- Гематоэнцефалический барьер
-
Гематоэнцефалический барьер
 Схема строения гематоэнцефалического барьера
Схема строения гематоэнцефалического барьера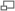
Гематоэнцефалический барьер (ГЭБ) (от др.-греч. αἷμα, род.п. αἷματο — «кровь» и др.-греч. εγκεφαλος — «головной мозг») — физиологический барьер между кровеносным руслом и центральной нервной системой. Имеется у всех позвоночных. Главной функцией гематоэнцефалического барьера является поддержание гомеостаза мозга.
Гематоэнцефалический барьер защищает мозг от циркулирующих в крови микроорганизмов, токсинов и различных других веществ. Он выполняет функцию высокоселективного фильтра, через который в мозг поступают питательные вещества, а из мозга выводятся продукты его жизнедеятельности.
С другой стороны наличие ГЭБ затрудняет лечение многих заболеваний, так как он не пропускает целый ряд лекарственных препаратов.
Первые исследования показавшие наличие барьера между кровью и мозгом были проведены Паулем Эрлихом в 1885 году. Окончательные доказательства существования ГЭБ были получены в 1967 году при электрономикроскопических исследованиях.
Содержание
Задачи гематоэнцефалического барьера
Масса головного мозга человека составляет приблизительно 2 % от массы его тела. При этом потребность в кислороде центральной нервной системы составляет 20 % от потребностей всего организма. Также в противоположность другим органам мозг обладает наименьшими запасами питательных веществ. Нервные клетки не могут обеспечить свои энергетические потребности анаэробно (путём одного лишь гликолиза). Прекращение поступления крови к мозгу в течение нескольких секунд приводит к потере сознания, а через 10 минут отмечается полная гибель нейронов [1]. Данная особенность головного мозга требует от ГЭБ активно транспортировать кислород и питательные вещества[2].
Нормальное функционирование мозга возможно также в условиях электролитного и биохимического гомеостаза. Колебания pH, концентрации калия крови и других показателей не должны отражаться на ткани головного мозга. Попадение циркулирующих в кровеносном русле нейромедиаторов в нервную ткань может разбалансировать её работу [1]. Также мозг должен быть защищён от попадания в него чужеродных веществ, таких как ксенобиотики и патогенные микроорганизмы. ГЭБ представляет собой в том числе и иммунологический барьер, так как является непроницаемым для различных микроорганизмов, антител и лейкоцитов[3] [4].
Чтобы обеспечить задачи обеспечения, выведения продуктов жизнедеятельности и поддержания гомеостаза вещества мозга, система сосудов центральной нервной системы имеет целый ряд структурно-функциональных отличий от сосудов других органов и тканей[1].
Изменения в функционировании ГЭБ могут вызывать нарушения функционирования центральной нервной системы. Целый ряд неврологических заболеваний напрямую или косвенно связан с его повреждением[2].
Строение
 Сравнительная схема строения периферического и церебрального капилляров
Сравнительная схема строения периферического и церебрального капилляров
нем. Periphere Kapillare — периферический капилляр
нем. Zerebrale Kapillare — церебральный капилляр
нем. Zellkern — клеточное ядро
нем. Lumen des Kapillargefäßes — просвет капиллярного сосуда
англ. Tight Junction — плотный контакт
нем. Intrazellularspalt — межклеточная щель
нем. Endothelzelle — эндотелиальная клетка
нем. Fenestrierung — фенестрация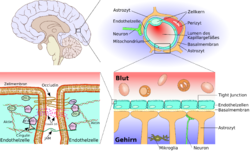 Строение ГЭБ — от ткани мозга к плотному контакту
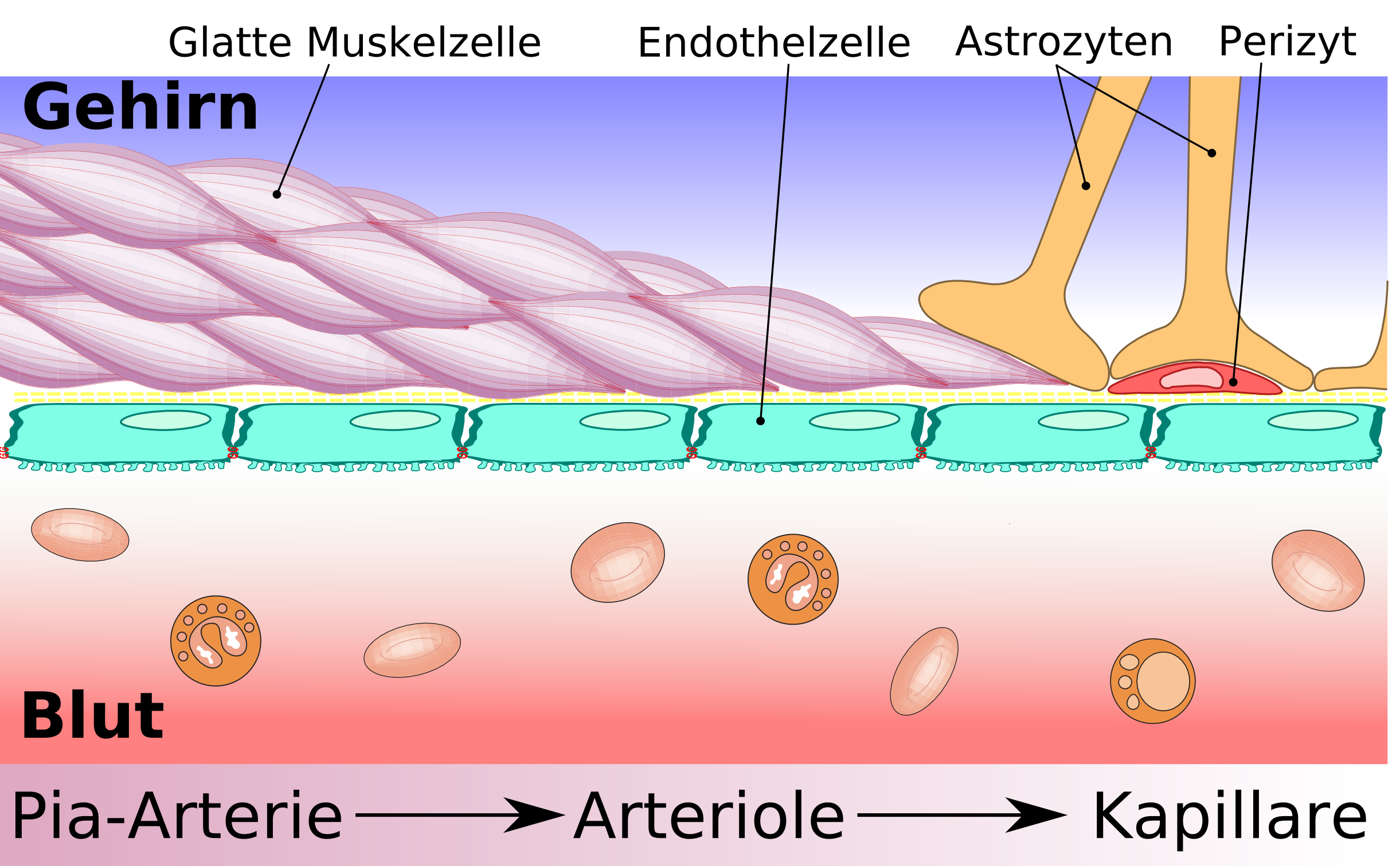
Строение ГЭБ — от ткани мозга к плотному контакту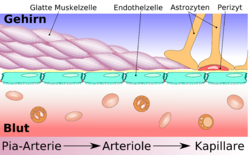 Схематическое строение сосудистой стенки артерии, артериолы и капилляра мозга
Схематическое строение сосудистой стенки артерии, артериолы и капилляра мозгаСущественным элементом структуры ГЭБ являются эндотелиальные клетки. Особенностью эндотелия сосудистой стенки церебральных сосудов является наличие между ними плотных межклеточных контактов. В структуре ГЭБ также большое значение имеют перициты и астроциты[1]. Межклеточные промежутки между эндотелиальными клетками, перицитами и астроцитами нейроглии ГЭБ являются наиболее узкими в сравнении с другими клетками организма. Эти три вида клеток являются структурной основой ГЭБ не только у человека, но и у большинства позвоночных[5] [6].
Эндотелий
Капиллярные сосуды выстланы эндотелиальными клетками. Эндотелий периферических сосудов содержит открытые промежутки (фенестрации) диаметром около 50 нм. и межклеточные щели от 0,1 до 1 мкм. Через эти пространства происходит свободная циркуляция воды и растворённых в ней веществ между кровью и межклеточным пространством. В церебральных сосудах между эндотелиальными клетками отсутствуют как фенестрации, так и межклеточные щели[7]. Таким образом можно говорить о сплошной эндотелиальной выстилке просвета капилляров мозга[8].
Другим отличием эндотелия церебральных капилляров от периферических является низкое количество в них пиноцитозных пузырьков (везикул)[9] [10].
В то же время количество митохондрий в эндотелиальных клетках сосудов мозга в 5-10 раз выше, чем в эндотелии периферических сосудов. Митохондрии являются органеллами синтезирующими молекулы АТФ, являющихся основным источником энергии для клетки. Большое количество митохондрий соответственно является показателем значительных энергетических потребностей эндотелиальных клеток ГЭБ, что связано с процессами активного транспорта и обмена веществ[4].
ГЭБ является также метаболическим или ферментативным (энзиматическим) барьером [11] [12] [13] [14] [15]. На поверхности клеточных мембран эндотелиальных клеток ГЭБ находится целый ряд ферментов в значительно большем количестве чем на других клетках паренхимы. Среди них стоит отметить гамма-глутамилтрансферазы и фосфатазы (в частности глюкоза-6-фосфатазу), катехол-О-метилтрансферазу, моноаминоксидазу и цитохром Р450[16] [17] [18]. В связи с большой концентрацией различных ферментов в эндотелиальных клетках ГЭБ многие вещества при транспортировании через цитоплазму эндотелия метаболизируются[10]. При этом по высоте эндотелиальная клетка ГЭБ составляет от 0,3 до 0,5 мкм. Энтероциты, эпителиальные клетки кишечника, к примеру имеют в высоту 17-30 мкм[19].
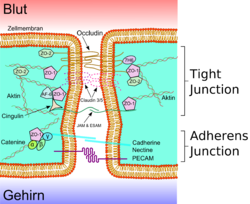 Схематическое изображение плотного контакта
Схематическое изображение плотного контакта Схематическое изображение плотного контакта
Схематическое изображение плотного контактаСоотношение холестерина к фосфолипидам в эндотелиальных клетках ГЭБ такое же, как и в эндотелиальных клетках периферических сосудов и составляет ≈ 0,7[20]. Пассивный транспорт через клеточные мембраны ГЭБ мало чем отличается от пассивной диффузии в других эндотелиальных клетках[21]. В мембранах эндотелиальных клеток содержится большое количество каналов, которые свободно пропускают моллекулы воды. Они делают возможным свободную диффузию моллекул воды как в направлении мозга, так и кровеносной системы[22].
Отсутствие фенестраций и небольшое число пиноцитарных везикул делают эндотелиальную выстилку капилляров мозга механическим барьером для крупных молекул и инородных веществ. Кроме этого ГЭБ обладает значительным электрическим сопротивлением — около 1500—2000 Ом. К примеру электрическое сопротивление для стенок капилляров мышечной ткани составляет 30 Ом[23].
Плотные контакты
Эндотелиальные клетки плотно прилежат к другу. Между соседними клетками образуются так называемые плотные контакты. Они вносят значительный вклад в обеспечении главной функции ГЭБ — предотвращении проникновения в ткань мозга различных нежелательных веществ из кровеносного русла[24] [25]. Плотные контакты между эндотелиальными клетками блокируют межклеточный (парацеллюлярный) пассивный транспорт[26] [27] [28]. При этом блокируется парацеллюлярный транспорт веществ как из кровеносного русла в ткань мозга, так и в обратном направлении — из мозга в кровь[6].
Большое количество трансмембранных белков, таких как окклюдин, разнообразные клаудины и замыкательные адгезионные молекулы связывают латеральные отделы клеточных стенок между собой, участвуют в формировании плотных контактов и делают возможным межклеточный транспорт и обмен веществ[29]. Основными белками обеспечивающими адгезию эндотелиальных клеток и формирование плотных контактов явдяются клаудин-5 и клаудин-24 [30]. Выключение (блокирование) CLDN5—гена, ответственного за синтез белка клаудина-5, приводил у подопытных мышей к тому, что их ГЭБ становился проницаемым для молекул с молярной массой до 800 г/моль. Такие подопытные генетически изменённые животные умирали через несколько часов после рождения[31].
Базальная мембрана
 Базальная мембрана эпителиальной клетки
Базальная мембрана эпителиальной клеткиЭндотелиальные клетки полностью покрывают подлежащий белковый слой, называемый базальной мембраной[8]. Вертикальный размер базальной мембраны колеблется от 40 до 50 нм. Она различима только под электронным микроскопом. Состоит в основном из коллагена IV типа, гепаринсульфат-протеогликанов, ламининов, фибронектина и других белков внеклеточного матрикса. Со стороны мозга базальная мембрана ограничена плазматической мембраной пластинчатых окончаний отростков астроцитов[10] [26].
Перициты
 Электронномикроскопическое изображение перицита (справа) и просвета сосуда с тремя эритроцитами (слева)
Электронномикроскопическое изображение перицита (справа) и просвета сосуда с тремя эритроцитами (слева)Перициты, ранее называвшиеся по имени первооткрывателя Шарля Мари Бенджамина Руже (1824—1904) клетками Руже[32], являются важной составной частью ГЭБ[33]. Они обладают несколькими важными для его функционирования свойствами: способностью к сокращению, регулированию функций эндотелия и макрофагальной активностью[34].
 Щелевые клеточные соединения (схема)
Щелевые клеточные соединения (схема)Около 20 % поверхности эндотелиальных клеток церебральных капилляров покрыты относительно маленькими, овальными перицитами. Каждая 2—4-я эндотелиальная клетка имеет контакт с клеткой-перицитом[6]. В основном перициты располагаются в местах контакта эндотелиальных клеток[35] [36]. Перициты имеются практически во всех артериолах, венулах и капиллярах организма. Уровень покрытия ими эндотелиального слоя капилляра коррелирует с проницаемостью сосудистой стенки. В органах и тканях с проницаемой сосудистой стенкой они могут мигрировать из кровеносного русла в межклеточное пространство. Так например в капиллярах скелетной мускулатуры соотношение перициты:эндотелиоциты составляет 1:100[37] [38].
Перициты, как и эндотелиоциты располагаются на базальной мембране[8].
Также перициты синтезируют целый ряд вазоактивных веществ[38] и играют важную роль в ангиогенезе[39] [40].
Клеточные контакты перицит — эндотелиоцит
 Фокальные клеточные адгезии
Фокальные клеточные адгезииПерициты крепко связаны с эндотелиоцитами. Эта связь осуществляется благодаря трём типам контактов: щелевым соединениям, фокальным адгезиям и инвагинациям мембраны одной клетки в полость другой[34]. Щелевые соединения непосредственно связывают цитоплазму двух клеток, являясь проницаемыми для ионов и небольших молекул[41]. С помощью фокальных адгезий осуществляется прочная механическая связь двух типов клеток[42]. нвагинации участков цитоплазмы одной клетки в другую обеспечивают как механическое связывание так и межклеточный обмен веществ[34] [43].
Благодаря тесным контактам клетки опосредованно влияют на митотическую активность, экспрессию генов и соответственно фенотип друг друга[39].
Сократительная функция
Перициты содержат большое количество способного к сокращению белка актина. Благодаря этой своей структурной особенности они в состоянии изменять просвет капилляров и таким образом регулировать местное кровяное давление[44] [45].
Макрофагальная активность
Данное свойство характерно только для церебральных перицитов. В капиллярной сети мозга они выполняют функцию макрофагов. Соответственно в цитоплазме церебральных перицитов располагается большое количество лизосом. В культуре тканей доказана способность перицитов к фагоцитозу[34] [46] [47] и презентации антигенов [48] [49].
 Астроцит (окрашен зелёным) в клеточной культуре
Астроцит (окрашен зелёным) в клеточной культуреМакрофагальные свойства перицитов образуют «вторую линию защиты мозга» от нейротоксических молекул, которые преодолели барьер эндотелиальных клеток[50]. Таким образом они являются важной составной частью иммуной системы мозга. Сбой макрофагальной активности перицитов может стать одним из факторов развития целого ряда аутоиммунных заболеваний. Имеются данные об опосредованной роли перицитов в развитии болезни Альцгеймера[51] [52].
Астроциты
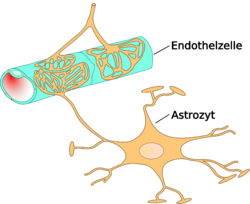 Взаимоотношение астроцитов и эндотелиоцитов
Взаимоотношение астроцитов и эндотелиоцитовАстроциты — большие нейроглиальные клетки звёздчатой формы. Своими отростками лни выстилают стенки мозговых капилляров со стороны мозговой ткани. В то же время, несмотря на то, что пластинчатыми окончаниями их клеточных отростков выстлано около 99% капиллярных сосудов, астроциты не выполняют прямой барьерной функции[6] [53]. Астроциты тесно взаимодействуют с эндотелиальными клетками. Между ними осуществляется постоянный обмен веществ[54]. Астроглиальные клетки индуцируют возникновение и формирование ГЭБ. При проведении экспериментов по пересадке сосудов мозга в периферические органы и наоборот — периферических сосудов в ткань головного мозга, отмечено формирование ГЭБ в периферических сосудах пересаженных в мозг (образование плотных контактов, перестройка эндотелиальных клеток) и разобщение эндотелиальных клеток и появление фенестраций между ними при пересадке мозговые сосуды[1] [55]. Также in vitro показано влияние астроцитов на фенотип эндотелия. В клеточной культуре содержащей астроциты и эндотелиоциты отмечено более плотное расположение эндотелия по сравнению с его чистой клеточной культурой[56].
Астроциты выделяют целый ряд веществ, которые влияют на проницаемость эндотелия[57]. Эндотелиоциты в свою очередь выделяют ингибирующий лейкемию фактор (LIF), цитокин интерлейкин-6, которые воздействуют на процесс дифференциации астроцитов[57]. Расстояние от пластинчатых окончаний отростков астроцитов до клеток эндотелия и перицитов составляет всего лишь 20 нм[8] [58].
Главными задачами астроглиальных клеток является обеспечение нейронов питательными веществами и поддержание необходимой концентрации электролитов внеклеточного пространства[57] [59]. Астроциты синтезируют большую часть необходимого клеткам мозга холестерина. Холестерин не проникает через ГЭБ. В то же время в ткани мозга находится 25% от общего холестерина организма. Большая его часть входит в состав миелина, который окутывает отростки нейронов аксоны. Нарушения процессов миелинизации нервных волокон вызывают развитие демиелинизирующих заболеваний, в частности рассеянный склероз[60].
Пластинчатые окончания отростков астроцитов неплотно покрывают со стороны мозга базальную мембрану сосудистой стенки с расположенными на ней эндотелиоцитами и перицитами. За счёт этого между эндотелиоцитами и тканью мозга возможна прямая диффузия различных веществ[57].
Заболевания, при которых происходит прямое или опосредованное поражение астроцитов (например, болезнь Альцгеймера, астроцитомы), сопровождаются нарушением функционирования ГЭБ.
Области мозга без ГЭБ
ГЭБ имеется в капиллярах большинства, но не всех областей мозга. В 6 анатомических образованиях мозга ГЭБ отсутствует:
- Самое заднее поле (лат. area postrema) ромбовидной ямки (дна IV желудочка) — располагается между треугольником блуждающего нерва (лат. trigonum nervi vagi) с окрамляющим его самостоятельным канатиком (лат. funiculus separans) и бугорком тонкого ядра[61]
- шишковидное тело (лат. corpus pineale) (синоним — эпифиз)
- Нейрогипофиз
- Прикреплённая пластинка (лат. lamina affixa) — эмбриональный остаток стенки конечного мозга, покрывающий верхнюю поверхность таламуса. Медиально она истончается, образует извитую пластинку — сосудистую ленту (лат. tenia choroidea)[62]
- Субфорникальный орган
- Субкомиссуральный орган
Данная гистологическая особенность имеет своё обоснование. Так например, нейрогипофиз выделяет в кровь гормоны, которые не могут пройти через ГЭБ, а нейроны лат. area postrema улавливают в крови наличие токсических веществ и стимулируют рвотный центр[63]. Защитным барьером соседней с данными образованиями мозговой ткани является скопление таницитов. Они представляют собой клетки эпендимы с плотными контактами[64].
Характеристика ГЭБ
Сеть из более чем 100 миллиардов капилляров суммарной протяжённостью около 600 км пронизывает мозг взрослого человека[21]. В среднем просвет капилляра составляет около 40 нм[65]. Мозг кровоснабжается значительно сильнее многих других тканей и органов организма. Наибольшая плотность сосудов характерна для коры головного мозга — от 300 до 800 капилляров на 1 мм2 ткани[1].
Суммарная поверхность стенок сосудов мозга составляет 12[66] — 20[67] м2. Ежеминутно через сосудистую сеть мозга протекает около 610 мл крови со средней скоростью 1 мм/с. Давление в ней колеблется в пределах 15-35 мм рт. ст.[4] Среднее время прохождения крови через капиллярное русло (англ. mean transit time (MTT)) составляет всего лишь 5 секунд. Для сравнения, в кишечнике, площадь сосудов которого достигает 180 м2 оно равно 40 часам[68] [69], а в печени с 70 м2 — 30 секунд[70] [71] [72].
Развитие
До конца 20-го столетия считалось, что у эмбриона и новорожденных ГЭБ не сформирован в полной степени и соответственно не выполняет своей функции. Причиной этого, до сих пор широко распространённого мнения, являются недостатки ранее проводившихся физиологических опытов. Эксперименты заключались в введении либо связанных с белками красителей либо других маркеров взрослым животным и эмбрионам. Первые опыты такого плана проводились в 1920 году [73]. Маркеры вводимые эмбрионам проникали в ткань мозга и спинномозговую жидкость в то время как у взрослых животных — нет. В ходе данных экспериментов был допущен ряд методических ошибок (использование чрезмерного объёма вводимого вещества, повышение осмотического давления) из-за которых происходило частичное повреждение сосудистой стенки и соответственно маркер попадал в ткань мозга[74] [75] [76]. При правильной постановке экспериментов пассажа маркера через сосудистую сеть отмечено не было[77] [78] [79].
В крови плода в большом количестве содержатся молекулы таких веществ как альбумин, α1-фетопротеин и трансферрин, отсутствуя при этом в межклеточном пространстве ткани мозга[80]. В эмбриональном эндотелии обнаружен транспортёр Р-гликопротеин[81]. Это свидетельствует о наличии ГЭБ в пренатальном периоде. В ходе развития организма происходит дальнейшее совершенствование ГЭБ[80].
Для небольших поляризированных молекул, например инулина и сахарозы, проницаемость ГЭБ эмбриона и новорожденного значительно выше чем у взрослых[82] [83] [84]. Схожий эффект отмечен и для ионов[85]. Транспорт аминокислот и инсулина через ГЭБ, по всей видимости в связи с большой потребностью в них растущего мозга, значительно ускорен[86] [87] [88] [89].
С другой стороны в мозге эмбриона имеется дополнительный, отсутствующий у взрослых, барьер на границе между ликвором и тканью мозга — так называемые ремневые контакты (англ. Strap Junctions) между клетками эпендимы[90].
Эволюция
В ходе эволюции нервной ткани от беспозвоночных к позвоночным с одной стороны отмечено её увеличение и приобретение центральных функций в жизнедеятельности организмов. С другой стороны большая масса мозга требует лучшего обеспечения питательными веществами и выведения ненужных и отработанных продуктов распада. Это привело к развитию густой капиллярной сети в ткани мозга. Следующим этапом эволюции стало появление защитного барьера от циркулирующих в крови токсичных для нейронов веществ — ксенобиотиков и токсинов[5] [91].
У многих беспозвоночных ГЭБ отсутствует. У них эндотелий капилляров нервной ткани не образует сплошной выстилки сосудистой стенки. У высших беспозвоночных — насекомых, ракообразных и головоногих[92] — защитный барьер между нейронами и кровью представлен исключительно глиальной тканью[93]. В этом случае речь идёт о глиальном гематоэнцефалическом барьере[94].
У всех видов позвоночных имеется ГЭБ, и у большинства из них он образован преимущественно клетками эндотелия сосудистой стенки скреплёнными между собой плотными контактами. Только у пластиножаберных (среди них акул и скатов), а также семейства осетровых рыб ГЭБ формируется периваскулярными астроцитами. Из этого следует, что в процессе эволюции происходит увеличение количества эндотелиальных клеток сосудов головного мозга, которые перенимают на себя барьерные функции.
Структурные различия глиального и эндотелиального гематоэнцефалических барьеров достаточно велики. Эндотелиальный барьер имеет целый ряд преимуществ. Одним из них является строгое разграничение функций эндотелиальных клеток и клеток астроглии, которые обеспечивают гомеостаз внеклеточной среды вещества мозга[93].
Гематоликворный барьер
Кроме гематоэнцефалического барьера существует также гематоликворный, который ограничивает центральную нервную систему от кровеносного русла. Он образован эпителиальными клетками с плотными контактами выстилающими сосудистое сплетение желудочков мозга[95] [96]. Гематоликворный барьер также имеет свою роль в поддержании гомеостаза мозга. Через него из крови в омывающую мозг спинномозговую жидкость поступают витамины, нуклеотиды и глюкоза. Общий вклад гематоликворного барьера в процессы обмена между мозгом и кровью невелик. Суммарная площадь гематоликворного барьера сосудистых сплетений желудочков мозга приблизительно в 5000 раз меньше в сравнении с площадью гематоэнцефалического.
Кроме гематоэнцефалического и гематоликворного барьеров в организме человека существуют гематоплацентарный, гематотестикулярный, гематоклубочковый, гематоретинальный, гематотимальный и гематолёгочный барьеры.
Транспорт веществ через ГЭБ
 Схема транспорта различных веществ черех гематоэнцефалический барьер
Схема транспорта различных веществ черех гематоэнцефалический барьер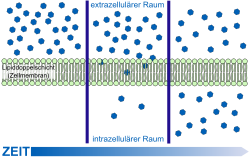 Простая диффузия через клеточную мембрану
Простая диффузия через клеточную мембрануГематоэнцефалический барьер не только задерживает и не пропускает целый ряд веществ из крови в вещество мозга, но и выполняет противоположную функцию — транспортируют необходимые для метаболизма ткани мозга вещества. Гидрофобные вещества и пептиды проникают в мозг либо с помощью специальных транспортных систем, либо каналы клеточной мембраны. Для большинства других веществ возможна пассивная диффузия[11] [15].
Межклеточный транспорт
В капиллярах периферических органов и тканей, транспорт веществ осуществляется в основном через фенестрации сосудистой стенки и межклеточные промежутки. В норме между клетками эндотелия сосудов мозга такие промежутки отсутствуют. В связи с этим питательные вещества проникает в мозг лишь через клеточную стенку[97]. Вода, глицерин и мочевина являются примерами тех небольших поляризированных молекул, которые могут свободно диффундировать через плотные контакты между эндотелиальными клетками ГЭБ[98].
Свободная диффузия
 Модель пассивной диффузии через клеточную мембрану
Модель пассивной диффузии через клеточную мембрануСамой простой формой транспорта через ГЭБ является свободная (или пассивная) диффузия. Она может осуществляться как через клеточные мембраны эндотелиоцитов так и через плотные межклеточные контакты. Для диффузии веществ движущей силой является разница концентраций. Диффузия веществ пропорциональна разнице концентраций в кровяном русле и ткани мозга. Для неё не требуется затрат клеточной энергии[99].
Липофильные структурные элементы клеточной мембраны, а также плотные межклеточные контакты снижают количество веществ, которые могут свободно диффундировать через ГЭБ. Проницаемость ГЭБ напрямую зависит от липофильности каждого конкретного вещества[100].
Проницаемость ГЭБ также зависит от молярной массы вещества. Молекулы с массой более 500 г/моль не могут диффундировать через ГЭБ. В то же время ГЭБ не является механическим барьером, который свободно пропускает молекулы меньшего размера и не пропускает большего. Процесс клеточной диффузии является динамическим, при этом он легче для веществ с молярной массой 200 г/моль, чем для веществ с 450 г/моль[20] [101].
Чем липофильнее и меньше вещество, тем легче оно диффундирует через клеточную мембрану[11].
Немецким биофизиком Германном Тройбле в 1971 году была высказана гипотеза о транспорте небольших молекул через клеточную мембрану. Согласно ней они проникают в клетку через небольшие промежутки между цепями жирных кислот двойного слоя мембраны. Эти промежутки изменчивы, их образование не требует клеточной энергии[102] [103] [104] [105]. Теория Тройбле была спектроскопически доказана в 1974 году[106] [107].
Прогноз и исследования относительно проницаемости ГЭБ для того или иного вещества возможно проводить как in vitro [15] [108] [109] [101] [110] так и in silico [111].
Липофильность и небольшая молекулярная масса не являются гарантией проницаемости ГЭБ для данного вещества. Высокомолекулярные соединения (например, моноклональные антитела, рекомбинантные белки и другие) удерживаются ГЭБ[112].
Функции
- поддержание гомеостаза
- транспортная
- защитная
Механизм действия
Гематоэнцефалический барьер непроницаем для множества соединений, как чужеродных, так и вырабатываемых самим организмом.
Гематоэнцефалический барьер препятствует проникновению в центральную нервную систему переносимых кровью токсических веществ, нейромедиаторов, гормонов, антибиотиков (что затрудняет лечение инфекционных поражений мозга и его оболочек), поддерживает электролитный баланс мозга, обеспечивает избирательный транспорт ряда веществ (глюкозы, аминокислот) из крови в мозг.
Для преодоления гематоэнцефалического барьера молекулы должны быть либо малы (как молекулы кислорода), либо обладать способностью растворяться в липидных компонентах мембран глиальных клеток (как этанол). Кроме того, некоторые вещества могут переноситься через гематоэнцефалический барьер путём активного транспорта.
Примечания
- ↑ 1 2 3 4 5 6 S. Wolf, B. Seehaus, Minol K. und andere Die Blut-Hirn-Schranke: Eine Besonderheit des cerebralen Mikrozirkulationssystems // Naturwissenschaften. — 1996. — № 83. — С. 302-311.
- ↑ 1 2 S. Ohtsuki New Aspects of the Blood–Brain Barrier Transporters; Its Physiological Roles in the Central Nervous System // Biological & Pharmaceutical Bulletin. — 2004. — № 27 (10). — С. 1489–1496.
- ↑ W. Risau, B. Engelhardt, H. Wekerle Immune function of the blood-brain barrier: incomplete presentation of protein (auto-) antigens by rat brain microvascular endothelium in vitro // The Journal of Cell Biology. — 1990. — № 110. — С. 1757–1766.
- ↑ 1 2 3 B. Bauer In vitro Zellkulturmodelle der Blut-Hirn-Schranke zur Untersuchung der Permeation und P-Glykoprotein-Interaktion von Arzneistoffen // Диссертация. Гейдельбергский университет им. Рупрехта-Карла. — 2002.
- ↑ 1 2 M. Bundgaard, N. J. Abbott All vertebrates started out with a glial blood-brain barrier 4-500 million years ago // Glia. — 2008. — № 56. — С. 699–708.
- ↑ 1 2 3 4 W. M. Pardridge Molecular biology of the blood–brain barrier // Mol Biotechnol. — 2005. — № 30 (1). — С. 57–70.
- ↑ J. C. Lee Evolution in the concept of the blood-brain barrier phenomen // Progress in neuropathology. — Verlag Grune und Stratton, 1971. — Т. 1. — С. 84–145. — ISBN 0-88167-188-6
- ↑ 1 2 3 4 M. Pavelka, J. Roth Funktionelle Ultrastruktur. — Verlag Springer. — С. 234–235. — ISBN 3-211-83563-6.
- ↑ 1 2 3 B. T. Hawkins, T. P. Davis The blood-brain barrier/neurovascular unit in health and disease // Pharmacol Rev. — 2005. — № 57. — С. 173–185.
- ↑ 1 2 3 S. Nobmann Isolierte Gehirn-Kapillaren als in vitro-Modell der Blut-Hirn Schranke // Диссертация. Гейдельбергский университет им. Рупрехта-Карла. — 2001.
- ↑ R. S. el-Bacha, A. Minn Drug metabolizing enzymes in cerebrovascular endothelial cells afford a metabolic protection to the brain // Cell Mol Biol. — 1999. — № 45. — С. 15–23.
- ↑ Chat M, Bayol-Denizot C, Suleman G, Roux F, Minn A. Drug metabolizing enzyme activities and superoxide formation in primary and immortalized rat brain endothelial cells // Life Sci. — 1998. — № 62. — С. 151–163.
- ↑ Minn A, Ghersi-Egea JF, Perrin R, Leininger B, Siest G. Drug metabolizing enzymes in the brain and cerebral microvessels // Life Sci. — 1991. — № 116. — С. 65–82.
- ↑ 1 2 3 Takakura Y, Audus KL, Borchardt RT. Blood-brain barrier: transport studies in isolated brain capillaries and in cultured brain endothelial cells // Adv Pharmacol. — 1991. — № 22. — С. 137–165.
- ↑ Méresse S, Dehouck MP, Delorme P, Bensaïd M, Tauber JP, Delbart C, Fruchart JC, Cecchelli R. Bovine brain endothelial cells express tight junctions and monoamine oxidase activity in long-term culture // J Neurochem. — 1989. — № 53. — С. 1363–1371.
- ↑ Perrin R, Minn A, Ghersi-Egea JF, Grassiot MC, Siest G. Distribution of cytochrome P450 activities towards alkoxyresorufin derivatives in rat brain regions, subcellular fractions and isolated cerebral microvessels // Biochem Pharmacol. — 1990. — № 40. — С. 2145–2151.
- ↑ Bendayan R, Lee G, Bendayan M. Functional expression and localization of P-glycoprotein at the blood brain barrier // Res Tech. — 2002. — № 57. — С. 365–380.
- ↑ Su Y, Sinko PJ. Drug delivery across the blood-brain barrier: why is it difficult? how to measure and improve it? // Expert Opin Drug Deliv. — 2006. — № 3. — С. 419–435.
- ↑ 1 2 Fischer H, Gottschlich R, Seelig A. Blood-brain barrier permeation: molecular parameters governing passive diffusion // J Membr Biol. — 1998. — № 165. — С. 201–211.
- ↑ 1 2 U. Fagerholm The highly permeable blood-brain barrier: an evaluation of current opinions about brain uptake capacity // J Membr Biol. — 2007. — № 12. — С. 1076–1082.
- ↑ Nico B, Frigeri A, Nicchia GP, Quondamatteo F, Herken R, Errede M, Ribatti D, Svelto M, Roncali L. Role of aquaporin-4 water channel in the development and integrity of the blood-brain barrier // J Cell Sci. — 2001. — № 114. — С. 1297–1307.
- ↑ Butt AM, Jones HC, Abbott NJ. Electrical resistance across the blood-brain barrier in anaesthetized rats: a developmental study // J Physiol. — 1990. — № 429. — С. 47-62.
- ↑ P. Claude, D. A. Goodenough Fracture faces of zonulae occludentes from "tight" and "leaky" epithelia // J Cell Biol. — 1973. — № 58. — С. 390-400.
- ↑ Wolburg H, Neuhaus J, Kniesel U, Krauss B, Schmid EM, Ocalan M, Farrell C, Risau W. Modulation of tight junction structure in blood-brain barrier endothelial cells. Effects of tissue culture, second messengers and cocultured astrocytes // J Cell Sci. — 1994. — № 107. — С. 1347–1357.
- ↑ 1 2 H. B. Newton Advances in strategies to improve drug delivery to brain tumors // Expert Rev Neurother. — 2006. — № 6. — С. 1495–1509.
- ↑ J. L. Madara Tight junction dynamics: is paracellular transport regulated? // Cell. — 1988. — № 53. — С. 497–498.
- ↑ H. C. Bauer et al. Proteins of the tight junctions in the blood-brain barrier // Blood-spinal Cord and Brain Barriers in Health and Disease. — Verlag Elsevier, 2004. — С. 1–10.
- ↑ Cecchelli R, Berezowski V, Lundquist S, Culot M, Renftel M, Dehouck MP, Fenart L. Modelling of the blood-brain barrier in drug discovery and development // Nat Rev Drug Discov. — 2007. — № 6. — С. 650–661.
- ↑ Matter K, Balda MS. Holey barrier: claudins and the regulation of brain endothelial permeability // J Cell Biol.. — 2003. — № 161. — С. 459–460.
- ↑ Nitta T, Hata M, Gotoh S, Seo Y, Sasaki H, Hashimoto N, Furuse M, Tsukita S. Size-selective loosening of the blood-brain barrier in claudin-5-deficient mice // J Cell Biol. — 2003. — № 161. — С. 653–660.
- ↑ P. Dore-Duffy Pericytes: pluripotent cells of the blood brain barrier // Curr Pharm Des. — 2008. — № 14. — С. 1581-1593.
- ↑ Balabanov R, Dore-Duffy P. Role of the CNS microvascular pericyte in the blood-brain barrier // J Neurosci Res.. — 1998. — № 53. — С. 637-644.
- ↑ 1 2 3 4 Rucker HK, Wynder HJ, Thomas WE. Cellular mechanisms of CNS pericytes // Brain Res Bull. — 2000. — № 51. — С. 363-369.
- ↑ P. A. D'Amore Culture and Study of Pericytes // Cell Culture Techniques in Heart and Vessel Research. — Verlag Springer, 1990. — С. 299. — ISBN 3-540-51934-3.
- ↑ N. J. Abbott Neurobiology. Glia and the blood-brain barrier // Nature. — 1987. — № 325. — С. 195.
- ↑ Lai CH, Kuo KH. The critical component to establish in vitro BBB model: Pericyte // Brain Res Brain Res Rev. — 2005. — № 50. — С. 258-265.
- ↑ 1 2 Shepro D, Morel NM. Pericyte physiology // FASEB. — 1993. — № 7. — С. 1031–1038.
- ↑ 1 2 Sims DE. Diversity within pericytes // Clin Exp Pharmacol Physiol. — 2000. — № 27. — С. 842–846.
- ↑ Engelhardt B. Development of the blood-brain barrier // Cell Tissue Res. — 2003. — № 314. — С. 119–129.
- ↑ Fujimoto K. Pericyte-endothelial gap junctions in developing rat cerebral capillaries: a fine structural study // Anat Rec. — 1995. — № 242. — С. 562-565.
- ↑ Díaz-Flores L, Gutiérrez R, Varela H, Rancel N, Valladares F. Microvascular pericytes: A review of their morphological and functional characteristics // Histol Histopath. — 1991. — № 6. — С. 269–286.
- ↑ D. E. Sims Recent advances in pericyte biology--implications for health and disease // Can J Cardiol. — 1991. — № 7. — С. 431–443.
- ↑ Herman IM, D'Amore PA. Microvascular pericytes contain muscle and nonmuscle actins // J Cell Biol. — 1985. — № 101. — С. 43–52.
- ↑ Hirschi KK, D'Amore PA. Pericytes in the microvasculature // Cardiovasc Res. — 1996. — № 32. — С. 687-698.
- ↑ Mato M, Ookawara S, Sugamata M, Aikawa E. Evidence for the possible function of the fluorescent granular perithelial cells in brain as scavengers of high-molecular-weight waste products // Experientia. — 1984. — № 40. — С. 399-402.
- ↑ Balabanov R, Washington R, Wagnerova J, Dore-Duffy P. CNS microvascular pericytes express macrophage-like function, cell surface integrin alphaM, and macrophage marker ED-2 // Microvasc Res. — 1996. — № 52. — С. 127-142.
- ↑ Hickey WF, Kimura H. Perivascular microglial cells of the CNS are bone marrow-derived and present antigen in vivo // Science. — 1988. — № 239. — С. 290-292.
- ↑ Fabry Z, Sandor M, Gajewski TF, Herlein JA, Waldschmidt MM, Lynch RG, Hart MN. Differential activation of Th1 and Th2 CD4+ cells by murine brain microvessel endothelial cells and smooth muscle/pericytes // J Immunol. — 1993. — № 151. — С. 38-47.
- ↑ Krause D, Kunz J, Dermietzel R. Cerebral pericytes - a second line of defense in controlling blood-brain barrier peptide metabolism // Adv Exp Med Biol. — 1993. — № 331. — С. 149-152.
- ↑ Thomas WE. Brain macrophages: on the role of pericytes and perivascular cells // Brain Res Brain Res Rev. — 1999. — № 31. — С. 42-57.
- ↑ Iadecola C. Neurovascular regulation in the normal brain and in Alzheimer's disease // Nat Rev Neurosci. — 2004. — № 5. — С. 347-360.
- ↑ Johanson CE. Permeability and vascularity of the developing brain: cerebellum vs cerebral cortex // Brain Res. — 2004. — № 190. — С. 3–16.
- ↑ Neuhaus J, Risau W, Wolburg H. Induction of blood-brain barrier characteristics in bovine brain endothelial cells by rat astroglial cells in transfilter coculture // Ann N Y Acad Sci. — 1991. — № 633. — С. 578–580.
- ↑ Stewart PA, Wiley MJ. Developing nervous tissue induces formation of blood-brain barrier characteristics in invading endothelial cells: a study using quail–chick transplantation chimeras // Dev Biol.. — 1981. — № 84. — С. 183–192.
- ↑ Raub TJ, Kuentzel SL, Sawada GA. Permeability of bovine brain microvessel endothelial cells in vitro: barrier tightening by a factor released from astroglioma cells // Exp Cell Res. — 1992. — № 199. — С. 330–340.
- ↑ 1 2 3 4 Abbott NJ. Astrocyte-endothelial interactions and blood-brain barrier permeability // J Anat.. — 2002. — № 200. — С. 629–638.
- ↑ Paulson OB, Newman EA. Does the release of potassium from astrocyte endfeet regulate cerebral blood flow? // Science. — 1987. — № 237. — С. 896-898.
- ↑ Abbott NJ, Rönnbäck L, Hansson E. Astrocyte-endothelial interactions at the blood-brain barrier // Nat Rev Neurosci. — 2006. — № 7. — С. 41–53.
- ↑ Björkhem I, Meaney S. Brain cholesterol: long secret life behind a barrier. // Arterioscler Thromb Vasc Biol. — 2004. — № 24. — С. 806-815.
- ↑ Синельников Р.Д., Синельников Я.Р. Атлас анатомии человека в 4 томах. Т.4.. — М.:: Медицина, 1996. — С. 82. — 320 с. — ISBN 5-225-02723-7
- ↑ Синельников Р.Д., Синельников Я.Р. Атлас анатомии человека в 4 томах. Т.4.. — М.:: Медицина, 1996. — С. 56. — 320 с. — ISBN 5-225-02723-7
- ↑ Duvernoy HM, Risold PY. The circumventricular organs: an atlas of comparative anatomy and vascularization // Brain Res Rev. — 2007. — № 56. — С. 119-147.
- ↑ C. Lohmann Die Blut-Hirn-Schranke in vitro: Regulation der Permeabilität durch Matrixmetalloproteasen // Диссертация. Вестфальский университет имени Вильгельма. — 2003.
- ↑ W. M. Pardridge Peptide Drug Delivery to the Brain. — Raven Press, 1991. — С. 123. — ISBN 0-88167-793-0
- ↑ Chiou WL, Barve A. Linear correlation of the fraction of oral dose absorbed of 64 drugs between humans and rats // Pharm Res. — 1998. — № 15. — С. 1792-1795.
- ↑ Goodwin JT, Clark DE. In silico predictions of blood-brain barrier penetration: considerations to "keep in mind" // J Pharmacol Exp Ther. — 2005. — № 315. — С. 477-483.
- ↑ Lindstedt L, Schaeffer PJ. Use of allometry in predicting anatomical and physiological parameters of mammals // Lab Anim. — 2002. — № 36. — С. 1-19.
- ↑ Lindstedt L, Schaeffer PJ A proposed blood circulation model for Reference Man // Health Phys. — 1995. — № 69. — С. 187-201.
- ↑ Willmann S, Schmitt W, Keldenich J, Lippert J, Dressman JB. A physiological model for the estimation of the fraction dose absorbed in humans // J Med Chem. — 2004. — № 47. — С. 4022-4031.
- ↑ Fagerholm U, Johansson M, Lennernäs H Comparison between permeability coefficients in rat and human jejunum // J Med Chem. — 1996. — № 13. — С. 1336-1342.
- ↑ Leggett RW, Williams LR. Suggested reference values for regional blood volumes in humans // Health Phys. — 1991. — № 60. — С. 139-154.
- ↑ G. B. Wislocki Experimental studies on fetal absorption. I. The vitally stained fetus // Contrib Embryol Carnegie Inst. — 1920. — № 5. — С. 45-52.
- ↑ Wakai S, Hirokawa N. Development of the blood-brain barrier to horseradish peroxidase in the chick embryo // Cell Tissue Res. — 1978. — № 195. — С. 195-203.
- ↑ Risau W, Hallmann R, Albrecht U. Differentiation-dependent expression of proteins in brain endothelium during development of the blood-brain barrier // Dev Biol.. — 1986. — № 117. — С. 537-545.
- ↑ Reynolds ML, Evans CA, Reynolds EO, Saunders NR, Durbin GM, Wigglesworth JS. Intracranial haemorrhage in the preterm sheep fetus // Early Hum Dev. — 1979. — № 3. — С. 163-186.
- ↑ L. Stern, R. Peyrot Le fonctionnement de la barrière hémato-éncephalique aux divers stades de développement chez les diverses espèces animales // Compte Rendu des Societe de Biologie (Paris). — 1927. — № 96. — С. 1124–1126.
- ↑ L. Stern et al Le fonctionnement de la barrière hémato-éncephalique aux divers stades de développement chez les diverses espèces animales // Compte Rendu Soc Biol. — 1929. — № 100. — С. 231–233.
- ↑ Saunders NR, Habgood MD, Dziegielewska KM. Barrier mechanisms in the brain, II. Immature brain // Clin Exp Pharmacol Physiol. — 1999. — № 26. — С. 85–91.
- ↑ 1 2 N. R. Saunders Development of the blood–brain barrier to macromolecules // The Fluids and Barriers of the Eye and Brain / M. B. Segal. — Verlag MacMillan. — Raven Press, 1991. — С. 128-155. — ISBN 0-8493-7707-2
- ↑ Schumacher U, Mollgård K. The multidrug-resistance P-glycoprotein (Pgp, MDR1) is an early marker of blood-brain barrier development in the microvessels of the developing human brain // Histochem Cell Biol. — 1997. — № 108. — С. 179–182.
- ↑ Dziegielewska KM, Evans CA, Malinowska DH, Møllgård K, Reynolds JM, Reynolds ML, Saunders NR. Studies of the development of brain barrier systems to lipid insoluble molecules in fetal sheep // J Physiol. — 1979. — № 292. — С. 207–231.
- ↑ Ferguson RK, Woodbury DM. Penetration of 14C-inulin and 14C-sucrose into brain, cerebrospinal fluid and skeletal muscle of developing rats // Exp Brain Res. — 1969. — № 7. — С. 181–194.
- ↑ Habgood MD, Knott GW, Dziegielewska KM, Saunders NR. The nature of the decrease in blood-cerebrospinal fluid barrier exchange during postnatal brain development in the rat // J Physiol. — 1993. — № 468. — С. 73–83.
- ↑ C. E. Johanson Ontogeny of the blood–brain barrier // Implications of the Blood–Brain Barrier and Its Manipulation / E. A. Neuwelt. — Plenum Press, 1989. — С. 157-198.
- ↑ Braun LD, Cornford EM, Oldendorf WH. Newborn rabbit blood-brain barrier is selectively permeable and differs substantially from the adult // J Neurochem. — 1980. — № 34. — С. 147–152.
- ↑ Cornford EM, Braun LD, Oldendorf WH. Developmental modulations of blood–brain barrier permeability as an indicator of changing nutritional requirements in the brain // Pediatr Res. — 1982. — № 16. — С. 324–328.
- ↑ Brenton DP, Gardiner RM. Transport of L-phenylalanine and related amino acids at the ovine blood-brain barrier // J Physiol. — 1988. — № 402. — С. 497–514.
- ↑ Frank HJ, Jankovic-Vokes T, Pardridge WM, Morris WL. Enhanced insulin binding to blood–brain barrier in vivo and to brain microvessels in vitro in newborn rabbits // Diabetes. — 1985. — № 34. — С. 728–733.
- ↑ Saunders NR, Knott GW, Dziegielewska KM. Barriers in the immature brain // Cell Mol Neurobiol. — 2000. — № 20. — С. 29–40.
- ↑ Abbott NJ, Bundgaard M Electron-dense tracer evidence for a blood-brain barrier in the cuttlefish Sepia officinalis // J Neurocytol. — 1992. — № 21. — С. 276–294.
- ↑ Abbott NJ, Pichon Y The glial blood-brain barrier of crustacea and cephalopods: a review // J Physiol (Paris). — 1982. — № 21. — С. 304–313.
- ↑ 1 2 Abbott NJ. Dynamics of CNS barriers: evolution, differentiation, and modulation // Cell Mol Neurobiol. — 2005. — № 25. — С. 5–23.
- ↑ N. J. Abbott Comparative physiology of the blood-brain barrier // Physiology and pharmacology of the bloodbrain barrier / M. W. B. Bradbury. — Springer-Verlag, 1992. — С. 371-396. — ISBN 0-387-54492-5
- ↑ N. Hettenbach Einfluss chronischer elektromagnetischer Befeldung mit Mobilfunkstrahlen (GSM und UMTS) auf die Integrität der Blut-Hirn-Schranke von Ratten // Диссертация. Мюнхенский университет Людвига-Максимилиана. — 2008.
- ↑ S. I. Rapoport Blood-brain Barrier in Physiology and Medicine. — Raven Press, 1976. — ISBN 0-89004-079-6
- ↑ M. Fromm Physiologie des Menschen // Transport in Membranen und Epithelien / R. F. Schmidt, F. Lang. — Verlag Springer. — С. 41-54. — ISBN 978-3-540-32908-4
- ↑ I. Sauer Apolipoprotein E abgeleitete Peptide als Vektoren zur Ьberwindung der Blut-Hirn-Schranke // Диссертация. Свободный университет Берлина. — 2004.
- ↑ Egleton RD, Davis TP Development of neuropeptide drugs that cross the blood-brain barrier // NeuroRx. — 2005. — № 2. — С. 44-53.
- ↑ Oldendorf WH Lipid solubility and drug penetration of the blood brain barrier // Proc Soc Exp Biol Med. — 1974. — № 147. — С. 813-815.
- ↑ 1 2 R. Kaliszan, M. Markuszewski Brain/blood distribution described by a combination of partition coefficient and molecular mass // International Journal of Pharmaceutics. — 1996. — № 145. — С. 9-16.
- ↑ Träuble H Carriers and specificity in membranes. 3. Carrier-facilitated transport. Kinks as carriers in membranes // Neurosci Res Program Bull. — 1971. — № 9. — С. 361-372.
- ↑ Träuble H Phase transitions in lipids. Possible switch processes in biological membranes // Naturwissenschaften. — 1971. — № 58. — С. 277-284.
- ↑ O. Vostowsky Chemie der Naturstoffe - Lipoproteine und Membranen // Эрлангенский университет. — 2005. — № 58. — С. 42.
- ↑ W. Hoppe, R. D. Bauer Biophysik. — Verlag Birkhäuser, 1982. — С. 447-448. — ISBN 0-387-11335-5
- ↑ Seelig A, Seelig J. The dynamic structure of fatty acyl chains in a phospholipid bilayer measured by deuterium magnetic resonance // Biochemistry. — 1974. — № 13. — С. 4839-4845.
- ↑ A. Elbert Die Permeation kleiner polarer Moleküle durch Phospholipidmodellmembranen // Диссертация. Университет Кайзерслаутерна. — 1999.
- ↑ Seelig A, Gottschlich R, Devant RM A method to determine the ability of drugs to diffuse through the blood-brain barrier // Proc Natl Acad Sci U S A. — 1994. — № 91. — С. 68-72.
- ↑ Dhopeshwarkar GA, Mead JF Uptake and transport of fatty acids into the brain and the role of the blood-brain barrier system // Adv Lipid Res. — 1973. — № 11. — С. 109-142.
- ↑ Gerebtzoff G, Seelig A In silico prediction of blood-brain barrier permeation using the calculated molecular cross-sectional area as main parameter // J Chem Inf Model. — 2006. — № 46. — С. 2638-2650.
- ↑ Seelig A, Gottschlich R, Devant RM A method to determine the ability of drugs to diffuse through the blood-brain barrier // Proc Natl Acad Sci USA. — 1994. — № 91. — С. 68-72.
- ↑ Pardridge WM The blood-brain barrier: bottleneck in brain drug development // NeuroRx. — 2005. — № 2. — С. 3-14.
Wikimedia Foundation. 2010.
ГЕМАТОЭНЦЕФАЛИЧЕСКИЙ БАРЬЕР — (от гемо... и греч. enkephalos мозг) физиологический механизм, регулирующий обмен веществ между кровью, спинномозговой жидкостью и мозгом. Защищает центральную нервную систему от проникновения чужеродных веществ, введенных в кровь, или продуктов… … Большой Энциклопедический словарь
ГЕМАТОЭНЦЕФАЛИЧЕСКИЙ БАРЬЕР — ГЕМАТОЭНЦЕФАЛИЧЕСКИЙ БАРЬЕР, механизм, предупреждающий попадание вредных веществ в мозг с током крови. Этот механизм включает различные способы защиты, в том числе, избирательную проницаемость клеток, которые предупреждают попадание «посторонних» … Научно-технический энциклопедический словарь
Гематоэнцефалический барьер — (греч. haima – кровь, enkephalos мозг) избирательно проницаемый для химических субстанций барьер, расположенный между кровью и тканями мозга. Отчасти образован, предположительно, клетками сосудистых капилляров в головном мозге, рост которых… … Энциклопедический словарь по психологии и педагогике
гематоэнцефалический барьер — (от гемо... и греч. enképhalos мозг), физиологический механизм, регулирующий обмен веществ между кровью, спинномозговой жидкостью и мозгом. Защищает центральную нервную систему от проникновения чужеродных веществ, введённых в кровь, или… … Энциклопедический словарь
ГЕМАТОЭНЦЕФАЛИЧЕСКИЙ БАРЬЕР — Избирательно проницаемый барьер между циркулирующей кровью и мозгом, который предотвращает попадание некоторых веществ в мозг. Барьер, как предполагается, состоит частично из клеток, составляющих стенки капилляров мозга. Предположительно,… … Толковый словарь по психологии
гематоэнцефалический барьер — (гемато + анат. encephalon головной мозг) гистогематический барьер между кровью, с одной стороны, и цереброспинальной жидкостью и нервной тканью с другой … Большой медицинский словарь
Гематоэнцефалический барьер — I Гематоэнцефалический барьер физиологический механизм, избирательно регулирующий обмен веществ между кровью, цереброспинальной жидкостью и центральной нервной системой и обеспечивающий постоянство внутренней среды головного и спинного мозга. Г.… … Медицинская энциклопедия
ГЕМАТОЭНЦЕФАЛИЧЕСКИЙ БАРЬЕР — (от гемо... и мозг), физиол. механизм, регулирующий обмен в в между кровью, спинномозговой жидкостью и мозгом. Защищает центр. нерв. систему от проникновения чужеродных в в, введённых в кровь, или продуктов нарушенного обмена в в … Естествознание. Энциклопедический словарь
Барьер Гематоэнцефалический (Blood-Brain Barrier) — механизм, с помощью которого циркулирующая в теле человека кровь не смешивается с жидкими тканями, окружающими клетки мозга. Гематоэнцефалический барьер является полупроницаемой мембраной, так как обеспечивает прохождение через него жидкостей и… … Медицинские термины
БАРЬЕР ГЕМАТОЭНЦЕФАЛИЧЕСКИЙ — (blood brain barrier) механизм, с помощью которого циркулирующая в теле человека кровь не смешивается с жидкими тканями, окружающими клетки мозга. Гематоэнцефалический барьер является полупроницаемой мембраной, так как обеспечивает прохождение… … Толковый словарь по медицине
Гематоэнцефалический барьер











18+
© Академик, 2000-2024
- Обратная связь: Техподдержка, Реклама на сайте
Экспорт словарей на сайты, сделанные на PHP, Joomla, Drupal, WordPress, MODx.