- Подсемейство каркасовые (Celtidoideae)
- Подсемейство каркасовые (Celtidoideae)
-
Подсемейство каркасовых — вечнозеленые, полулистопадные или листопадные деревья, реже вечнозеленые лазящие лианы, распространенные в тропиках и субтропиках всех частей света. Из 9 составляющих подсемейство родов (около 80 видов) только один монотипный род птероцельтис (Pteroceltis) но выходит за пределы, теплоумеренной зоны, остальные 8 родов включают преимущественно тропические растения, и лишь отдельные их виды произрастают в районах умеренно теплого климата.По большинству морфологических признаков и общему уровню развития каркасовые являются значительно более специализированной группой, чем ильмовые.В подсемейство преобладают виды с однополыми цветками, хотя у некоторых обычен небольшой процент обоеполых цветков. В тепло-умеренной зоне каркасовые представлены только однодомными растениями, в тропиках — и однодомными и двудомными, причем даже в пределах одного вида можно видеть разную степень дифференцировки деревьев по преобладанию женских или мужских цветков. Хетакма остистая (Chaetacmo aristata), например, в тропиках Восточной Африки почти всегда имеет двудомные цветки, а в Южной Африке чаще развивается как однодомное растение. Все переходы от однодомности к двудомности отмечены у тремы восточной (Trema orientalis), тремы Ламарка (Т. lamarckiana), каркаса Дюрана (Celtis durandii) и др.Мужские цветки каркасовых собраны в многоцветковые соцветия в пазухах чешуевидных листьев, женские расположены выше по побегу в пазухах золеных листьев, по 1—3, или в сложных многоцветковых соцветиях.В отличие от подсемейства ильмовых, характеризующихся сухими плодами, все каркасовые имеют один тип плода — костянку (табл.36), но строение ее, размеры и форма разнообразны (рис. 132).

,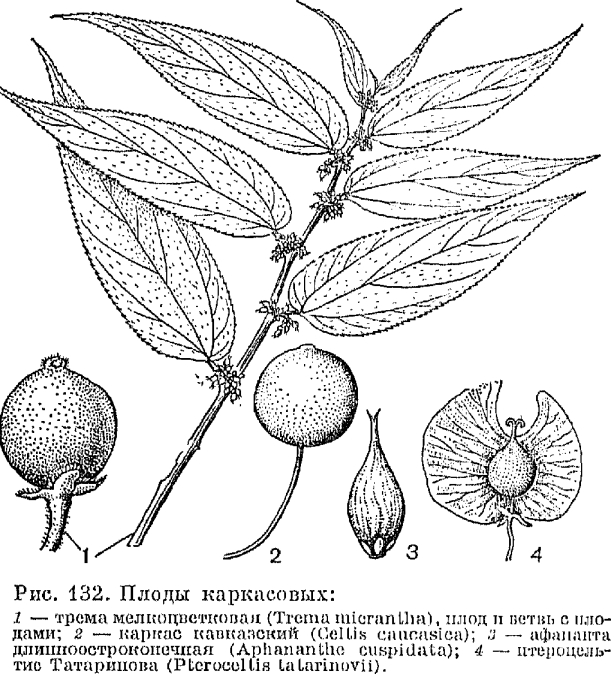 Семена каркасовых обычно округлые, зародыш поперек согнут, поперек сложен или свернут в спираль; в зрелых семенах эндосперм сохраняется, он окружает зародыш и заполняет углубления его складок; семенная кожура всегда однослойная, околоплодник 3—4-слойный.Основное хромосомное число у каркасовых такое же, как у коноплевых (х —10). Характерна полиплоидия, причем отдельные виды достигают очень высокой степени плоидности (трема амбоинская — Trema amboinensis — 16-плоид!).Листья у большинства каркасовых имеют 3 четко выраженные базальные жилки, что приближает их жилкование к пальчатому типу. У большинства каркасов Южной Америки на нижней стороне листовой пластинки в результате разрастания оснований этих жилок (вероятно, под воздействием поселяющихся в них клещиков) развиваются специфические структуры, похожие на вздутые кармашки. Наконец, листьям каркасовых свойственны округлые цистолиты и многообразный характер опушения (у вест-индской тремы Ламарка на одном листе развивается до 4 типов волосков, включая и железистые).Основными родами подсемейства каркасовых являются 2 пантропических рода: каркас (Celtis, табл. 36) и трема (Trema), включающие более 85% всех видов. Каркас — не только самый крупный (более 50 видов), наиболее полиморфный, но и наиболее широко распространенный род. Его ареал опоясывает земной шар огромной полосой, северная граница которой колеблется около 40° северной широты, проходя по Японии, континентальной Азии, Кавказу, Южной Европе и Северной Америке; южная идет примерно по 35o южной широты через Новую Каледонию, Восточную Австралию, Капскую область Африки и Южную Аргентину. Несмотря на большую полиморфность, каркасы сохраняют в пределах всего ареала единый тип строения цветков, плодов и листьев. Виды каркаса или диплоидные (число хромосом в соматических клетках 2n = 20), или тетраплоидные (2 n = 40) растения. К последним относятся все виды СССР, распространенные также в Средиземноморье: каркас кавказский (С. caucasica), каркас голый (С. glabrata), каркас южный (С. australis), каркас Турнефора (С. tournefortii). Это листопадные, летнезеленые деревья с гладкой светло-серой корой и раскидистой кроной, достигающие в высоту иногда 30 м, со стволом диаметром до 3 м, или растущие небольшими деревцами и приобретающие кустарниковидную форму в неблагоприятных условиях среды (каркас голый и каркас Турнефора).Виды каркаса занимают различные экологические ниши и растут в разнообразных сообществах. Каркас кавказский и каркас Турнефора чаще распространены в сухих лесах и аридных редколесьях, на низменностях и в горах, нередко поднимаясь до высоты 2500—2800 м над уровнем моря, обычно в местах, удаленных от моря. Напротив, каркас южный и каркас голый растут преимущественно в приморских районах. Все эти виды обитают на открытых каменистых склонах и в ущельях, среди скал, на осыпях, по каменистым берегам небольших рек или по склонам к морю.Богато представлены и разнообразны каркасы тропических стран, где наряду с листопадными и полулистопадными обычны и вечнозеленые растения, причем у многих из них почечные чешуи не развиваются и зачатки формирующегося побега прикрыты лишь прилистниками кроющего листа.Очень своеобразны каркасы Австралии и особенно Новой Каледонии, отличающиеся цельно-крайними толстыми суккулентными листьями. У каркаса скученного (С. conferta), произрастающего в прибрежной зоне и нередкого среди мангровых зарослей, листья собраны на верхушке побега и иногда супротивны. В тропиках Азии, Африки и Южной Америки каркасы входят в сообщества вечнозеленых дождевых лесов низменностей (каркас Уайта — С. wightii, каркас Мильдбреда — С. mildbraedii), а также горных лесов (каркас Дюрана и др.). Растут тропические каркасы и в сухих вечнозеленых лесах, нередко образуя листопадные редколесья (каркас африканский — С. africana, каркас цельнолистный — С. integrifolia).В нарушенных рубкой лесах тропиков Нового Света интенсивно развиваются вечнозеленые лазящие лианы: каркас игуановый (С. iguanaea), каркас колючий (С. spinosa), каркас боливийский (С. boliviensis). В первые годы жизни эти каркасы растут как прямостоячие деревца или кустарники (высотой 1,5—5 м), позднее их верхние ветви интенсивно удлиняются и, цепляясь загнутыми колючками за близ стоящие деревья и кустарники, используют их как опору. Растение таким образом превращается в лазящую лиану и сохраняет этот облик до конца жизни.В теплоумеренном климате каркасы цветут весной, почти одновременно с распусканием листьев. Их мужские цветки раскрываются на несколько дней раньше обоеполых и женских. Цветки опыляются ветром, и, хотя их посещают насекомые, энтомофилия не имеет большого значения, так как пыльца лишена характерных признаков зерен энтомофильных растений и в момент выпрямления тычиночной нити мгновенно высыпается в окружающее пространство.Плоды созревают осенью, к этому времени внутренняя часть околоплодника (косточка) становится очень твердой, а мучнистый наружный слой приобретает окраску от ярко-желтой (каркас голый) до почти черной (каркас южный). Костянки каркасов охотно поедаются птицами и ими же распространяются.Семена каркасов прорастают обычно весной следующего года, всходы, как и у ильмовых, развиваются по гетеробластному типу и в первый же год переходят к симподиальному ветвлению.Растут каркасы сравнительно медленно и долго живут (до 200, а по отдельным сведениям — до 600 лет). Однако в настоящее время старых крупных деревьев каркаса немного. Высокая ценность его древесины и произрастание главным, образом в малолесных районах влекут за собой регулярные рубки молодых деревьев. После рубок деревья сравнительно быстро восстанавливаются благодаря интенсивному порослевому (от пня) возобновлению.Каркас нередко называют также каменным деревом за его твердую, крепкую, тяжелую (плотность 0,78) древесину. Несмотря на ряд ценных свойств, она все же не имеет большого промышленного значения и идет в настоящее время в основном на мелкие поделки и декоративные изделия. В аридных странах каркасы издавна разводят, любят эти деревья и у нас в Средней Азии, на Кавказе и в Крыму, где часто используют в озеленении поселков и городов, а также в защитных насаждениях.Наиболее близок к каркасам род птероцельтис, распространенный в Центральном Китае. Единственный вид его был описан русским ботаником К. И. Максимовичем и назван в честь коллектора этого растения — птероцельтис Татаринова (Pleroceltis tatarinovii). Очень своеобразные крылатые плоды птероцельтиса нередко рассматривают как переходное звено между бескрылыми плодами каркасовых и крылатками ильмовых, что дает основание сближать и сами таксоны, однако сходство плодов только внешнее — это пример конвергентного развития. Плод птероцельтиса — настоящая шаровидная костянка с очень толстым, крепким эндокарпием, выросты которого образуют деревянистые крылья, утончающиеся к краю. Вверху крылья широко расставлены, между ними и изолированно от них, в отличие от ильмовых, расположен столбик с двумя рыльцами (рис. 132). Птероцельтис и по другим признакам типичный представитель подсемейства каркасовых: у него однополые цветки, костянки с 4-слойным околоплодником, однослойная семенная кожура, складчато-сложенный зародыш, 3 жилки в основании листа, основное хромосомное число х равно 20. Растет птероцельтис обычно вдоль рек и по скалистым местам: на сравнительно небольших высотах (до 1200 м над уровнем моря). Чаще всего это деревья высотой 12—17 м, с раскидистой кроной и короткими толстыми стволами диаметром до 1,5 м.Очень интересный и, по-видимому, монотипный род хетакма (Chaetacme) распространен на Мадагаскаре и в экваториальной и Южной Африке. Хетакма остистая развивается как небольшое деревце или кустарник высотой 3—7 м, она имеет блестящие кожистые листья, иногда оканчивающиеся топким заострением — колючкой, однополые цветки и мелкие плоды с твердой косточкой. Вид этот очень полиморфен и включает растения с однодомными и двудомными цветками, представлен сильно опушенными и голыми формами, колючими и без колючек, листья у него зубчатые, колючезубчатые или цельнокрайние, Хетакма остистая произрастает в зоне, переходной от леса к саванне, встречается в листопадных, полулистопадных лесных и кустарниковых формациях, в склерофильных галерейных лесах, где иногда вместе с другими растениями образует непроходимые колючие заросли, столь характерные для этих стран.В тропических, странах широко известен род трема, представленный преимущественно вечнозелеными деревцами, иногда кустарниковидными растениями, высотой 2—16 м, распространенными на всех континентах и многих островах от низменностей до 2500 м над уровнем моря. У них негустая распластанная крона, разветвленные пазушные соцветия, несущие многочисленные мелкие, обычно однополые цветки. Плоды — небольшие мясистые костянки, ярко-желто-оранжевые у тремы Ламарка и темные у тремы восточной и тремы мелкоцветковой.Виды рода трема трудно разграничиваются, и среди систематиков до сих пор имеются разногласия относительно их объема и числа. По-видимому, в роде не более 20 видов, все они близки и образуют полиплоидный ряд с числом хромосом в соматических клетках от 20 до 160.Тремы — быстро растущие неприхотливые растения, обитающие на опушках вечнозеленых и полулистопадных лесов равнин и горных склонов, обычны вдоль дорог и на вырубках. Виды тремы — обычные компоненты вторичных растительных формаций тропиков, в частности, характерные представители своеобразных вторичных формаций с древовидными папоротниками, развивающихся после пожаров и рубок на месте дождевых горных тропических лесов. На Кубе это сообщество тремы мелкоцветковой (Trema micrantha) и папоротника циатеи древовидной (Cyathea arborea), со сплошным покровом из папоротника орляка хвостатого (Pteridium caudatum). Аналогичные растительные формации характерны и для тропиков Старого Света. На Яве, например, имеются сообщества тремы восточной и циатеи грязноватой (Cyathea contaminans), в подлеске — посконник инулолистный (Eupatorium inulifolium).Трема пока единственный род в порядке крапивных, у которого обнаружен симбиоз с азотфиксирующими бактериями. На корнях тремы восточной недавно были найдены клубеньковые бактерии из группы Rhizobium, что дает основание относить ее к почвоулучшающим растениям. Возможно, отчасти благодаря этому свойству трему охотно используют на плантациях кофейного дерева и какао для создания негустого полога, под которым и высаживают эти культуры.Очень близок к роду трема и трудно отличим от него небольшой род параспония (Parasponia), распространенный на островах Океании. Представители обоих родов — типичные пионерные растения, отмечены как первые поселенцы лавовых потоков (остров Бали).Род афананта (Aphananthe) имеет разорванный ареал, охватывающий отдаленные друг от друга районы тропической и теплоумеренной Азии, включая Малезию, районы Соломоновых островов, Восточной Австралии, Мадагаскара и Мексики. Эти огромные дизъюнкции говорят о более широком распространении афананты в прошлом.В тропиках Южной Азии, Индонезии и Океании (в основном на тропических островах) распространен род жироньера (Gironniera), представленный огромными вечнозелеными деревьями дождевых тропических лесов (жироньера каркасолистная — G. celtidiiolia) или небольшими, высотой до 16 м, деревьями более сухих полулистопадных формаций тропиков (жироньера полуравная — G. subaequalis и др.).
Семена каркасовых обычно округлые, зародыш поперек согнут, поперек сложен или свернут в спираль; в зрелых семенах эндосперм сохраняется, он окружает зародыш и заполняет углубления его складок; семенная кожура всегда однослойная, околоплодник 3—4-слойный.Основное хромосомное число у каркасовых такое же, как у коноплевых (х —10). Характерна полиплоидия, причем отдельные виды достигают очень высокой степени плоидности (трема амбоинская — Trema amboinensis — 16-плоид!).Листья у большинства каркасовых имеют 3 четко выраженные базальные жилки, что приближает их жилкование к пальчатому типу. У большинства каркасов Южной Америки на нижней стороне листовой пластинки в результате разрастания оснований этих жилок (вероятно, под воздействием поселяющихся в них клещиков) развиваются специфические структуры, похожие на вздутые кармашки. Наконец, листьям каркасовых свойственны округлые цистолиты и многообразный характер опушения (у вест-индской тремы Ламарка на одном листе развивается до 4 типов волосков, включая и железистые).Основными родами подсемейства каркасовых являются 2 пантропических рода: каркас (Celtis, табл. 36) и трема (Trema), включающие более 85% всех видов. Каркас — не только самый крупный (более 50 видов), наиболее полиморфный, но и наиболее широко распространенный род. Его ареал опоясывает земной шар огромной полосой, северная граница которой колеблется около 40° северной широты, проходя по Японии, континентальной Азии, Кавказу, Южной Европе и Северной Америке; южная идет примерно по 35o южной широты через Новую Каледонию, Восточную Австралию, Капскую область Африки и Южную Аргентину. Несмотря на большую полиморфность, каркасы сохраняют в пределах всего ареала единый тип строения цветков, плодов и листьев. Виды каркаса или диплоидные (число хромосом в соматических клетках 2n = 20), или тетраплоидные (2 n = 40) растения. К последним относятся все виды СССР, распространенные также в Средиземноморье: каркас кавказский (С. caucasica), каркас голый (С. glabrata), каркас южный (С. australis), каркас Турнефора (С. tournefortii). Это листопадные, летнезеленые деревья с гладкой светло-серой корой и раскидистой кроной, достигающие в высоту иногда 30 м, со стволом диаметром до 3 м, или растущие небольшими деревцами и приобретающие кустарниковидную форму в неблагоприятных условиях среды (каркас голый и каркас Турнефора).Виды каркаса занимают различные экологические ниши и растут в разнообразных сообществах. Каркас кавказский и каркас Турнефора чаще распространены в сухих лесах и аридных редколесьях, на низменностях и в горах, нередко поднимаясь до высоты 2500—2800 м над уровнем моря, обычно в местах, удаленных от моря. Напротив, каркас южный и каркас голый растут преимущественно в приморских районах. Все эти виды обитают на открытых каменистых склонах и в ущельях, среди скал, на осыпях, по каменистым берегам небольших рек или по склонам к морю.Богато представлены и разнообразны каркасы тропических стран, где наряду с листопадными и полулистопадными обычны и вечнозеленые растения, причем у многих из них почечные чешуи не развиваются и зачатки формирующегося побега прикрыты лишь прилистниками кроющего листа.Очень своеобразны каркасы Австралии и особенно Новой Каледонии, отличающиеся цельно-крайними толстыми суккулентными листьями. У каркаса скученного (С. conferta), произрастающего в прибрежной зоне и нередкого среди мангровых зарослей, листья собраны на верхушке побега и иногда супротивны. В тропиках Азии, Африки и Южной Америки каркасы входят в сообщества вечнозеленых дождевых лесов низменностей (каркас Уайта — С. wightii, каркас Мильдбреда — С. mildbraedii), а также горных лесов (каркас Дюрана и др.). Растут тропические каркасы и в сухих вечнозеленых лесах, нередко образуя листопадные редколесья (каркас африканский — С. africana, каркас цельнолистный — С. integrifolia).В нарушенных рубкой лесах тропиков Нового Света интенсивно развиваются вечнозеленые лазящие лианы: каркас игуановый (С. iguanaea), каркас колючий (С. spinosa), каркас боливийский (С. boliviensis). В первые годы жизни эти каркасы растут как прямостоячие деревца или кустарники (высотой 1,5—5 м), позднее их верхние ветви интенсивно удлиняются и, цепляясь загнутыми колючками за близ стоящие деревья и кустарники, используют их как опору. Растение таким образом превращается в лазящую лиану и сохраняет этот облик до конца жизни.В теплоумеренном климате каркасы цветут весной, почти одновременно с распусканием листьев. Их мужские цветки раскрываются на несколько дней раньше обоеполых и женских. Цветки опыляются ветром, и, хотя их посещают насекомые, энтомофилия не имеет большого значения, так как пыльца лишена характерных признаков зерен энтомофильных растений и в момент выпрямления тычиночной нити мгновенно высыпается в окружающее пространство.Плоды созревают осенью, к этому времени внутренняя часть околоплодника (косточка) становится очень твердой, а мучнистый наружный слой приобретает окраску от ярко-желтой (каркас голый) до почти черной (каркас южный). Костянки каркасов охотно поедаются птицами и ими же распространяются.Семена каркасов прорастают обычно весной следующего года, всходы, как и у ильмовых, развиваются по гетеробластному типу и в первый же год переходят к симподиальному ветвлению.Растут каркасы сравнительно медленно и долго живут (до 200, а по отдельным сведениям — до 600 лет). Однако в настоящее время старых крупных деревьев каркаса немного. Высокая ценность его древесины и произрастание главным, образом в малолесных районах влекут за собой регулярные рубки молодых деревьев. После рубок деревья сравнительно быстро восстанавливаются благодаря интенсивному порослевому (от пня) возобновлению.Каркас нередко называют также каменным деревом за его твердую, крепкую, тяжелую (плотность 0,78) древесину. Несмотря на ряд ценных свойств, она все же не имеет большого промышленного значения и идет в настоящее время в основном на мелкие поделки и декоративные изделия. В аридных странах каркасы издавна разводят, любят эти деревья и у нас в Средней Азии, на Кавказе и в Крыму, где часто используют в озеленении поселков и городов, а также в защитных насаждениях.Наиболее близок к каркасам род птероцельтис, распространенный в Центральном Китае. Единственный вид его был описан русским ботаником К. И. Максимовичем и назван в честь коллектора этого растения — птероцельтис Татаринова (Pleroceltis tatarinovii). Очень своеобразные крылатые плоды птероцельтиса нередко рассматривают как переходное звено между бескрылыми плодами каркасовых и крылатками ильмовых, что дает основание сближать и сами таксоны, однако сходство плодов только внешнее — это пример конвергентного развития. Плод птероцельтиса — настоящая шаровидная костянка с очень толстым, крепким эндокарпием, выросты которого образуют деревянистые крылья, утончающиеся к краю. Вверху крылья широко расставлены, между ними и изолированно от них, в отличие от ильмовых, расположен столбик с двумя рыльцами (рис. 132). Птероцельтис и по другим признакам типичный представитель подсемейства каркасовых: у него однополые цветки, костянки с 4-слойным околоплодником, однослойная семенная кожура, складчато-сложенный зародыш, 3 жилки в основании листа, основное хромосомное число х равно 20. Растет птероцельтис обычно вдоль рек и по скалистым местам: на сравнительно небольших высотах (до 1200 м над уровнем моря). Чаще всего это деревья высотой 12—17 м, с раскидистой кроной и короткими толстыми стволами диаметром до 1,5 м.Очень интересный и, по-видимому, монотипный род хетакма (Chaetacme) распространен на Мадагаскаре и в экваториальной и Южной Африке. Хетакма остистая развивается как небольшое деревце или кустарник высотой 3—7 м, она имеет блестящие кожистые листья, иногда оканчивающиеся топким заострением — колючкой, однополые цветки и мелкие плоды с твердой косточкой. Вид этот очень полиморфен и включает растения с однодомными и двудомными цветками, представлен сильно опушенными и голыми формами, колючими и без колючек, листья у него зубчатые, колючезубчатые или цельнокрайние, Хетакма остистая произрастает в зоне, переходной от леса к саванне, встречается в листопадных, полулистопадных лесных и кустарниковых формациях, в склерофильных галерейных лесах, где иногда вместе с другими растениями образует непроходимые колючие заросли, столь характерные для этих стран.В тропических, странах широко известен род трема, представленный преимущественно вечнозелеными деревцами, иногда кустарниковидными растениями, высотой 2—16 м, распространенными на всех континентах и многих островах от низменностей до 2500 м над уровнем моря. У них негустая распластанная крона, разветвленные пазушные соцветия, несущие многочисленные мелкие, обычно однополые цветки. Плоды — небольшие мясистые костянки, ярко-желто-оранжевые у тремы Ламарка и темные у тремы восточной и тремы мелкоцветковой.Виды рода трема трудно разграничиваются, и среди систематиков до сих пор имеются разногласия относительно их объема и числа. По-видимому, в роде не более 20 видов, все они близки и образуют полиплоидный ряд с числом хромосом в соматических клетках от 20 до 160.Тремы — быстро растущие неприхотливые растения, обитающие на опушках вечнозеленых и полулистопадных лесов равнин и горных склонов, обычны вдоль дорог и на вырубках. Виды тремы — обычные компоненты вторичных растительных формаций тропиков, в частности, характерные представители своеобразных вторичных формаций с древовидными папоротниками, развивающихся после пожаров и рубок на месте дождевых горных тропических лесов. На Кубе это сообщество тремы мелкоцветковой (Trema micrantha) и папоротника циатеи древовидной (Cyathea arborea), со сплошным покровом из папоротника орляка хвостатого (Pteridium caudatum). Аналогичные растительные формации характерны и для тропиков Старого Света. На Яве, например, имеются сообщества тремы восточной и циатеи грязноватой (Cyathea contaminans), в подлеске — посконник инулолистный (Eupatorium inulifolium).Трема пока единственный род в порядке крапивных, у которого обнаружен симбиоз с азотфиксирующими бактериями. На корнях тремы восточной недавно были найдены клубеньковые бактерии из группы Rhizobium, что дает основание относить ее к почвоулучшающим растениям. Возможно, отчасти благодаря этому свойству трему охотно используют на плантациях кофейного дерева и какао для создания негустого полога, под которым и высаживают эти культуры.Очень близок к роду трема и трудно отличим от него небольшой род параспония (Parasponia), распространенный на островах Океании. Представители обоих родов — типичные пионерные растения, отмечены как первые поселенцы лавовых потоков (остров Бали).Род афананта (Aphananthe) имеет разорванный ареал, охватывающий отдаленные друг от друга районы тропической и теплоумеренной Азии, включая Малезию, районы Соломоновых островов, Восточной Австралии, Мадагаскара и Мексики. Эти огромные дизъюнкции говорят о более широком распространении афананты в прошлом.В тропиках Южной Азии, Индонезии и Океании (в основном на тропических островах) распространен род жироньера (Gironniera), представленный огромными вечнозелеными деревьями дождевых тропических лесов (жироньера каркасолистная — G. celtidiiolia) или небольшими, высотой до 16 м, деревьями более сухих полулистопадных формаций тропиков (жироньера полуравная — G. subaequalis и др.).
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.
.