства полевичковые и просовые) характерно радиальное (или венцовое) расположение хлоренхимы вокруг проводящих пучков, причем при хлоридоидном типе внутренняя (склеренхимная) обкладка пучков хорошо развита, а при паникоидном и аристидоидном типах она отсутствует или слабо развита (рис. 198, 5).Оказалось, что с радиальным (венцовым) расположением хлоренхимы и хорошо обособленной от нее наружной (паренхимной) обкладкой проводящих пучков связано много других физиологических и биохимических особенностей (так называемый кранц-синдром, от нем. kranz —- венок), прежде всего особый способ фотосинтеза —- С4 путь фиксации углекислоты, или кооперативный фотосинтез, основанный на кооперации клеток хлоренхимы и паренхимных обкладок, выполняющих разные функции. По сравнению с обычным С3 путем фиксации углекислоты этот путь очень экономичен в отношении расходования влаги и потому выгоден при обитании в аридных условиях. Преимущества кранц-синдрома можно видеть на примере обладающих им видов полевички (Eragrostis), щетинника (Setaria) и скрытницы (Crypsis) в южных районах СССР: максимум развития этих видов приходится на самое засушливое здесь время года —- июль —- август, когда большинство злаков заканчивает вегетацию.По строению эпидермы листьев, особенно окремневших клеток и волосков, приведенные выше типы анатомического строения листьев также хорошо различаются. Очень своеобразны устьица злаков. Они парацитные, с замыкающими клетками особого, так называемого граминоидного типа. В средней части эти клетки узкие с сильно утолщенными стенками, а по концам, напротив, расширенные с тонкими стенками. Такое строение позволяет регулировать ширину устьичной щели за счет расширения или сужения тонкостенных частей замыкающих клеток.Цветки злаков приспособлены к опылению ветром и имеют редуцированный околоцветник, тычинки с длинными гибкими нитями и повисающими на них пыльниками, длинные перистоволосистые рыльца и вполне сухие пыльцевые зерна с гладкой поверхностью. Они собраны в очень характерные для злаков элементарные соцветия —- колоски, которые, в свою очередь, образуют общие соцветия различного типа —- метелки, кисти, колосья или головки. Типичный многоцветковый колосок (рис. 199, 1) состоит из оси и очередно расположенных на ней двумя рядами чешуи. Две самые нижние чешуи, не несущие в своих пазухах цветков, называются колосковыми,- нижней и верхней (обычно более крупной), а выше расположенные чешуи с цветками и их пазухах —- нижними цветковыми чешуями. И те и другие гомологичны листовым влагалищам, причем нижние цветковые чешуи часто несут придатки в виде остей, которые обычно считаются гомологичными листовым пластинкам. У некоторых бамбуковых имеется более двух колосковых чешуи, а у листоколосника (Phyllostachys) такие чешуи часто несут небольшие листовые пластинки (рис. 200, 7). Напротив, у некоторых травянистых злаков одна (у плевела —- Lolium) или обе (у влагалищецветника —- Coleanthus, рис. 201, 6) колосковые чешуи могут полностью редуцироваться. Истинные колосковые чешуи по происхождению являются верховыми листьями, а не прицветниками (брактеями), как нижние цветковые чешуи. Однако во многих случаях (особенно в трибе просовых) редукция цветков в пазухах самых нижних цветковых чешуи делает последние очень похожими на дополнительные колосковые чешуи. Колосковые и нижние цветковые чешуя наиболее примитивных бамбуковых имеют, подобно листовым влагалищам, большое и непостоянное число жилок, которое в ходе эволюции семейства уменьшалось до 5, 3 или даже 1 жилки.
Анатомическое строение листовых пластинок как систематический признак имеет у злаков еще большую ценность, чем анатомическое строение стеблей, и обычно бывает характерным для подсемейств и триб. В настоящее время выделяют 6 основных типов анатомического строения листовых пластинок: фестукоидный, бамбузоидный (от Bambusa —- бамбук), арундиноидный (от Arundo —- арундо), паникоидный, аристидоидный (от Aristida —- триостренница) и хлоридоидный или эрагростоидный (от Chloris —- хлорис и Eragrostis —- полевичка). Для фестукоидного типа (преимущественно внетропические трибы злаков) характерно неупорядоченное расположение хлоренхимы, хорошо развитая внутренняя (склеренхимная) и относительно слабо отграниченная от хлоренхимы наружная (паренхимная) обкладки проводящих пучков (рис. 198, 1). Бамбузоидный тип, свойственный подсемейству бамбуковых, во многом сходен с фестукоидным, но отличается хлоренхимой, состоящей из своеобразных лопастных клеток, расположенных параллельными эпидермису рядами, а также более обособленной от хлоренхимы наружной обкладкой проводящих пучков (рис. 198, 2). При арундиноидном типе, свойственном подсемейству тростниковых (Arundinoideae), внутренняя обкладка пучков слабо развита, а наружная —- хорошо развита и состоит из крупных клеток без хлоропластов, клетки хлоренхимы расположены плотно и отчасти радиально вокруг пучков. Для остальных типов (преимущественно тропические подсемейства полевичковые и просовые) характерно радиальное (или венцовое) расположение хлоренхимы вокруг проводящих пучков, причем при хлоридоидном типе внутренняя (склеренхимная) обкладка пучков хорошо развита, а при паникоидном и аристидоидном типах она отсутствует или слабо развита (рис. 198, 5).Оказалось, что с радиальным (венцовым) расположением хлоренхимы и хорошо обособленной от нее наружной (паренхимной) обкладкой проводящих пучков связано много других физиологических и биохимических особенностей (так называемый кранц-синдром, от нем. kranz —- венок), прежде всего особый способ фотосинтеза —- С4 путь фиксации углекислоты, или кооперативный фотосинтез, основанный на кооперации клеток хлоренхимы и паренхимных обкладок, выполняющих разные функции. По сравнению с обычным С3 путем фиксации углекислоты этот путь очень экономичен в отношении расходования влаги и потому выгоден при обитании в аридных условиях. Преимущества кранц-синдрома можно видеть на примере обладающих им видов полевички (Eragrostis), щетинника (Setaria) и скрытницы (Crypsis) в южных районах СССР: максимум развития этих видов приходится на самое засушливое здесь время года —- июль —- август, когда большинство злаков заканчивает вегетацию.По строению эпидермы листьев, особенно окремневших клеток и волосков, приведенные выше типы анатомического строения листьев также хорошо различаются. Очень своеобразны устьица злаков. Они парацитные, с замыкающими клетками особого, так называемого граминоидного типа. В средней части эти клетки узкие с сильно утолщенными стенками, а по концам, напротив, расширенные с тонкими стенками. Такое строение позволяет регулировать ширину устьичной щели за счет расширения или сужения тонкостенных частей замыкающих клеток.Цветки злаков приспособлены к опылению ветром и имеют редуцированный околоцветник, тычинки с длинными гибкими нитями и повисающими на них пыльниками, длинные перистоволосистые рыльца и вполне сухие пыльцевые зерна с гладкой поверхностью. Они собраны в очень характерные для злаков элементарные соцветия —- колоски, которые, в свою очередь, образуют общие соцветия различного типа —- метелки, кисти, колосья или головки. Типичный многоцветковый колосок (рис. 199, 1) состоит из оси и очередно расположенных на ней двумя рядами чешуи. Две самые нижние чешуи, не несущие в своих пазухах цветков, называются колосковыми,- нижней и верхней (обычно более крупной), а выше расположенные чешуи с цветками и их пазухах —- нижними цветковыми чешуями. И те и другие гомологичны листовым влагалищам, причем нижние цветковые чешуи часто несут придатки в виде остей, которые обычно считаются гомологичными листовым пластинкам. У некоторых бамбуковых имеется более двух колосковых чешуи, а у листоколосника (Phyllostachys) такие чешуи часто несут небольшие листовые пластинки (рис. 200, 7). Напротив, у некоторых травянистых злаков одна (у плевела —- Lolium) или обе (у влагалищецветника —- Coleanthus, рис. 201, 6) колосковые чешуи могут полностью редуцироваться. Истинные колосковые чешуи по происхождению являются верховыми листьями, а не прицветниками (брактеями), как нижние цветковые чешуи. Однако во многих случаях (особенно в трибе просовых) редукция цветков в пазухах самых нижних цветковых чешуи делает последние очень похожими на дополнительные колосковые чешуи. Колосковые и нижние цветковые чешуя наиболее примитивных бамбуковых имеют, подобно листовым влагалищам, большое и непостоянное число жилок, которое в ходе эволюции семейства уменьшалось до 5, 3 или даже 1 жилки.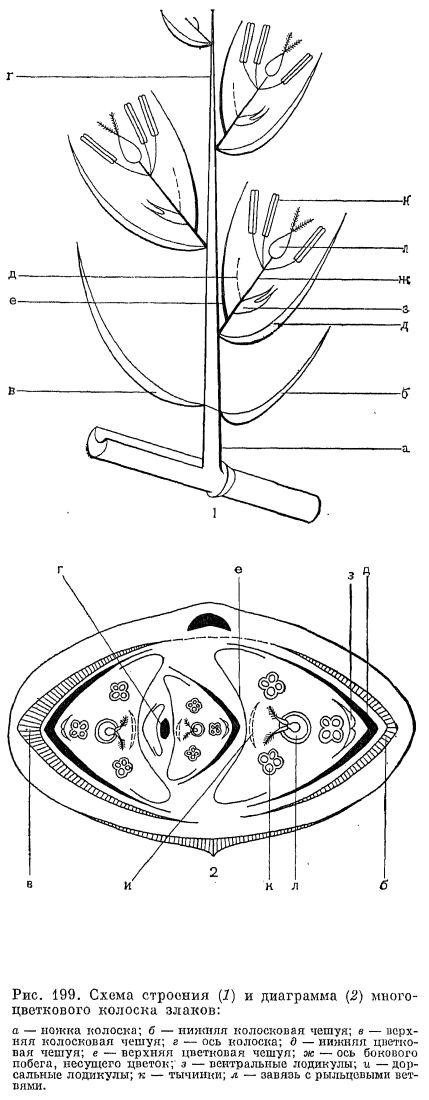
,
, Количество цветков в колосках может варьировать от очень большого и неопределенного (например, у двуколоски —- Trachynia —- до 30 цветков, рис. 201, 14, 15} до постоянно одного (у вейника или лисохвоста) или двух (у аиры —- Aira). Очень примитивные многоцветковые колоски с сильно удлиненной и часто разветвленной осью имеет китайский бамбук многоветочник длинноколосковый (Pleioblastus dolichanthus). Такие колоски больше похожи не на колоски, а на веточки метельчатого общего соцветия (рис. 200, 1). Еще менее различимы колоски в общих соцветиях тропического бамбука мелоканны (Melocanna). У него в пазухах расставленных нижних цветковых чешуи помещаются не 1, а 2 или 3 цветка на снабженных прицветничками боковых осях. Вполне вероятно, что эволюция общих соцветий у злаков шла от таких, еще не дифференцированных на колоски общих соцветий к соцветиям с хорошо обособленными, сначала многоцветковыми, а затем одноцветковыми колосками.Ось многоцветкового колоска обычно имеет сочленения под каждой нижней цветковой чешуей и при плодах распадается на членики. Основание нижней цветковой чешуи, срастаясь с таким члеником, образует утолщенный каллус, который может быть длинным и острым, как у ковыля. Часть колоска, включающую один цветок, цветковые чешуи и прилежащий к ним членик оси колоска нередко называют антецием. В одиоцветковых колосках сочленения под нижней цветковой чешуей может не быть, и тогда колоски опадают при плодах целиком.
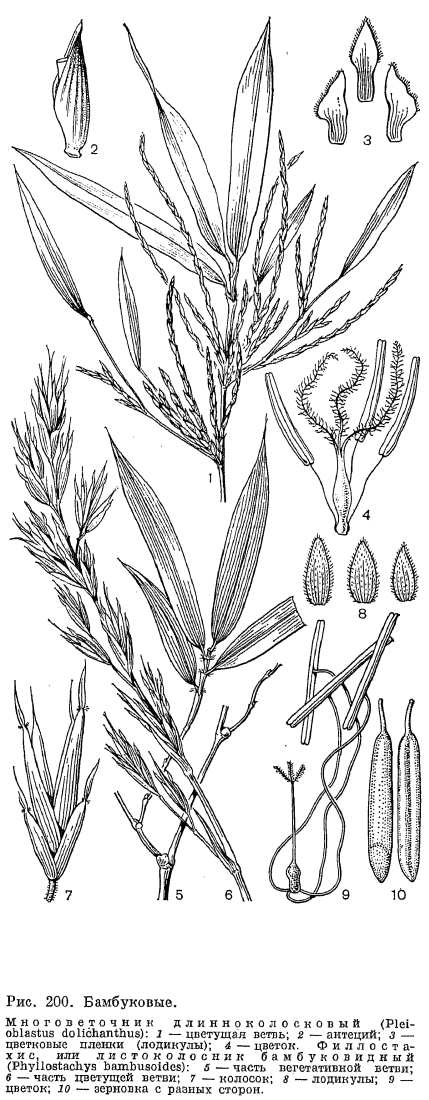
Количество цветков в колосках может варьировать от очень большого и неопределенного (например, у двуколоски —- Trachynia —- до 30 цветков, рис. 201, 14, 15} до постоянно одного (у вейника или лисохвоста) или двух (у аиры —- Aira). Очень примитивные многоцветковые колоски с сильно удлиненной и часто разветвленной осью имеет китайский бамбук многоветочник длинноколосковый (Pleioblastus dolichanthus). Такие колоски больше похожи не на колоски, а на веточки метельчатого общего соцветия (рис. 200, 1). Еще менее различимы колоски в общих соцветиях тропического бамбука мелоканны (Melocanna). У него в пазухах расставленных нижних цветковых чешуи помещаются не 1, а 2 или 3 цветка на снабженных прицветничками боковых осях. Вполне вероятно, что эволюция общих соцветий у злаков шла от таких, еще не дифференцированных на колоски общих соцветий к соцветиям с хорошо обособленными, сначала многоцветковыми, а затем одноцветковыми колосками.Ось многоцветкового колоска обычно имеет сочленения под каждой нижней цветковой чешуей и при плодах распадается на членики. Основание нижней цветковой чешуи, срастаясь с таким члеником, образует утолщенный каллус, который может быть длинным и острым, как у ковыля. Часть колоска, включающую один цветок, цветковые чешуи и прилежащий к ним членик оси колоска нередко называют антецием. В одиоцветковых колосках сочленения под нижней цветковой чешуей может не быть, и тогда колоски опадают при плодах целиком.
,
,
,
,Общие соцветия злаков обычно имеют вид метелки, нередко очень густой и колосовидной, кисти или колоса. Лишь мелкие экземпляры двуколоски (рис. 201, 14), видов костра (Bromus) и некоторых других злаков несут на верхушке стебля лишь один крупный колосок. Встречаются также очень густые, головкообразные общие соцветия, например у африканского бамбукового окситенантеры абиссинской (Охуtenanthera abyssinica, рис. 193, 1) или у средиземноморских эфемеров ежовницы (Echinaria, рис. 201, 11), и песочницы (Ammochloa, рис. 201, 7). У колючещетинника (Cenchrus) общее соцветие состоит из нескольких колючих головок (рис. 202, 8, 9). Результатом более высокой специализации общих соцветий является и упорядоченное расположение колосков по одному или группами по 2-3 на одной стороне сплюснутых осей колосовидных веточек, которые, в свою очередь, могут быть расположенными очередно или пальчато (как у свинороя —- Cynodon, рис. 194, 4). При таком расположении колосков, особенно характерном для триб просовых, сорговых и свинороевых, часть колосков на колосовидных веточках (обычно расположенные на ножках рядом с сидячими обоеполыми колосками) могут быть мужскими или вообще имеющими лишь рудимент цветка. У артраксона из трибы сорговых от колоска на ножке остается только ножка с едва заметным рудиментом колоска. Однополые колоски встречаются у злаков вообще не так уж редко. В этом случае колоски с мужскими и колоски с женскими цветками могут располагаться в пределах одного и того же соцветия (у зизании —- Zizania, рис. 196, 7, 9), на разных соцветиях одного и того же растения (у кукурузы) или на разных растениях (у пампасской травы, или кортадерии Селло —- Cortaderia selloana, табл. 45, 3, 4). В пазухах нижних цветковых чешуи со стороны оси колоска располагается еще одна чешуя, обычно имеющая 2 киля и более или менее заметную выемку на верхушке. Поскольку она принадлежит не оси колоска, а оси цветка и, следовательно, располагается выше основания нижней цветковой чешуи, ее называют верхней цветковой чешуей. Прежде Л. Челаковский (1889, 1894) и другие авторы принимали ее за 2 сросшихся сегмента наружного круга околоцветника, однако в настоящее время большинство авторов считают ее предлистом расположенного в пазухе нижней цветковой чешуи сильно укороченного побега, несущего цветок. У некоторых родов злаков (например, у лисохвоста) верхняя цветковая чешуя может полностью редуцироваться, а у очень оригинального американского травянистого бамбукового стрептохеты (Streptochaeta) она почти до основания расщеплена.Выше верхней цветковой чешуи на оси цветка значительного большинства злаков располагаются 2 маленькие бесцветные чешуйки, называемые цветковыми пленками или лодикулами. В отношении их природы пока еще нет единого мнения. Одни авторы принимают их за рудименты одного из двух трехчленных кругов околоцветника, другие —- за рудименты прицветничков. Присутствие у многих бамбуковых, а также у родов трибы ковылевых третьей, дорсальной лодикулы как будто подтверждает первую из этих, точек зрения, хотя дорсальная лодикула обычно отличается по строению от двух вентральных, обычно тесно сближенных и нередко соединенных друг с другом при основании.
В пазухах нижних цветковых чешуи со стороны оси колоска располагается еще одна чешуя, обычно имеющая 2 киля и более или менее заметную выемку на верхушке. Поскольку она принадлежит не оси колоска, а оси цветка и, следовательно, располагается выше основания нижней цветковой чешуи, ее называют верхней цветковой чешуей. Прежде Л. Челаковский (1889, 1894) и другие авторы принимали ее за 2 сросшихся сегмента наружного круга околоцветника, однако в настоящее время большинство авторов считают ее предлистом расположенного в пазухе нижней цветковой чешуи сильно укороченного побега, несущего цветок. У некоторых родов злаков (например, у лисохвоста) верхняя цветковая чешуя может полностью редуцироваться, а у очень оригинального американского травянистого бамбукового стрептохеты (Streptochaeta) она почти до основания расщеплена.Выше верхней цветковой чешуи на оси цветка значительного большинства злаков располагаются 2 маленькие бесцветные чешуйки, называемые цветковыми пленками или лодикулами. В отношении их природы пока еще нет единого мнения. Одни авторы принимают их за рудименты одного из двух трехчленных кругов околоцветника, другие —- за рудименты прицветничков. Присутствие у многих бамбуковых, а также у родов трибы ковылевых третьей, дорсальной лодикулы как будто подтверждает первую из этих, точек зрения, хотя дорсальная лодикула обычно отличается по строению от двух вентральных, обычно тесно сближенных и нередко соединенных друг с другом при основании.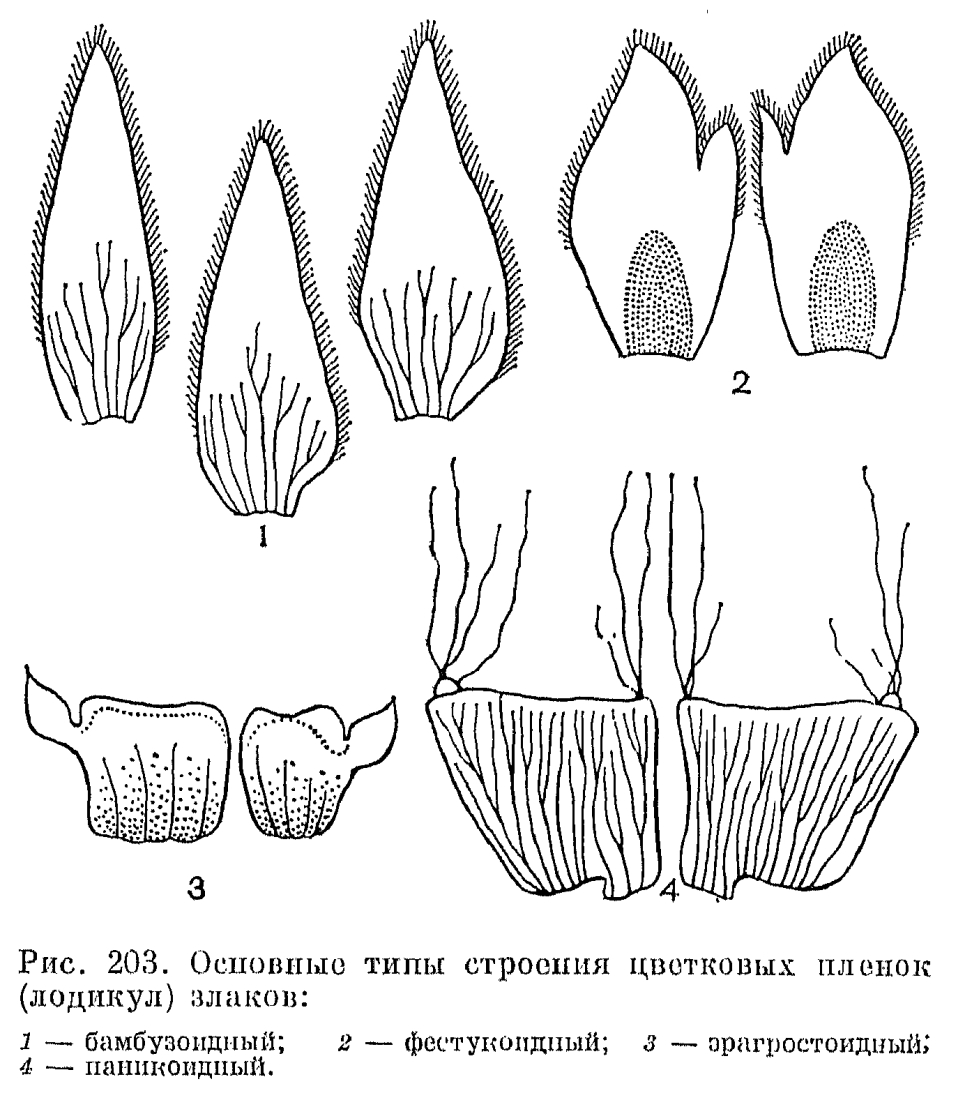 Строение лодикул считается важным систематическим признаком, характерным для целых триб злаков (рис. 203). Крупные чешуевидные лодикулы с проводящими пучками есть у многих бамбуковых, где они имеют преимущественно защитную функцию. У большинства же других злаков лодикулы имеют вид маленьких цельных или двулопастных чешуек, лишенных или почти лишенных проводящих пучков и в нижней половине сильно утолщенных. Предполагают, что такие лодикулы накапливают в себе питательные вещества для развития завязи, регулируют водный режим цветка и способствуют раздвиганию цветковых чешуи при цветении. Обычно различают 4 основных типа строения лодикул: бамбузоидный, фестукоидный, паникоидный и хлоридоидный, соответствующие основным типам анатомии листьев. Нередко выделяется еще меликоидный тип (от Melica —- перловник), свойственный трибе перловниковых (Meliceae): очень короткие (как бы обрубленные в верхней части) лодикулы слипаются друг с другом своими передними краями. 3 крупные, спирально расположенные лодикулы имеются у упомянутой выше стрептохеты, однако не все авторы принимают их за лодикулы. Наконец, у многих родов (в том числе у лисохвоста и влагалищецветника) лодикулы полностью редуцированы.Наиболее примитивное число тычинок —- 6 —- встречается среди злаков только у многих бамбуковых и рисовых (Oryzoideae). Значительное большинство злаков имеют 3 тычинки, а у некоторых родов их количество уменьшается до 2 (у душистого колоска —- Anthoxanthum) или до 1 (у цинны —- Cinna). Очень варьирует число и строение тычинок в подсемействе бамбуковых. Так, у южноазиатского рода охландра (Ochlandra) нити тычинок многократно ветвятся, вследствие чего в одном цветке может быть до 50-120 тычинок. У родов гигантохлоя (Gigantochloa) и окситенантера (Oxytenanthera) нити 6 тычинок срастаются в довольно длинную трубку, окружающую завязь (рис. 193, 3). У бразильской аномохлои 4 тычинки. Нити тычинок злаков способны быстро удлиняться при цветении. Так, у риса они удлиняются на 2,5 мм в минуту. Пыльцевые зерна злаков всегда однопоровые с гладкой и сухой оболочкой, что является приспособлением к ветроопылению.О строении гинецея в цветке злаков пока еще нет единого мнения. Согласно более широко распространенной точке зрения, гинецей злаков образован 3 сросшимися своими краями плодолистиками, а плод злаков —- зерновка —- является разновидностью паракарпного плода. Согласно другой точке зрения, гинецей злаков образован одним плодолистиком, что является следствием редукции двух других плодолистиков первично 3-членного апокарпного гинецея. Завязь всегда одногнездная с одним семязачатком, который может быть от ортотропного до гемитропного (редко кампилотропного) с направленным вниз микропиле. Интегумент обычно двойной, но у аномального во многих других отношениях рода мелоканна он простой. Обычно завязь переходит на верхушке в 2 перистоволосистые рыльцевые ветви, однако у многих бамбуковых их может быть и 3. Голые основания рыльцевых ветвей очень различаются по длине в разных трибах. Особенно длинные они у преимущественно тропической трибы просовых, что, по-видимому, связано с более тесно сомкнутыми цветковыми чешуями. У некоторых злаков рыльцевые ветви могут быть по всей или почти по всей длине сросшимися друг с другом. Так, у кукурузы свободными являются только верхние части очень длинных рыльцевых ветвей, а у белоуса (Nardus) завязь переходит на верхушке в совершенно цельное нитевидное рыльце, покрытое не волосками, как у других злаков, а короткими сосочками. У бамбукового —- стрептогины (Streptogyna) покрытые шипиками рыльцевые ветви после цветения становятся очень жесткими и служат для распространения зерновок (рис. 204, 4).
Строение лодикул считается важным систематическим признаком, характерным для целых триб злаков (рис. 203). Крупные чешуевидные лодикулы с проводящими пучками есть у многих бамбуковых, где они имеют преимущественно защитную функцию. У большинства же других злаков лодикулы имеют вид маленьких цельных или двулопастных чешуек, лишенных или почти лишенных проводящих пучков и в нижней половине сильно утолщенных. Предполагают, что такие лодикулы накапливают в себе питательные вещества для развития завязи, регулируют водный режим цветка и способствуют раздвиганию цветковых чешуи при цветении. Обычно различают 4 основных типа строения лодикул: бамбузоидный, фестукоидный, паникоидный и хлоридоидный, соответствующие основным типам анатомии листьев. Нередко выделяется еще меликоидный тип (от Melica —- перловник), свойственный трибе перловниковых (Meliceae): очень короткие (как бы обрубленные в верхней части) лодикулы слипаются друг с другом своими передними краями. 3 крупные, спирально расположенные лодикулы имеются у упомянутой выше стрептохеты, однако не все авторы принимают их за лодикулы. Наконец, у многих родов (в том числе у лисохвоста и влагалищецветника) лодикулы полностью редуцированы.Наиболее примитивное число тычинок —- 6 —- встречается среди злаков только у многих бамбуковых и рисовых (Oryzoideae). Значительное большинство злаков имеют 3 тычинки, а у некоторых родов их количество уменьшается до 2 (у душистого колоска —- Anthoxanthum) или до 1 (у цинны —- Cinna). Очень варьирует число и строение тычинок в подсемействе бамбуковых. Так, у южноазиатского рода охландра (Ochlandra) нити тычинок многократно ветвятся, вследствие чего в одном цветке может быть до 50-120 тычинок. У родов гигантохлоя (Gigantochloa) и окситенантера (Oxytenanthera) нити 6 тычинок срастаются в довольно длинную трубку, окружающую завязь (рис. 193, 3). У бразильской аномохлои 4 тычинки. Нити тычинок злаков способны быстро удлиняться при цветении. Так, у риса они удлиняются на 2,5 мм в минуту. Пыльцевые зерна злаков всегда однопоровые с гладкой и сухой оболочкой, что является приспособлением к ветроопылению.О строении гинецея в цветке злаков пока еще нет единого мнения. Согласно более широко распространенной точке зрения, гинецей злаков образован 3 сросшимися своими краями плодолистиками, а плод злаков —- зерновка —- является разновидностью паракарпного плода. Согласно другой точке зрения, гинецей злаков образован одним плодолистиком, что является следствием редукции двух других плодолистиков первично 3-членного апокарпного гинецея. Завязь всегда одногнездная с одним семязачатком, который может быть от ортотропного до гемитропного (редко кампилотропного) с направленным вниз микропиле. Интегумент обычно двойной, но у аномального во многих других отношениях рода мелоканна он простой. Обычно завязь переходит на верхушке в 2 перистоволосистые рыльцевые ветви, однако у многих бамбуковых их может быть и 3. Голые основания рыльцевых ветвей очень различаются по длине в разных трибах. Особенно длинные они у преимущественно тропической трибы просовых, что, по-видимому, связано с более тесно сомкнутыми цветковыми чешуями. У некоторых злаков рыльцевые ветви могут быть по всей или почти по всей длине сросшимися друг с другом. Так, у кукурузы свободными являются только верхние части очень длинных рыльцевых ветвей, а у белоуса (Nardus) завязь переходит на верхушке в совершенно цельное нитевидное рыльце, покрытое не волосками, как у других злаков, а короткими сосочками. У бамбукового —- стрептогины (Streptogyna) покрытые шипиками рыльцевые ветви после цветения становятся очень жесткими и служат для распространения зерновок (рис. 204, 4).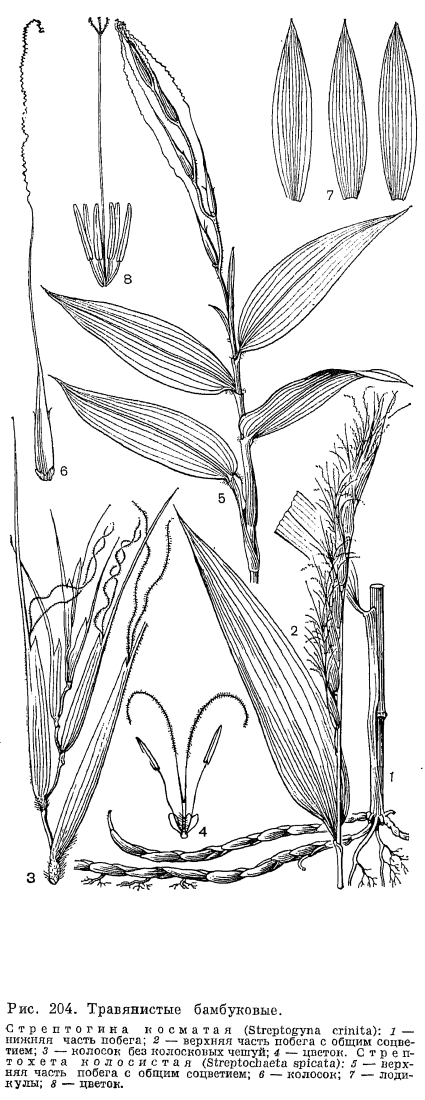 Невскрывающийся сухой односемянный плод злаков, называемый зерновкой, имеет тонкий околоплодник, обычно настолько плотно прилегающий к семенной кожуре, что кажется с ней сросшимся. Нередко при созревании зерновки ее околоплодник слипается и с плотно прилегающими к нему цветковыми чешуями. У споробола (Sporobolus) околоплодник остается разъединенным с семенем и зерновки в этом случае называются мешочковидными. Форма зерновок варьирует от почти шаровидной (у проса) до узкоцилиндрической (у многих ковылей). На выпуклой, плоской или вогнутой в виде продольного желобка брюшной (вентральной) стороне зерновки имеется рубчик, или гилум, обычно окрашенный в более темный цвет по сравнению с остальной частью зерновки и имеющий форму от почти округлого (у мятлика) до линейного и почти равного по длине всей зерновки (у пшеницы). Рубчик является местом прикрепления семязачатка к семяножке (фуникулусу), и его форма определяется ориентацией семязачатка.Наиболее оригинальны по своему строению зерновки некоторых бамбуковых, которые могут быть ягодовидными с толстым мясистым околоплодником или ореховидными с довольно толстым и очень твердым по консистенции околоплодником, отделенным от семенной кожуры. У распространенной в Юго-Восточной Азии мелоканны ягодовидные зерновки имеют обратногрушевидную форму и достигают в поперечнике 3-6 см (рис. 193, 9, 10). У них есть еще одна особенность, отсутствующая у всех других злаков: в ходе развития зародыша эндосперм семени полностью усваивается зародышем и в зрелой зерновке от него остается только сухая пленка между околоплодником, и сильно разросшимся щитком.У всех других злаков большую часть зрелой зерновки составляет эндосперм, причем соотношение в размерах эндосперма и зародыша имеет существенное систематическое значение. Так, для фестукоидных злаков характерны, относительно небольшие размеры зародыша, а для паникоидных —- более крупные по сравнению с эндоспермом. Обычно эндосперм зрелых зерновок твердый по консистенции, но может быть более рыхлым —- мучнистым, когда в нем мало белков, или более плотным —- стекловидным при относительно большом содержании белков. Можно отметить, что в эндосперме зерновок злаков содержатся очень характерные для них и не встречающиеся у других растений белки проламины. В зерновках некоторых злаков (особенно из трибы овсовых) эндосперм, особенно богат маслами и сохраняет полужидкую (желеобразную) консистенцию в период их полной зрелости. Такой эндосперм отличается необыкновенной устойчивостью к высыханию, сохраняя полужидкую консистенцию даже у зерновок, хранившихся в гербариях свыше 50 лет.Крахмальные зерна эндосперма имеют разное строение в разных группах злаков. Так, у пшеницы и других представителей трибы пшеницевых они простые, очень варьирующие по величине и без заметных граней на своей поверхности (тритикоидный тип, от лат. Triticum —- пшеница); у проса и других паникоидных злаков они тоже простые, но менее варьируют по величине и имеют гранистую поверхность, а у овсяницы и многих других фестукоидных злаков крахмальные зерна сложные, состоящие из более мелких гранул (рис. 205).
Невскрывающийся сухой односемянный плод злаков, называемый зерновкой, имеет тонкий околоплодник, обычно настолько плотно прилегающий к семенной кожуре, что кажется с ней сросшимся. Нередко при созревании зерновки ее околоплодник слипается и с плотно прилегающими к нему цветковыми чешуями. У споробола (Sporobolus) околоплодник остается разъединенным с семенем и зерновки в этом случае называются мешочковидными. Форма зерновок варьирует от почти шаровидной (у проса) до узкоцилиндрической (у многих ковылей). На выпуклой, плоской или вогнутой в виде продольного желобка брюшной (вентральной) стороне зерновки имеется рубчик, или гилум, обычно окрашенный в более темный цвет по сравнению с остальной частью зерновки и имеющий форму от почти округлого (у мятлика) до линейного и почти равного по длине всей зерновки (у пшеницы). Рубчик является местом прикрепления семязачатка к семяножке (фуникулусу), и его форма определяется ориентацией семязачатка.Наиболее оригинальны по своему строению зерновки некоторых бамбуковых, которые могут быть ягодовидными с толстым мясистым околоплодником или ореховидными с довольно толстым и очень твердым по консистенции околоплодником, отделенным от семенной кожуры. У распространенной в Юго-Восточной Азии мелоканны ягодовидные зерновки имеют обратногрушевидную форму и достигают в поперечнике 3-6 см (рис. 193, 9, 10). У них есть еще одна особенность, отсутствующая у всех других злаков: в ходе развития зародыша эндосперм семени полностью усваивается зародышем и в зрелой зерновке от него остается только сухая пленка между околоплодником, и сильно разросшимся щитком.У всех других злаков большую часть зрелой зерновки составляет эндосперм, причем соотношение в размерах эндосперма и зародыша имеет существенное систематическое значение. Так, для фестукоидных злаков характерны, относительно небольшие размеры зародыша, а для паникоидных —- более крупные по сравнению с эндоспермом. Обычно эндосперм зрелых зерновок твердый по консистенции, но может быть более рыхлым —- мучнистым, когда в нем мало белков, или более плотным —- стекловидным при относительно большом содержании белков. Можно отметить, что в эндосперме зерновок злаков содержатся очень характерные для них и не встречающиеся у других растений белки проламины. В зерновках некоторых злаков (особенно из трибы овсовых) эндосперм, особенно богат маслами и сохраняет полужидкую (желеобразную) консистенцию в период их полной зрелости. Такой эндосперм отличается необыкновенной устойчивостью к высыханию, сохраняя полужидкую консистенцию даже у зерновок, хранившихся в гербариях свыше 50 лет.Крахмальные зерна эндосперма имеют разное строение в разных группах злаков. Так, у пшеницы и других представителей трибы пшеницевых они простые, очень варьирующие по величине и без заметных граней на своей поверхности (тритикоидный тип, от лат. Triticum —- пшеница); у проса и других паникоидных злаков они тоже простые, но менее варьируют по величине и имеют гранистую поверхность, а у овсяницы и многих других фестукоидных злаков крахмальные зерна сложные, состоящие из более мелких гранул (рис. 205).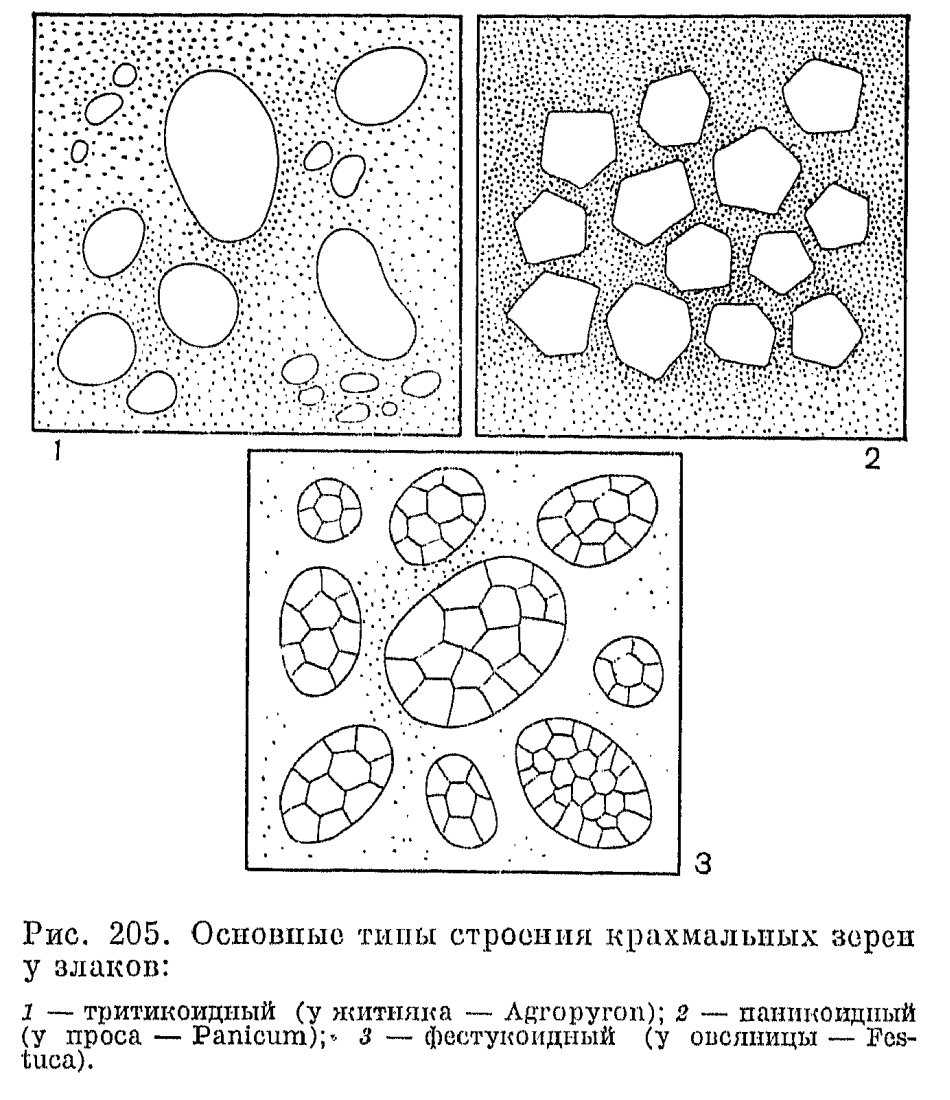
,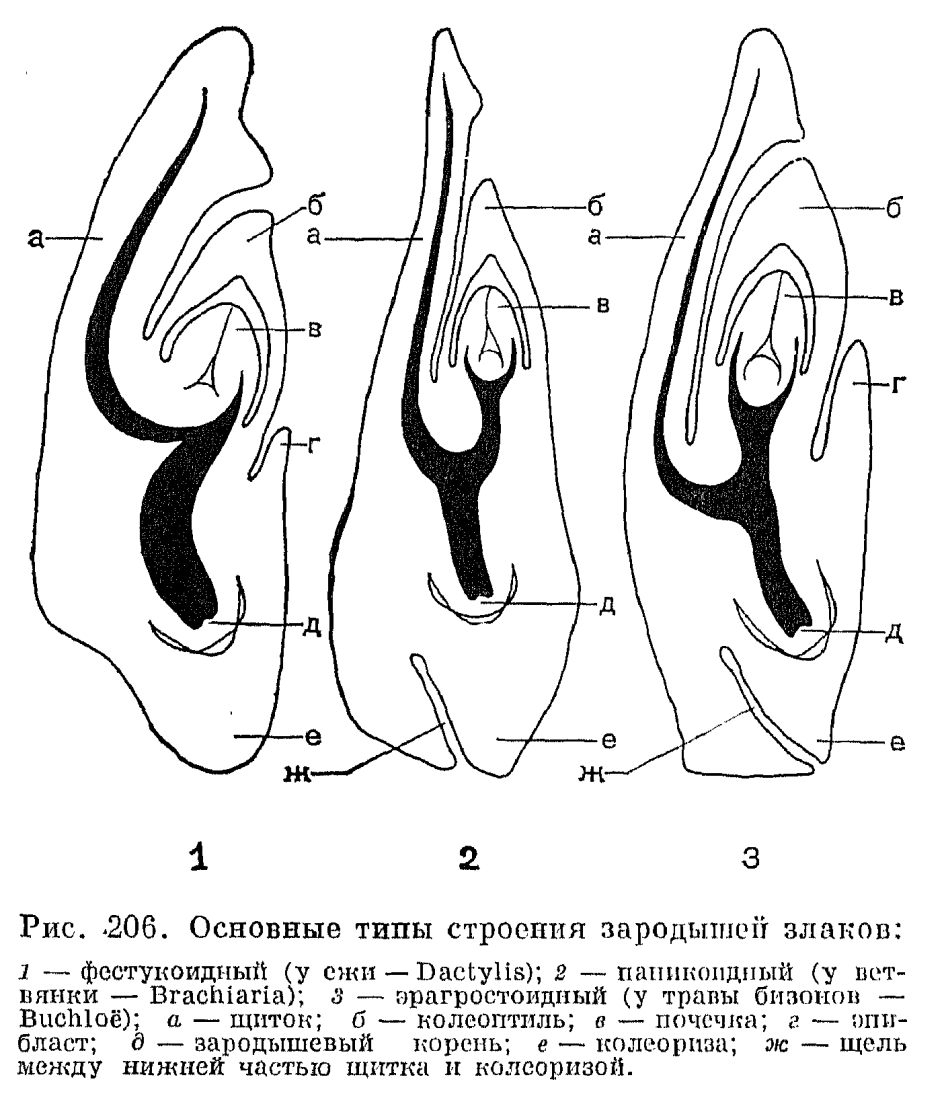 Зародыш злаков (рис. 206) довольно сильно отличается по своему строению от зародышей других однодольных. На стороне, прилегающей к эндосперму, он имеет щитовидное тело —- щиток. Снаружи от него и ближе к его верхней части находится зародышевая почечка, одетая двукилевым влагалищеобразным листом —- колеоптилем. У многих злаков против щитка с наружной стороны почечки имеется небольшой складкообразный вырост —- эпибласт. В нижней части зародыша находится зародышевый корень, одетый корневым влагалищем, или колеоризой. Природа всех этих частей зародыша является предметом дискуссий. Щиток обычно принимается за единственную, видоизмененную семядолю, а колеоптиль —- за его вырост или за первый лист почечки. Эпибласт, когда он имеется, принимают или за складкообразный вырост колеоризы, или за рудимент второй семядоли. Колеориза, по мнению одних авторов, представляет собой нижнюю часть подсемядольного колена —- гипокотиля, в которой закладывается зародышевый корень, по мнению других —- видоизмененный главный корень зародыша.Особенности строения зародыша злаков имеют большое систематическое значение. На основании присутствия или отсутствия эпибласта или щели между нижней частью щитка и колеоризой, а также различий в ходе проводящих пучков зародыша и в форме первого листа зародыша на поперечном срезе были установлены 3 основных тина строения зародыша: фестукоидный, паникоидный и промежуточный между ними эрагростоидный (рис. 206, 3). Таким образом, и здесь были выявлены существенные анатомо-морфологические различия между преимущественно внетропическими, фестукоидными злаками и преимущественно тропическими, паникоидными и хлоридоидными злаками.Анатомо-морфологические особенности злаков определяют очень высокую пластичность и приспособляемость представителей этого семейства к самым различным экологическим условиям, что позволило им распространиться по всей суше земного шара .вплоть до самых крайних пределов существования цветковых растений. Злаки встречаются почти во всех растительных группировках, хотя наиболее характерны они для лугов, степей и саванн различных типов. Есть виды, обитающие на подвижных песках (селин —- Stipagrostis, песколюбка —- Ammophila и др.) и солончаках (особенно прибрежница —- Aeluropus и бескильница —- Puccinellia), как приморских, так и внутриконтинентальных. Некоторые виды бескильницы растут в полосе, затопляемой приливами, причем один арктический вид, приуроченный к таким местообитаниям,- бескильница ползучая (P. phryganodes) -часто не цветет, размножаясь с помощью стелющихся и укореняющихся в узлах вегетативных побегов. Для равнинных и нагорных лугов Евразии особенно характерны многочисленные виды родов мятлик, овсяница, полевица (Agrostis), вейник (Calamagrostis), лисохвост, кострец (Bromopsis), тимофеевка (Phleum), трясунка (Briza) и др. В степной зоне и в нагорных степях Евразии ведущее значение приобретают ковыль, овсяница-типчак, тонконог (Koeleria), житняк (Agropyron), овсец (Helictotrichon), а в более южных районах —- бородач (Bothriochloa). В прериях Северной Америки на первое место выдвигаются хлоридоидные злаки: бутелуа (Bouteloua), хлорис (Chloris), трава бизонов (Buchloe dactyloides) и др. В аридных районах Азии своеобразные растительные группировки —- чиевники —- образует крупнодерновинный злак чий блестящий (Achnatherum splendens). В пампасах Южной Америки большую роль играют виды пампасской травы. —- кортадерии (Cortaderia), образующие гигантские дерновины (табл. 45, 3, 4).В лесах роль злаков в растительном покрове, естественно, менее значительна, однако и здесь некоторые виды этого семейства могут доминировать в травянистом ярусе. Так, в еловых лесах Евразии нередко в изобилии разрастается вейник тростниковидный (Calamagrostis arundinacea), а в дубравах —- мятлик лесной (Роа nemoralis), элимус собачий (Elymus caninus), овсяница гигантская (Festuca gigantea) и другие виды. В отличие от степных злаков, обычно плотнодерновинных и имеющих очень узкие, вдоль сложенные листовые пластинки, лесные злаки имеют менее плотные дерновины, более широкие и менее жесткие листовые пластинки. Из двух распространенных в лиственных и смешанных лесах Евразии видов перловника более северный —- перловник поникший (Melica nutans) принадлежит к рыхлодерновинным злакам, а более южный и потому более ксерофильный перловник покрашенный (M. picta) —- к плотнодерновинным. Среди тропических и субтропических лесных злаков многие имеют лежачие или лазающие густооблиственные побеги и очень широкие, ланцетные или ланцетно-яйцевидные пластинки листьев, напоминая по внешнему облику широко распространенные в оранжерейной и комнатной культуре виды традесканции. Такую жизненную форму имеют, например, представители рода остянка (Oplismenus), один из видов которого —- остянка курчаволистная (O. undulatifolius) —- встречается во влажных лесах Средиземноморья, а также в Колхидской низменности (рис. 202, 1),. а другой —- остянка сложнометельчатая (O. compositus) —- очень обычен в лесах Южной Азии.Что касается злаков подсемейства бамбуковых, то их роль в растительности влажных тропиков и субтропиков довольно велика. Древовидные бамбуковые обычно образуют большие заросли по берегам водоемов, вдоль спускающихся с гор водотоков, на опушках и вырубках тропических лесов. Многие травянистые бамбуковые растут под пологом тропического дождевого леса и выносят значительную затененность. Надземные побеги древовидных бамбуковых нередко считают гомологичными корневищам других злаков. Они отличаются чрезвычайно быстрым ростом и по всей длине несут чешуевидные листья —- катафиллы, характерные для корневищ других злаков. Все древовидные бамбуки —- вечнозеленые растения, хотя листья их постепенно опадают в результате образования отделительной ткани или у основания черешков, или у основания влагалищ, которые в этом случае опадают вместе с пластинками.
Зародыш злаков (рис. 206) довольно сильно отличается по своему строению от зародышей других однодольных. На стороне, прилегающей к эндосперму, он имеет щитовидное тело —- щиток. Снаружи от него и ближе к его верхней части находится зародышевая почечка, одетая двукилевым влагалищеобразным листом —- колеоптилем. У многих злаков против щитка с наружной стороны почечки имеется небольшой складкообразный вырост —- эпибласт. В нижней части зародыша находится зародышевый корень, одетый корневым влагалищем, или колеоризой. Природа всех этих частей зародыша является предметом дискуссий. Щиток обычно принимается за единственную, видоизмененную семядолю, а колеоптиль —- за его вырост или за первый лист почечки. Эпибласт, когда он имеется, принимают или за складкообразный вырост колеоризы, или за рудимент второй семядоли. Колеориза, по мнению одних авторов, представляет собой нижнюю часть подсемядольного колена —- гипокотиля, в которой закладывается зародышевый корень, по мнению других —- видоизмененный главный корень зародыша.Особенности строения зародыша злаков имеют большое систематическое значение. На основании присутствия или отсутствия эпибласта или щели между нижней частью щитка и колеоризой, а также различий в ходе проводящих пучков зародыша и в форме первого листа зародыша на поперечном срезе были установлены 3 основных тина строения зародыша: фестукоидный, паникоидный и промежуточный между ними эрагростоидный (рис. 206, 3). Таким образом, и здесь были выявлены существенные анатомо-морфологические различия между преимущественно внетропическими, фестукоидными злаками и преимущественно тропическими, паникоидными и хлоридоидными злаками.Анатомо-морфологические особенности злаков определяют очень высокую пластичность и приспособляемость представителей этого семейства к самым различным экологическим условиям, что позволило им распространиться по всей суше земного шара .вплоть до самых крайних пределов существования цветковых растений. Злаки встречаются почти во всех растительных группировках, хотя наиболее характерны они для лугов, степей и саванн различных типов. Есть виды, обитающие на подвижных песках (селин —- Stipagrostis, песколюбка —- Ammophila и др.) и солончаках (особенно прибрежница —- Aeluropus и бескильница —- Puccinellia), как приморских, так и внутриконтинентальных. Некоторые виды бескильницы растут в полосе, затопляемой приливами, причем один арктический вид, приуроченный к таким местообитаниям,- бескильница ползучая (P. phryganodes) -часто не цветет, размножаясь с помощью стелющихся и укореняющихся в узлах вегетативных побегов. Для равнинных и нагорных лугов Евразии особенно характерны многочисленные виды родов мятлик, овсяница, полевица (Agrostis), вейник (Calamagrostis), лисохвост, кострец (Bromopsis), тимофеевка (Phleum), трясунка (Briza) и др. В степной зоне и в нагорных степях Евразии ведущее значение приобретают ковыль, овсяница-типчак, тонконог (Koeleria), житняк (Agropyron), овсец (Helictotrichon), а в более южных районах —- бородач (Bothriochloa). В прериях Северной Америки на первое место выдвигаются хлоридоидные злаки: бутелуа (Bouteloua), хлорис (Chloris), трава бизонов (Buchloe dactyloides) и др. В аридных районах Азии своеобразные растительные группировки —- чиевники —- образует крупнодерновинный злак чий блестящий (Achnatherum splendens). В пампасах Южной Америки большую роль играют виды пампасской травы. —- кортадерии (Cortaderia), образующие гигантские дерновины (табл. 45, 3, 4).В лесах роль злаков в растительном покрове, естественно, менее значительна, однако и здесь некоторые виды этого семейства могут доминировать в травянистом ярусе. Так, в еловых лесах Евразии нередко в изобилии разрастается вейник тростниковидный (Calamagrostis arundinacea), а в дубравах —- мятлик лесной (Роа nemoralis), элимус собачий (Elymus caninus), овсяница гигантская (Festuca gigantea) и другие виды. В отличие от степных злаков, обычно плотнодерновинных и имеющих очень узкие, вдоль сложенные листовые пластинки, лесные злаки имеют менее плотные дерновины, более широкие и менее жесткие листовые пластинки. Из двух распространенных в лиственных и смешанных лесах Евразии видов перловника более северный —- перловник поникший (Melica nutans) принадлежит к рыхлодерновинным злакам, а более южный и потому более ксерофильный перловник покрашенный (M. picta) —- к плотнодерновинным. Среди тропических и субтропических лесных злаков многие имеют лежачие или лазающие густооблиственные побеги и очень широкие, ланцетные или ланцетно-яйцевидные пластинки листьев, напоминая по внешнему облику широко распространенные в оранжерейной и комнатной культуре виды традесканции. Такую жизненную форму имеют, например, представители рода остянка (Oplismenus), один из видов которого —- остянка курчаволистная (O. undulatifolius) —- встречается во влажных лесах Средиземноморья, а также в Колхидской низменности (рис. 202, 1),. а другой —- остянка сложнометельчатая (O. compositus) —- очень обычен в лесах Южной Азии.Что касается злаков подсемейства бамбуковых, то их роль в растительности влажных тропиков и субтропиков довольно велика. Древовидные бамбуковые обычно образуют большие заросли по берегам водоемов, вдоль спускающихся с гор водотоков, на опушках и вырубках тропических лесов. Многие травянистые бамбуковые растут под пологом тропического дождевого леса и выносят значительную затененность. Надземные побеги древовидных бамбуковых нередко считают гомологичными корневищам других злаков. Они отличаются чрезвычайно быстрым ростом и по всей длине несут чешуевидные листья —- катафиллы, характерные для корневищ других злаков. Все древовидные бамбуки —- вечнозеленые растения, хотя листья их постепенно опадают в результате образования отделительной ткани или у основания черешков, или у основания влагалищ, которые в этом случае опадают вместе с пластинками.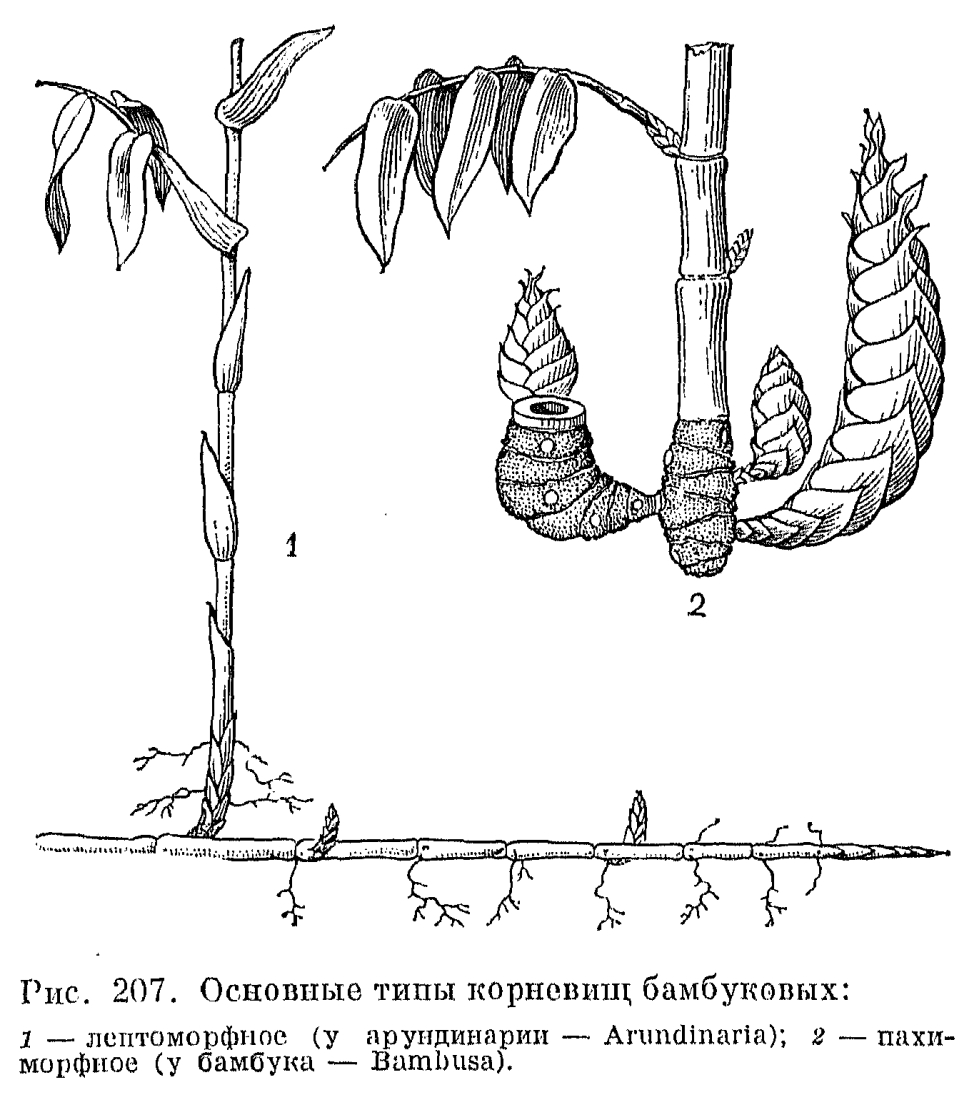 Среди бамбуковых с более или менее одревесневающими стеблями различают две основные жизненные формы, приуроченные к разным климатическим условиям (рис. 207). У большинства тропических бамбуковых, развитие которых в естественных условиях контролируется уровнем влажности (обычно наступлением дождливого сезона), стебли относительно сближены, формируя своего рода рыхлый куст. Такие бамбуковые имеют так называемые пахиморфные (от греч. «пахис» —- толстый) корневища: короткие и толстые, симподиальные, с заполненными сердцевиной асимметричными междоузлиями, ширина которых больше длины. Другая группа бамбуковых распространена в областях с относительно прохладной или даже холодной зимой, где начало активного роста их побегов контролируется температурными условиями. Принадлежащие к ней роды имеют лептоморфные (от греч. «лептос» —- тонкий) корневища: длинные и тонкие, моноподиальные, с полыми междоузлиями, длина которых значительно больше их ширины. Такие бамбуковые обычно имеют относительно небольшие общие размеры, хотя некоторые виды листоколосника бывают высотой до 10 и даже 15 м. Лептоморфные корневища имеет и единственный дикорастущий в СССР род бамбуковых —- саза (Sasa), образующий очень густые и труднопроходимые заросли по склонам гор на юге Сахалина и Курильских островов.Травянистые бамбуковые, подобно злакам других подсемейств, цветут ежегодно, но бамбуковые с одревесневающими стеблями, как правило, цветут один раз в 30-120 лет и после этого обычно погибают, являясь облигатными или факультативными монокарпиками. В 1969 г. дочти во всей Японии наблюдалось массовое и одновременное цветение очень широко культивируемого там в технических целях листоколосника бамбуковидного (Phyllostachys bambusoides). Это было настоящим бедствием для тех, кто его выращивал, так как значительная часть плантаций после цветения погибла. Почти весь японский листоколосник происходил от одного и того же клона, завезенного в Японию из Китая, и потому не удивительно, что он повсюду зацвел в одно и то же время.Среди многолетних травянистых злаков, особенно тропических, имеются гигантские формы, не уступающие по высоте многим бамбуковым. Таковы, например, тростник обыкновенный (Phragmites australis) и арундо тростниковый (Arundo donax), которые имеют многоузловые, но неразветвленные стебли высотой до 3, иногда до 5 м и длинные, сильно разветвленные корневища (рис. 208, 3).
Среди бамбуковых с более или менее одревесневающими стеблями различают две основные жизненные формы, приуроченные к разным климатическим условиям (рис. 207). У большинства тропических бамбуковых, развитие которых в естественных условиях контролируется уровнем влажности (обычно наступлением дождливого сезона), стебли относительно сближены, формируя своего рода рыхлый куст. Такие бамбуковые имеют так называемые пахиморфные (от греч. «пахис» —- толстый) корневища: короткие и толстые, симподиальные, с заполненными сердцевиной асимметричными междоузлиями, ширина которых больше длины. Другая группа бамбуковых распространена в областях с относительно прохладной или даже холодной зимой, где начало активного роста их побегов контролируется температурными условиями. Принадлежащие к ней роды имеют лептоморфные (от греч. «лептос» —- тонкий) корневища: длинные и тонкие, моноподиальные, с полыми междоузлиями, длина которых значительно больше их ширины. Такие бамбуковые обычно имеют относительно небольшие общие размеры, хотя некоторые виды листоколосника бывают высотой до 10 и даже 15 м. Лептоморфные корневища имеет и единственный дикорастущий в СССР род бамбуковых —- саза (Sasa), образующий очень густые и труднопроходимые заросли по склонам гор на юге Сахалина и Курильских островов.Травянистые бамбуковые, подобно злакам других подсемейств, цветут ежегодно, но бамбуковые с одревесневающими стеблями, как правило, цветут один раз в 30-120 лет и после этого обычно погибают, являясь облигатными или факультативными монокарпиками. В 1969 г. дочти во всей Японии наблюдалось массовое и одновременное цветение очень широко культивируемого там в технических целях листоколосника бамбуковидного (Phyllostachys bambusoides). Это было настоящим бедствием для тех, кто его выращивал, так как значительная часть плантаций после цветения погибла. Почти весь японский листоколосник происходил от одного и того же клона, завезенного в Японию из Китая, и потому не удивительно, что он повсюду зацвел в одно и то же время.Среди многолетних травянистых злаков, особенно тропических, имеются гигантские формы, не уступающие по высоте многим бамбуковым. Таковы, например, тростник обыкновенный (Phragmites australis) и арундо тростниковый (Arundo donax), которые имеют многоузловые, но неразветвленные стебли высотой до 3, иногда до 5 м и длинные, сильно разветвленные корневища (рис. 208, 3).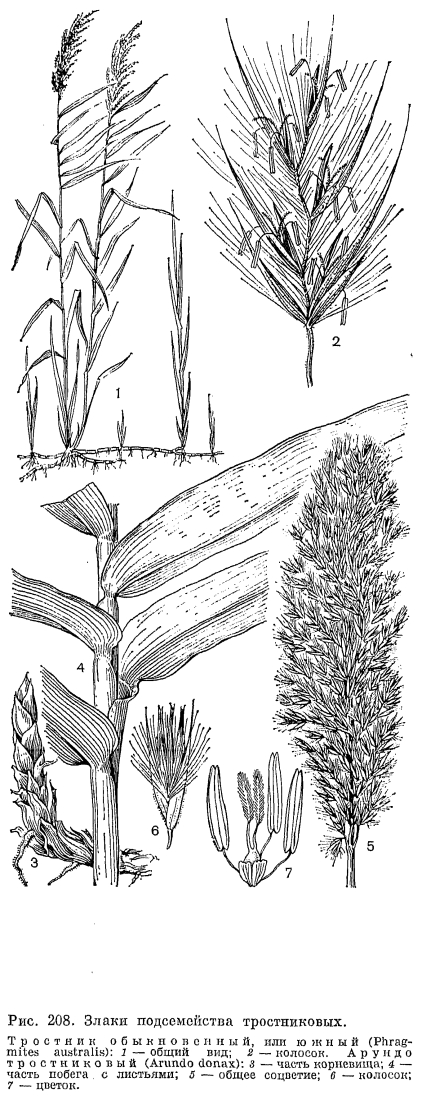 Тростники принадлежат к числу влаголюбивых растений, образующих большие и почти чистые заросли по берегам водоемов, а нередко и в воде. Тростник обыкновенный почти космополит и широко распространен на всех континентах как в тропиках, так и в умеренно теплых странах. Этот вид обладает довольно широкой экологической амплитудой. Он может расти также на болотах различных типов, в болотистых лесах, на горных склонах с подтоком грунтовых вод и на солончаках, образуя в крайних условиях существования своеобразную форму со стелющимися по земле и только вегетативными побегами. Впрочем, и у нормально развитых цветущих клонов тростника зерновки образуются далеко не всегда и в небольшом количестве, что, по-видимому, связано с большой древностью этого вида. Другой гигантский, высотой до 3 м, злак —- пампасская трава, или кортадерия, один из видов которой интродуцирован в страны Средиземноморья, образует очень густые дерновины с внутривлагалищными побегами (табл. 45, 3, 4). Ее узкие и очень жесткие листовые пластинки по краям и средней жилке несут крупные шипики, напоминая в этом отношении листья водного растения телореза (Stratiotes).Образование плотных дерновин особенно выгодно в условиях аридного климата, так как в этом случае основание растения хорошо защищено от перегревающегося верхнего слоя почвы. Именно поэтому среди степных и пустынных злаков так много плотнодерновинных (например, чий блестящий, многие виды ковыля и др.). Напротив, к длиннокорневищным относятся многие луговые злаки, особенно обитающие на рыхлых, слабо задерненных почвах, например пырей ползучий и кострец безостый (Bromopsis inermis), нередко в изобилии разрастающиеся на лугах прирусловых пойм, а также некоторые прибрежные виды, подобно тростнику образующие густые заросли, например виды манника (Glyceria), тростянки (Scolochloa), зизания широколистная, (Zizania latifolia) и др. Среди видов вообще гидрофильной трибы рисовых (Oryzeae) есть и настоящие водные растения. Такова, например, южноазиатская гигрориза остистая (Hygroryza aristata) с короткими и широкими листьями, собранными в розетки, плавающие на поверхности воды благодаря сильно вздутым влагалищам.Большую и очень интересную во многих отношениях группу жизненных форм образуют злаки-однолетники, которые могут быть как яровыми, когда прорастание семени начинается весной, так и озимыми, когда семена начинают прорастать осенью и молодые растения зимуют, продолжая свое развитие весной. У такого широко культивируемого хлебного растения, как пшеница, имеется не только много яровых и озимых сортов, но и сорта «двуручки», которые могут быть яровыми или озимыми в зависимости от сроков посева. Однолетние злаки могут быть разделены на 2 группы также по их происхождению. Одну из таких групп составляют весенние эфемеры. Быстро заканчивающие свой жизненный цикл в течение весны —- начале лета, они играют очень существенную роль в составе эфемеровой растительности в аридных и субаридных областях Евразии, Африки и Северной Америки. Очень важно, что от древнесредиземноморских эфемеров происходят такие ценные продовольственные и кормовые культуры, как пшеница, рожь, овес и ячмень.Другая большая группа однолетних злаков принадлежит к преимущественно тропическим трибам просовых, сорговых, свинороевых, триостренницевых и др., хотя некоторые виды этой группы (например, виды щетинника, полевички, росички —- Digitaria и ежовника) проникают далеко за пределы тропиков. Все это злаки относительно теплолюбивые и поздноразвивающиеся. Обычно они цветут во второй половине лета —- начале осени, будучи хорошо приспособленными к перенесению засушливого времени года. Среди поздних однолетников также имеется немало хозяйственно-ценных видов (сорго, просо, чумиза и др.), но много и злостных сорняков полей и плантаций различных культур.Среди однолетних злаков известны очень оригинальные по облику виды. Так, у двуколоски деуколосой (Trachynia distachya) общее соцветие состоит всего из 1-2 крупных многоцветковых колосков (рис. 201, 14); у ежовницы головчатой (Echinaria capitata) колоски собраны в почти шаровидную, колючую при плодах верхушечную головку (рис. 201, 11); у корнеголовника восточного (Rhizocephalus orientalis) и песочницы палестинской (Ammochloa palaestina) собранные в густую головку колоски расположены в центре листовых розеток (рис. 201, 1-7). У последнего вида, известного в СССР только с песков Апшеронского полуострова, часто почти все растение бывает засыпано песком, из которого видны только верхушки листьев розетки. Очень интересен в биологическом отношении поздний эфемер влагалищецветник маленький (Coleanthus subtilis), обитающий на береговых отмелях более или менее крупных рек. Он очень быстро развивается после выхода из-под воды отмелей, достигая полного развития в сентябре —- начале октября. Это маленькое растеньице, высотой 3-5 см, с лежачими или восходящими побегами и очень мелкими одноцветковыми колосками без колосковых чешуй, собранными зонтикообразными пучками (рис. 201, 5). В годы, когда отмели остаются залитыми водой, этот вид совсем не развивается и вообще может исчезать на многие годы. Распространен он во внетропических странах северного полушария, но крайне спорадично. Так, в СССР его находили лишь по верхнему течению Волхова, среднему течению Оби и по Амуру.Выше уже отмечалась высокая специализация цветков злаков к опылению с помощью ветра. Однако случайный перенос пыльцы злаков насекомыми даже у внетропических злаков не может считаться полностью исключенным. В последнее время установлено, что травянистые бамбуковые из родов олира (Olyra) и париана (Pariana), растущие под пологом деревьев в тропических дождевых лесах, где движение воздуха крайне незначительно, как правило, опыляются с помощью насекомых, главным образом мух и жуков, хотя такой вторичный переход к энтомофилии пока еще не связан с какими-либо специальными приспособлениями.Значительное большинство многолетних злаков опыляется перекрестно, причем самоопылению обычно препятствует полная или частичная самостерильность. Однако среди однолетников очень много факультативно самоопыляющихся видов. Таковы, например, все виды пшеницы и эгилопса (Aegilops), а также большинство видов костра (Bromus). Некоторые злаки, кроме обычных колосков с хазмогамными цветками, развивают еще колоски с клейстогамными цветками, опыляющимися при сомкнутых чешуях. Образование этих колосков гарантирует возможность семенного размножения при неблагоприятных погодных условиях или при чрезмерном обкусывании растения травоядными животными. Так, у широко распространенного прибрежного злака леерсии рисовидной (Leersia oryzoides) и североамериканского споробола скрытноцветкового (Sporobolus cryptandrus) в неблагоприятные годы образуются только колоски с клейстогамными цветками и метелки не выступают из расширенного влагалища верхнего листа. В метелках многих ковылей флоры СССР в засушливые годы образуются только клейстогамные цветки, а при более прохладной и влажной погоде все или почти все цветки метелки цветут открыто. Многие арктические злаки в условиях особенно холодной погоды также цветут в основном клейстогамно.
Тростники принадлежат к числу влаголюбивых растений, образующих большие и почти чистые заросли по берегам водоемов, а нередко и в воде. Тростник обыкновенный почти космополит и широко распространен на всех континентах как в тропиках, так и в умеренно теплых странах. Этот вид обладает довольно широкой экологической амплитудой. Он может расти также на болотах различных типов, в болотистых лесах, на горных склонах с подтоком грунтовых вод и на солончаках, образуя в крайних условиях существования своеобразную форму со стелющимися по земле и только вегетативными побегами. Впрочем, и у нормально развитых цветущих клонов тростника зерновки образуются далеко не всегда и в небольшом количестве, что, по-видимому, связано с большой древностью этого вида. Другой гигантский, высотой до 3 м, злак —- пампасская трава, или кортадерия, один из видов которой интродуцирован в страны Средиземноморья, образует очень густые дерновины с внутривлагалищными побегами (табл. 45, 3, 4). Ее узкие и очень жесткие листовые пластинки по краям и средней жилке несут крупные шипики, напоминая в этом отношении листья водного растения телореза (Stratiotes).Образование плотных дерновин особенно выгодно в условиях аридного климата, так как в этом случае основание растения хорошо защищено от перегревающегося верхнего слоя почвы. Именно поэтому среди степных и пустынных злаков так много плотнодерновинных (например, чий блестящий, многие виды ковыля и др.). Напротив, к длиннокорневищным относятся многие луговые злаки, особенно обитающие на рыхлых, слабо задерненных почвах, например пырей ползучий и кострец безостый (Bromopsis inermis), нередко в изобилии разрастающиеся на лугах прирусловых пойм, а также некоторые прибрежные виды, подобно тростнику образующие густые заросли, например виды манника (Glyceria), тростянки (Scolochloa), зизания широколистная, (Zizania latifolia) и др. Среди видов вообще гидрофильной трибы рисовых (Oryzeae) есть и настоящие водные растения. Такова, например, южноазиатская гигрориза остистая (Hygroryza aristata) с короткими и широкими листьями, собранными в розетки, плавающие на поверхности воды благодаря сильно вздутым влагалищам.Большую и очень интересную во многих отношениях группу жизненных форм образуют злаки-однолетники, которые могут быть как яровыми, когда прорастание семени начинается весной, так и озимыми, когда семена начинают прорастать осенью и молодые растения зимуют, продолжая свое развитие весной. У такого широко культивируемого хлебного растения, как пшеница, имеется не только много яровых и озимых сортов, но и сорта «двуручки», которые могут быть яровыми или озимыми в зависимости от сроков посева. Однолетние злаки могут быть разделены на 2 группы также по их происхождению. Одну из таких групп составляют весенние эфемеры. Быстро заканчивающие свой жизненный цикл в течение весны —- начале лета, они играют очень существенную роль в составе эфемеровой растительности в аридных и субаридных областях Евразии, Африки и Северной Америки. Очень важно, что от древнесредиземноморских эфемеров происходят такие ценные продовольственные и кормовые культуры, как пшеница, рожь, овес и ячмень.Другая большая группа однолетних злаков принадлежит к преимущественно тропическим трибам просовых, сорговых, свинороевых, триостренницевых и др., хотя некоторые виды этой группы (например, виды щетинника, полевички, росички —- Digitaria и ежовника) проникают далеко за пределы тропиков. Все это злаки относительно теплолюбивые и поздноразвивающиеся. Обычно они цветут во второй половине лета —- начале осени, будучи хорошо приспособленными к перенесению засушливого времени года. Среди поздних однолетников также имеется немало хозяйственно-ценных видов (сорго, просо, чумиза и др.), но много и злостных сорняков полей и плантаций различных культур.Среди однолетних злаков известны очень оригинальные по облику виды. Так, у двуколоски деуколосой (Trachynia distachya) общее соцветие состоит всего из 1-2 крупных многоцветковых колосков (рис. 201, 14); у ежовницы головчатой (Echinaria capitata) колоски собраны в почти шаровидную, колючую при плодах верхушечную головку (рис. 201, 11); у корнеголовника восточного (Rhizocephalus orientalis) и песочницы палестинской (Ammochloa palaestina) собранные в густую головку колоски расположены в центре листовых розеток (рис. 201, 1-7). У последнего вида, известного в СССР только с песков Апшеронского полуострова, часто почти все растение бывает засыпано песком, из которого видны только верхушки листьев розетки. Очень интересен в биологическом отношении поздний эфемер влагалищецветник маленький (Coleanthus subtilis), обитающий на береговых отмелях более или менее крупных рек. Он очень быстро развивается после выхода из-под воды отмелей, достигая полного развития в сентябре —- начале октября. Это маленькое растеньице, высотой 3-5 см, с лежачими или восходящими побегами и очень мелкими одноцветковыми колосками без колосковых чешуй, собранными зонтикообразными пучками (рис. 201, 5). В годы, когда отмели остаются залитыми водой, этот вид совсем не развивается и вообще может исчезать на многие годы. Распространен он во внетропических странах северного полушария, но крайне спорадично. Так, в СССР его находили лишь по верхнему течению Волхова, среднему течению Оби и по Амуру.Выше уже отмечалась высокая специализация цветков злаков к опылению с помощью ветра. Однако случайный перенос пыльцы злаков насекомыми даже у внетропических злаков не может считаться полностью исключенным. В последнее время установлено, что травянистые бамбуковые из родов олира (Olyra) и париана (Pariana), растущие под пологом деревьев в тропических дождевых лесах, где движение воздуха крайне незначительно, как правило, опыляются с помощью насекомых, главным образом мух и жуков, хотя такой вторичный переход к энтомофилии пока еще не связан с какими-либо специальными приспособлениями.Значительное большинство многолетних злаков опыляется перекрестно, причем самоопылению обычно препятствует полная или частичная самостерильность. Однако среди однолетников очень много факультативно самоопыляющихся видов. Таковы, например, все виды пшеницы и эгилопса (Aegilops), а также большинство видов костра (Bromus). Некоторые злаки, кроме обычных колосков с хазмогамными цветками, развивают еще колоски с клейстогамными цветками, опыляющимися при сомкнутых чешуях. Образование этих колосков гарантирует возможность семенного размножения при неблагоприятных погодных условиях или при чрезмерном обкусывании растения травоядными животными. Так, у широко распространенного прибрежного злака леерсии рисовидной (Leersia oryzoides) и североамериканского споробола скрытноцветкового (Sporobolus cryptandrus) в неблагоприятные годы образуются только колоски с клейстогамными цветками и метелки не выступают из расширенного влагалища верхнего листа. В метелках многих ковылей флоры СССР в засушливые годы образуются только клейстогамные цветки, а при более прохладной и влажной погоде все или почти все цветки метелки цветут открыто. Многие арктические злаки в условиях особенно холодной погоды также цветут в основном клейстогамно.
,У всех видов евроазиатского рода змеевка (Cleistogenes) и некоторых представителей других родов клейстогамные колоски постоянно образуются на коротких боковых веточках, скрытых во влагалищах верхних и средних стеблевых листьев (рис. 194, 2). Центрально-азиатский девятиостник северный (Enneapogon borealis) образует одиночные колоски с клейстогамными цветками внутри особых почкообразных побегов, расположенных при основании дерновины. Благодаря такой особенности этот вид получает возможность размножаться даже в условиях усиленного стравливания пастбищ, когда ежегодно все дерновины почти до основания обкусывает скот. При этом пасущийся скот разбивает ногами дерновины и разносит вместе с прилипшими к ним комочками земли зерновки девятиостника. Еще более высокая специализация в этом отношении отмечается у североамериканского амфикарпума (Amphicarpum). Его одиночные колоски с клейстогамными цветками образуются на верхушках ползучих подземных побегов под поверхностью почвы (рис. 202, 3).Однополые цветки встречаются у злаков нередко, но преимущественно у тропических видов. Эти цветки могут располагаться в одном и том же колоске вместе с обоеполыми цветками, например у зубровки (Hierochloe) из 3 цветков колоска верхний обоеполый, а 2 нижних мужские, но чаще находятся в разных колосках. Такие однополые колоски могут, в свою очередь, располагаться в одном и том же соцветии или в разных соцветиях. Как уже отмечалось выше, для многих родов трибы сорговых очень характерно расположение колосков на колосовидных веточках общего соцветия группами по 2: один сидячий с обоеполым цветком, другой на ножке —- с мужским цветком. Обоеполые, но с однополыми колосками соцветия южноамериканского травянистого бамбукового растения пирезии (Piresia) располагаются на стелющихся корневищеподобных побегах, одетых чешуевидными листьями, и нередко скрыты под подстилкой из опавших листьев. К сожалению, способ опыления цветков у видов этого рода пока остается неизвестным. В верхней части метелкообразных соцветий зизании располагаются более крупные колоски с женскими цветками, в нижней части —- более мелкие с мужскими цветками. У родственного кукурузе рода трипсакум (Tripsacum) колоски с женскими, цветками располагаются в нижней части колосовидных веточек метелки, а с мужскими —- в их верхней части (рис. 209, 6). У кукурузы колоски с мужскими цветками образуют верхушечное метелкообразное соцветие, а колоски с женскими цветками собраны продольными рядами на сильно утолщенной оси початков, расположенных в пазухах средних стеблевых листьев и окутанных влагалищеобразными листьями (рис. 209, 1-3). Еще более оригинально расположение однополых колосков у южноазиатского родственника кукурузы —- бусенника (Coix). Нижняя, женская часть колосовидных веточек, расположенных в пазухах верхних стеблевых листьев, состоит здесь из одного колоска с женским цветком и рудиментов двух других колосков, заключенных вместе в своеобразный ложный плод с очень плотной, роговидной или каменистой оболочкой. По происхождению этот плод —- видоизмененное влагалище верхушечного листа. Из верхней части его выходят длинные рыльцевые ветви женского цветка и ножка мужской части веточки, представляющей собой довольно густой ложный колос (рис. 210, 7).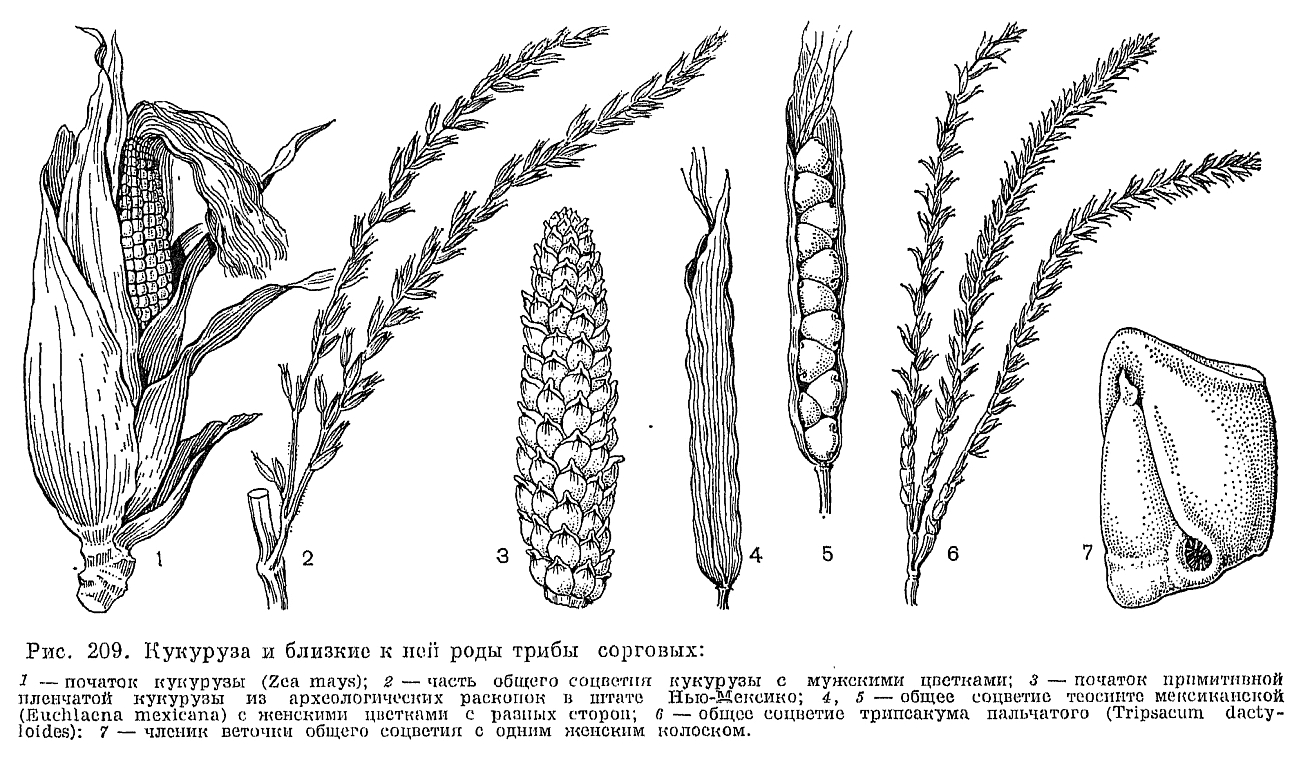
,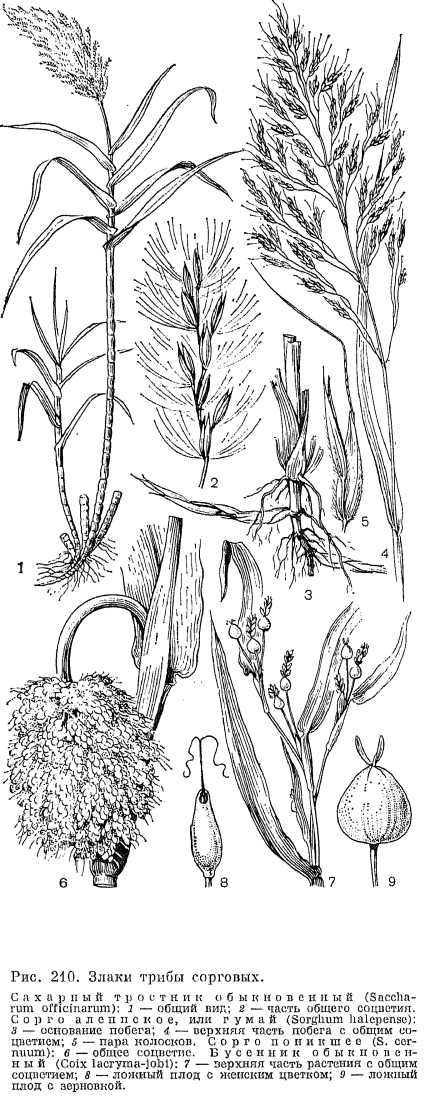 Примерами двудомных злаков могут служить культивируемая в садах и парках на юге СССР пампасская трава (Cortaderia selloana, табл.45, 3, 4) и трава бизонов (Buchloe dactyloides) из американских прерий, мужские и женские экземпляры которой были описаны сначала как виды разных родов (рис. 194, 6-9). Довольно широко представлены среди злаков различные способы бесполого размножения. В частности, вегетативное размножение с помощью ползучих корневищ, а также стелющихся и укореняющихся в узлах надземных побегов встречается у очень многих многолетних злаков. Преимущественно корневищами размножается, например, тростник обыкновенный, во внетропических странах лишь редко образующий нормально разнятые зерновки. Некоторые злаки-эфемероиды аридных областей Евразии, в том числе мятлик луковичный (Роа bulbosa) и катаброзелла низкая (Calabrosella humilis), имеют луковичкообразно утолщенные основания побегов дерновины. Позднее, в засушливое время года, их дерновины разбиваются травоядными животными, и луковички разносятся ветром или на ногах животных по пастбищу.
Примерами двудомных злаков могут служить культивируемая в садах и парках на юге СССР пампасская трава (Cortaderia selloana, табл.45, 3, 4) и трава бизонов (Buchloe dactyloides) из американских прерий, мужские и женские экземпляры которой были описаны сначала как виды разных родов (рис. 194, 6-9). Довольно широко представлены среди злаков различные способы бесполого размножения. В частности, вегетативное размножение с помощью ползучих корневищ, а также стелющихся и укореняющихся в узлах надземных побегов встречается у очень многих многолетних злаков. Преимущественно корневищами размножается, например, тростник обыкновенный, во внетропических странах лишь редко образующий нормально разнятые зерновки. Некоторые злаки-эфемероиды аридных областей Евразии, в том числе мятлик луковичный (Роа bulbosa) и катаброзелла низкая (Calabrosella humilis), имеют луковичкообразно утолщенные основания побегов дерновины. Позднее, в засушливое время года, их дерновины разбиваются травоядными животными, и луковички разносятся ветром или на ногах животных по пастбищу.
,Не менее часто встречается у злаков и бесполое размножение с помощью тех частей или органов растения, которые имеют отношение к половому размножению. Сюда принадлежит вивипария, когда молодое растение развивается не из семени, а из видоизмененных в луковичкообразные почки колосков. Полное или почти полное превращение всех колосков метелки в такие почки встречается у целого ряда арктических злаков из родов мятлик, овсяница, щучка, а также у широко распространенного в аридных областях Евразии мятлика луковичного. Во всех случаях вивипарию можно рассматривать как приспособление к более суровым условиям обитания, хотя вивипарные виды и разновидности могут возникать и в результате гибридизации между видами.Случаи апомиксиса в узком понимании этого термина или агамоспермии, когда молодое растение развивается из семени, но без предшествовавшего его образованию слияния гамет, еще более часты, особенно в преимущественно тропических трибах просовых и сорговых. Из внетропических злаков много апомиктических и полуапомиктических видов в родах мятлик и вейник.Для злаков, высокоспециализированных анемофильных растений, особое значение приобретает суточная ритмика цветения и опыления. Точное совмещение цветения всех особей данного вида в течение какого-либо ограниченного времени суток существенно повышает шансы перекрестного опыления и является важным приспособлением ко все более совершенной анемофилии. Среди внетропических злаков выделяют несколько групп видов, различающихся по времени цветения: с одноразовым утренним цветением (наиболее многочисленная группа), с одноразовым полуденным или послеполуденным цветением, с двухразовым, утренним и вечерним цветением (вечернее более слабое), с круглосуточным цветением, с ночным цветением. Последнее встречается лишь у немногих внетропических злаков. Однако в жарких и сухих районах тропиков ночное цветение известно у многих видов, так как оно позволяет избежать перегрева и быстрой гибели пыльцы во время жаркого дня. Интересно, что у тропических злаков с ночным цветением при выходе за пределы тропиков цветение смещается на раннее утро, так как опасность перегрева пыльцы уменьшается. У злаков, цветущих в полдень и после полудня, цветение приходится на самое жаркое время суток. Пыльцевые зерна в это время относительно быстро сморщиваются и погибают, однако таким злакам особенно часто свойственно так называемое взрывчатое цветение, при котором массовое и одновременное раскрывание цветков происходит за очень короткое время —- не более 3-5 мин. При порционном цветении, также свойственном многим злакам, в течение дня происходит не один, а несколько таких взрывов цветения. Было показано, что даже очень близкие виды, например степные овсяницы: валлисская (Festuca valosiaca) и ложноовечья (F. pseudovina), при совместном: обитании могут быть генетически вполне изолированными друг от друга, ибо цветут в разное время суток. Таким образом, определенная суточная ритмика цветения у злаков оказалась хорошим видовым систематическим признаком.Единицей распространения плодов —- диаспорой —- у злаков обычно бывает антеций: зерновка, заключенная в цветковые чешуи с прилегающим к ним члеником оси колоска. Значительно реже диаспорами служат голые (лишенные всяких чешуи) зерновки, целые колоски, части общего соцветия, все общее соцветие или даже все растение. У упомянутого выше влагалищецветника маленького сильно выступающие из цветковых чешуи зерновки выпадают из них и разносятся водой при колебаниях уровня рек, связанных с паводками, дождями, изменением направления ветра и т. п. Псаммофильный эфемер песочница палестинская может служить редким примером, когда выпадающие из колосков зерновки разносятся с помощью ветра. У широко распространенного в тропиках споробола (Sporobolus) мешочкообразные зерновки при смачивании их дождем или росой быстро набухают, лопаются, и выдавленные из них семена, окруженные клейкой слизью, свешиваются из колосков, прилипая к шерсти животных и перьям птиц. Выпадающие из колосков крупные зерновки многих бамбуковых распространяются преимущественно водными потоками во время тропических ливней, а также с помощью птиц. Ягодообразные зерновки мелоканны начинают прорастать еще на материнском растении, не имея периода покоя, затем падают на влажную почву острым концом вниз и продолжают свое развитие уже самостоятельно. Могут распространяться они также с помощью поедающих их птиц и зверей.Распространение с помощью целых общих соцветий или их частей встречается у злаков также не очень редко. Колосовидные метелки щетинника мутовчатого (Setaria verticillata), очень цепкие благодаря присутствию на окружающих колоски щетинках направленных назад шипиков, часто прицепляются к шерсти животных или одежде человека вместе со стеблями. Колосья многих видов эгилопса (Aegilops) с крупными, оттопыренными в сторону остями легко запутываются в шерсти животных, но могут переноситься на большие расстояния и ветром. Группы колосков ячменя гривастого (Hordeum jubatum), несущие очень длинные и тонкие ости, также могут переноситься как с помощью животных, так и ветром. В последнем случае многочисленные группы колосков могут сцепляться вместе, образуя шарообразное перекати-поле, переносимое ветром на большие расстояния, особенно вдоль шоссейных дорог. Многие другие злаки распространяются ветром по типу перекати-поле, причем основу последнего составляют очень крупные, широко и растопыренно разветвленные метелки. Примерами такого рода могут служить сибирский мятлик широкометельчатый (Роа subfastigiata) или нижневолжская цингерия Биберштейна (Zingeria biebersteinii). У литорального азиатского и австралийского рода спинифекс (Spinifex, рис. 211, 3) женские общие соцветия, имеющие почти шаровидную форму, опадают целиком, затем перекатываются ветром по песчаному побережью или плавают в воде и, уже задержавшись где-нибудь, постепенно распадаются. Очень любопытен и способ распространения змеевки растопыренной (Cleistogenes squarrosa) —- одного из характерных растений степей и пустынь Евразии (рис. 194, 2). Стебли этого вида при плодах змеевидно изгибаются и обламываются у своего основания. Сцепляясь друг с другом, они образуют легко переносимое ветром перекати-поле, причем зерновки постепенно выпадают не только из верхушечной метелки, но и из пазух стеблевых листьев, где находятся укороченные веточки с клейстогамными колосками.
,У злаков почти одинаково представлено распространение диаспор с помощью ветра и животных, а во многих случаях диаспоры могут распространяться обоими путями (например, у обычного в степях Евразии ковыля-тырсы —- Stipa capillata). По-видимому, во многих группах злаков в ходе эволюции шел переход от преимущественно зоохорного способа распространения к преимущественно анемохорному. Так, в роде вейник диаспоры более древних, лесных видов (вейника тростниковидного и др.) имеют длинные коленчато согнутые ости и пучок коротких жестких волосков на каллусе —- приспособление к зоохории, а диаспоры относительно более молодого вида вейника наземного (Саlamagrostis epigeios) снабжены очень короткой остью и пучком очень длинных (длиннее цветковых чешуи) волосков на каллусе, распространяясь исключительно анемохорно. Виды нередко объединяемого с ковылем, но более примитивного рода чий (Achnatherum) также имеют небольшие зоохорно распространяющиеся диаспоры, в то время как среди ковылей известны высокоспециализированные анемохорные виды с очень длинными (40 см и более), дважды коленчато согнутыми и перистоволосистыми в верхней части остями. Длинный и острый каллус с направленными вверх жесткими волосками дает возможность диаспорам ковылей как бы ввинчиваться в почву. При этом верхняя, горизонтально расположенная часть ости закрепляется среди других растений, а ее нижняя, скрученная часть обладает гигроскопичностью и при изменениях влажности то скручивается, то раскручивается, продвигая цветковые чешуи с зерновкой все глубже и глубже в почву. У некоторых ковылей, способных распространяться на шерсти животных, например у ковыля-тырсы, диаспоры могут ввинчиваться в их кожу, причиняя животным серьезный ущерб.Увеличение парусности диаспор у анемохорных злаков особенно часто осуществляется за счет длинных волосков, которые могут располагаться по бокам нижней цветковой чешуи (у перловника транссильванского —- Melica transsilvanica), на сильно удлиненном каллусе нижней цветковой чешуи (у тростника), на членике оси колоска над основанием цветковых чешуи (у многих видов вейника), на сильно удлиненных остях (у многих ковылей). У распространенного в песчаных пустынях Евразии селина перистого (Stipagrostis pennata) ость делится на 3 перистоволосистые ветви, напоминая по своему облику парашют. У многих видов хлориса парашютное устройство выглядит как поперечный ряд длинных волосков в верхней части нижних цветковых чешуи, а у девятиостника персидского (Enneapogon persicus) —- как поперечный ряд из 9 перистоволосистых остей. Легко переносятся ветром толстые, но очень легкие членики колосьев псаммофильных родов —- двучешуйника (Parapholis) и одночешуйницы (Monerma). Парусность диаспор, состоящих из целого колоска, может увеличиваться, за счет крылатых колосковых чешуй (у канареечника —- Phalaris) или за счет их мешковидного вздутия (у бекманнии —- Beckmannia). У трясунки (Briza) парусность диаспор-антециев увеличивается благодаря сильно расширенным и почти целиком перепончатым нижним цветковым чешуям.
,Приспособления злаков к зоохории не менее разнообразны. Особенно часто их диаспоры-антеции имеют коленчато согнутые шероховатые ости и жесткие волоски на каллусе, однако у представителей рода козлец (Tragus) и некоторых других родов на спинке нижних цветковых чешуи рядами располагаются крючковатые шипы. У травянистого бамбукового лептасписа улитковидного (Leptaspis cochleata) опадающие вместе с зерновкой замкнутые и вздутые нижние цветковые чешуи покрыты мелкими крючковидно загнутыми на верхушке шипиками и легко прикрепляются к шерсти животных (рис. 197, 4). У колючещетинника (Cenchrus) экзозоохорно распространяются довольно крупные колючие головки, состоящие из нескольких колосков, заключенных в обертку из расширенных и сросшихся в нижней части щетинок —- видоизмененных веточек общего соцветия (рис. 202, 8-9). Плодоносящие колоски тропического рода лазиацис (Lasiacis) распространяются птицами, которых привлекают богатые маслами утолщенные колосковые чешуи. Диаспоры многих видов перловника (Melica) имеют на верхушке оси колоска сочные придатки из недоразвитых цветковых чешуй и распространяются с помощью поедающих эти придатки муравьев. Диаспоры многих водных и прибрежных злаков (например, зизании, манника и др.) обладают хорошей плавучестью и легко разносятся потоками воды, а некоторых других видов (например, овсов-овсюгов, рис. 212) способны к самостоятельному передвижению (автохории) за счет гигроскопического скручивания или раскручивания остей. В настоящее время чрезвычайно возросла как сознательная, так и бессознательная роль человека в распространении злаков. Значительно расширяются ареалы культивируемых видов нередко вместе со специфическими для них сорняками. Вводятся в культуру в качестве кормовых растений, а затем дичают многие злаки с других континентов (например, в СССР широко распространился пырей бескорневищный или элимус новоанглийский —- Elymus novae-angliae, интродуцированный из Северной Америки). Многие уже давно введенные в культуру виды злаков потеряли свойственный их предкам способ распространения. Так, у культивируемых видов пшеницы, ржи, ячменя колосья не распадаются на членики; у культивируемого овса нет сочленений на оси колоска; у чумизы и могара (Setaria italica) нет сочленений у основания колосков, характерных для дикорастущих представителей этого рода. Только в культуре известны неспособные размножаться без помощи человека такие злаки, как кукуруза и бусенник.При прорастании зерновки прежде всего начинает расти зародышевой корешок, а затем почечка зародыша, прикрытая колеоптилем. После выхода колеоптиля на поверхность почвы из него выступает первый лист проростка, который продолжает быстро удлиняться и принимает характерную для данного вида форму. У злаков различают 2 основных типа проростков: фестукоидный, когда первый лист проростка узкий и почти вертикально вверх направленный (он встречается у фестукоидных триб злаков), и паникоидный, когда первый лист проростка широкий (ланцетный или ланцетно-яйцевидный) и почти горизонтально отклоненный от оси побега (он известен у паникоидных триб). Кроме того, встречается промежуточный между ними эрагростоидный тип, а в последнее время выделены еще 2 типа —- бамбузоидный и оризоидный, у которых на оси проростка вслед за колеоптилем следуют не обычные листья, а один или несколько катафиллов —- чешуевидных листьев, причем при бамбузоидном типе, свойственном подсемейству бамбуковых, первый вполне развитый лист проростка построен по паникоидному типу, а при оризоидном типе, характерном для подсемейства рисовых, он более близок к фестукоидному типу.Первоначальные варианты системы злаков основывались главным: образом на легко бросающихся в глаза признаках в строении общих соцветий и колосков. Долгое время общепринятой была система известного специалиста по злакам —- Э. Гаккеля (1887). Эта система была построена по принципу постепенного усложнения в строении колосков, от триб сорговых и просовых, обычно имеющих колоски с одним развитым цветком, и до бамбуковых, многие из которых имеют многоцветковые колоски очень примитивного строения. Однако уже в начале XX в. накопилось много новых данных по анатомии листьев и стеблей, строению зародыша и проростков, мелким деталям в строении цветков, строению крахмальных зерен, которые позволили коренным образом пересмотреть систему Гаккеля. Стало ясно, что основным направлением эволюции генеративных органов злаков было не их усложнение, а, напротив, упрощение: уменьшение количества цветков в колоске, цветковых пленок, тычинок и рыльцевых ветвей.Важные данные для построения новой системы дало также изучение хромосом злаков, связанное с бурным развитием генетики. В классической работе Н. П. Авдулова, вышедшей в 1931 г., было установлено, что величина хромосом и их основное число (х) в семействе злаков являются признаками не только постоянными пределах большинства родов, но и характерными для более крупных подразделений этого семейства. Относительно мелкие хромосомы при основном числе, равном 6, 9 и 10, оказались свойственными преимущественно тропическим трибам злаков (сорговым, просовым, свинороевым и др.), а более крупные хромосомы при основном числе 7 —- преимущественно внетропическим трибам мятликовых, овсовых, пшеницевых и др. В предложенной Авдуловым системе злаки были разделены на 2 подсемейства —- сахарнотростниковые (Sacchariflorae) и мятликовые (Poatae). Последнее подсемейство, в свою очередь, делилось на 2 серии: тростниковые (Phragmitiformis) с более древними трибами, имеющими мелкие хромосомы, и овсяницевые (Festuciformis) с большинством внетропических триб злаков, имеющих крупные хромосомы обычно в числе, кратном 7.Система Авдулова стала основой для последующих систем злаков, в которых первое место заняло подсемейство бамбуковые (Bainbusoideае). На основании упомянутых выше признаков были выделены еще 5 подсемейств, одно из которых —- рисовые (Oryzoideae) —- занимает как бы промежуточное положение между бамбуковыми и другими злаками, а остальные 4 —- мятликовые (Pooideae), тростниковые (Arundinoideae), полевичковые (Eragrostoideae) и просовые (Panicoideae) —- образуют постепенный переход от полного набора фестукоидных признаков, характерных для внетропических злаков, к полному набору паникоидных признаков, характерных для тропических злаков. Следует отметить, что различия между 4 последними подсемействами оказались не так уж выдержанными, как это казалось сначала, вследствие чего они признаются не всеми авторами. Так, среди просовых оказался целый ряд видов (в том числе в роде просо) с фестукоидной анатомией листьев (и, следовательно, без кранц-синдрома). Среди мятликовых, для которых характерны относительно крупные хромосомы с основным числом 7, имеются роды с мелкими хромосомами (например, коротконожка —- Brachypodium) и роды с основным числом хромосом 6 (канареечник —- Phalaris), 9 (перловник) и 10 (манник). В последнее время у двух фестукоидных злаков —- цингерии Биберштейна (Zingeria biebersteinii) и колподиума разноцветного (Colpodium versicolor) обнаружено наименьшее у высших растений общее число хромосом (2n = 4) при основном хромосомном числе 2. Ранее такое число было известно только у одного американского вида из семейства сложноцветных. Даже в пределах одного и того же фестукоидного вида, средиземноморского эфемера бора весеннего (Milium vernale), выявлены расы с основными числами хромосом 5, 7 и 9.
Диаспоры многих водных и прибрежных злаков (например, зизании, манника и др.) обладают хорошей плавучестью и легко разносятся потоками воды, а некоторых других видов (например, овсов-овсюгов, рис. 212) способны к самостоятельному передвижению (автохории) за счет гигроскопического скручивания или раскручивания остей. В настоящее время чрезвычайно возросла как сознательная, так и бессознательная роль человека в распространении злаков. Значительно расширяются ареалы культивируемых видов нередко вместе со специфическими для них сорняками. Вводятся в культуру в качестве кормовых растений, а затем дичают многие злаки с других континентов (например, в СССР широко распространился пырей бескорневищный или элимус новоанглийский —- Elymus novae-angliae, интродуцированный из Северной Америки). Многие уже давно введенные в культуру виды злаков потеряли свойственный их предкам способ распространения. Так, у культивируемых видов пшеницы, ржи, ячменя колосья не распадаются на членики; у культивируемого овса нет сочленений на оси колоска; у чумизы и могара (Setaria italica) нет сочленений у основания колосков, характерных для дикорастущих представителей этого рода. Только в культуре известны неспособные размножаться без помощи человека такие злаки, как кукуруза и бусенник.При прорастании зерновки прежде всего начинает расти зародышевой корешок, а затем почечка зародыша, прикрытая колеоптилем. После выхода колеоптиля на поверхность почвы из него выступает первый лист проростка, который продолжает быстро удлиняться и принимает характерную для данного вида форму. У злаков различают 2 основных типа проростков: фестукоидный, когда первый лист проростка узкий и почти вертикально вверх направленный (он встречается у фестукоидных триб злаков), и паникоидный, когда первый лист проростка широкий (ланцетный или ланцетно-яйцевидный) и почти горизонтально отклоненный от оси побега (он известен у паникоидных триб). Кроме того, встречается промежуточный между ними эрагростоидный тип, а в последнее время выделены еще 2 типа —- бамбузоидный и оризоидный, у которых на оси проростка вслед за колеоптилем следуют не обычные листья, а один или несколько катафиллов —- чешуевидных листьев, причем при бамбузоидном типе, свойственном подсемейству бамбуковых, первый вполне развитый лист проростка построен по паникоидному типу, а при оризоидном типе, характерном для подсемейства рисовых, он более близок к фестукоидному типу.Первоначальные варианты системы злаков основывались главным: образом на легко бросающихся в глаза признаках в строении общих соцветий и колосков. Долгое время общепринятой была система известного специалиста по злакам —- Э. Гаккеля (1887). Эта система была построена по принципу постепенного усложнения в строении колосков, от триб сорговых и просовых, обычно имеющих колоски с одним развитым цветком, и до бамбуковых, многие из которых имеют многоцветковые колоски очень примитивного строения. Однако уже в начале XX в. накопилось много новых данных по анатомии листьев и стеблей, строению зародыша и проростков, мелким деталям в строении цветков, строению крахмальных зерен, которые позволили коренным образом пересмотреть систему Гаккеля. Стало ясно, что основным направлением эволюции генеративных органов злаков было не их усложнение, а, напротив, упрощение: уменьшение количества цветков в колоске, цветковых пленок, тычинок и рыльцевых ветвей.Важные данные для построения новой системы дало также изучение хромосом злаков, связанное с бурным развитием генетики. В классической работе Н. П. Авдулова, вышедшей в 1931 г., было установлено, что величина хромосом и их основное число (х) в семействе злаков являются признаками не только постоянными пределах большинства родов, но и характерными для более крупных подразделений этого семейства. Относительно мелкие хромосомы при основном числе, равном 6, 9 и 10, оказались свойственными преимущественно тропическим трибам злаков (сорговым, просовым, свинороевым и др.), а более крупные хромосомы при основном числе 7 —- преимущественно внетропическим трибам мятликовых, овсовых, пшеницевых и др. В предложенной Авдуловым системе злаки были разделены на 2 подсемейства —- сахарнотростниковые (Sacchariflorae) и мятликовые (Poatae). Последнее подсемейство, в свою очередь, делилось на 2 серии: тростниковые (Phragmitiformis) с более древними трибами, имеющими мелкие хромосомы, и овсяницевые (Festuciformis) с большинством внетропических триб злаков, имеющих крупные хромосомы обычно в числе, кратном 7.Система Авдулова стала основой для последующих систем злаков, в которых первое место заняло подсемейство бамбуковые (Bainbusoideае). На основании упомянутых выше признаков были выделены еще 5 подсемейств, одно из которых —- рисовые (Oryzoideae) —- занимает как бы промежуточное положение между бамбуковыми и другими злаками, а остальные 4 —- мятликовые (Pooideae), тростниковые (Arundinoideae), полевичковые (Eragrostoideae) и просовые (Panicoideae) —- образуют постепенный переход от полного набора фестукоидных признаков, характерных для внетропических злаков, к полному набору паникоидных признаков, характерных для тропических злаков. Следует отметить, что различия между 4 последними подсемействами оказались не так уж выдержанными, как это казалось сначала, вследствие чего они признаются не всеми авторами. Так, среди просовых оказался целый ряд видов (в том числе в роде просо) с фестукоидной анатомией листьев (и, следовательно, без кранц-синдрома). Среди мятликовых, для которых характерны относительно крупные хромосомы с основным числом 7, имеются роды с мелкими хромосомами (например, коротконожка —- Brachypodium) и роды с основным числом хромосом 6 (канареечник —- Phalaris), 9 (перловник) и 10 (манник). В последнее время у двух фестукоидных злаков —- цингерии Биберштейна (Zingeria biebersteinii) и колподиума разноцветного (Colpodium versicolor) обнаружено наименьшее у высших растений общее число хромосом (2n = 4) при основном хромосомном числе 2. Ранее такое число было известно только у одного американского вида из семейства сложноцветных. Даже в пределах одного и того же фестукоидного вида, средиземноморского эфемера бора весеннего (Milium vernale), выявлены расы с основными числами хромосом 5, 7 и 9.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.
.