- Размножение ланцетников
-
Размножение ланцетников — примитивных морских животных из семейства ланцетниковых (лат. Branchiostomidae) — и их последующее эмбриональное развитие является примером для рассмотрения эмбриогенеза хордовых в целом. Изучение онтогенеза ланцетника имеет значение для филогенетики, в особенности для истории происхождения позвоночных животных, а также нахождения филогенетических связей между различными таксонами животных.
Содержание
История изучения
Ранние исследования развития ланцетника, в частности работы А. О. Ковалевского (1865, 1867), доказывали единство животного мира и поэтому имели большое значение для утверждения эволюционной теории. Эти работы послужили стимулом к изучению ланцетника для освещения филогенетических связей между животными. Первыми, это, казалось бы, невзрачное животное изучали такие учёные как Бертольд Гатчек (1882, 1893), Бовери (1892), Эдвин Г. Конклин (1932), Эдвин С. Гудрич (1934) и другие.
В первом экспериментальном изучении проблемы филогенетических связей ланцетника, Эдмунд Б. Вильсон (1893) исследовал развитие изолированных бластомеров и неполных эмбрионов. Он заключил, что регулятивная способность ланцетников — нечто среднее между ней же у морских ежей и асцидий.
До недавнего времени исследования эмбриогенеза ланцетников до первого деления были поверхностными из-за некоторых существенных трудностей с искусственным оплодотворением, и исследуемые эмбрионы собирались после естественного оплодотворения.[1] Таким образом, ранние стадии эмбриогенза, такие как кортикальные реакции, движение пронуклеусов и первое деление не были основаны на полноценных материалах.
Размножение
Ланцетники — раздельнополые животные. В популяции число мужских и женских особей одинаково.[2]
Размножаются ланцетники в весенее, летнее или осеннее время. Сразу же после захода солнца самки начинают метать зрелые икринки (яйца).Так как половых протоков нет, созревшие половые клетки выходят в атриальную полость через разорванные стенки гонад[3] и стенки тела[4], а уже оттуда попадают в воду.
Прогенез
Проэмбриональное развитие, предзиготный период или прогенез — образование и формирование родительских половых клеток, которые сформируют зиготу.
Яйцо
Яйца ланцетников принадлежат к изолецитальному и олиголецитальному типам — содержат мало желтка, который относительно равномерно распределён в цитоплазме. Яйцеклетки очень мелки (100—120 мкм).[5]
В ооците выделяют анимальный и вегетативный полюс.[5] Развивающиеся ооциты анимальным полюсом крепятся к стенкам яичника и покрыты фолликулярным эпителием.[6] В удалённом от центра слое цитоплазмы ооцита нет желтка, но содержатся красящиеся гранулы, возможно это митохондрии. Диаметр ядра ооцита (зародышевый пузырёк) равен половине диаметра всего ооцита и располагается эксцентрически — ближе к анимальному полюсу, в свободном от желточных зёрен месте. Впоследствии это место становится анимальным полюсом яйца.
Кроме того, в области анимального полюса при созревании яйцеклетки, ещё до проникновения сперматозоида и образования оболочки оплодотворения, отделяется первое редукционное тельце. Оно уносится водой и погибает.[5] После проникновения сперматозоида в яйцеклетку вокруг неё образуется оболочка оплодотворения, не дающая проникнуть в яйцо другим спермиям. Сразу же отделяется второе редукционное тельце, которое лежит между желточной оболочкой и самой яйцеклеткой.[5] По расположению второго тельца можно установить где находится анимальный полюс.[6] С проникновением спермия заканчивается мейоз яйцеклетки.
Оплодотворение
Оплодотворение наружное, моноспермное (в одну яйцеклетку попадает один сперматозоид).
Перед делениями созревания в яйце исчезает оболочка ядра и ядерный сок попадает в цитоплазму. Так, в середине яйца образуется бедная желтком миксоплазма. Сперматозоид проникает в яйцеклетку на стадии метафазы 2—го деления возле вегетативного полюса, определяя вентральную сторону зиготы. Ядро сперматозоида с центросомой продвигаются к сформированному женскому пронуклеусу. При этом на дорсальной стороне зиготы, несколько ниже главной оси яйца, образуется серый (мезодермальный) серп — скопление безжелтковой периферической цитоплазмы, будущая спинная губа бластопора[7], одна из частей будущей мезодермы. Появление этого серпа связывается с маркированием участка, где впоследствии появится спинная губа бластопора и перераспределением материала в яйце. Движение материала зиготы определяет вегетативный полюс зародыша, так как белково-липидные включения скапливаются у этой области зиготы.
Оба пронуклеуса лежат в светлой цитоплазме, из которой позже формируется веретено первого деления дробления. Она имеет вид перевёрнутой чаши и нижним краем прикасается с мезодермальным серпом. Во время дробления светлая цитоплазма попадает преимущественно в клетки будущей эктодермы на анимальном полюсе яйца, так как концентрируется вокруг ядер бластомеров. Желток же попадает в клетки энтодермы, так как занимает центральную и вегетативную части яйца.[6]
Из этого следует, что в яйце ланцетника ещё до начала дробления цитоплазматический материал дифференцирован по трём зародышевым листкам. Более того, в структуре яйца уже имеется главная ось и проявляется билатеральная симметрия.[6] Все эти признаки проявляются благодаря наличию мезодермального серпа, середина которого представляет будущий задний конец эмбриона. Благодаря ему же можно определить и все стороны зародыша, так, например, бо́льшая часть анимальной области яйца соответствует брюшной стороне, бо́льшая часть вегетативной поверхности — дорсальной.
Последующий онтогенез животного также происходит в воде.
Эмбриональное развитие
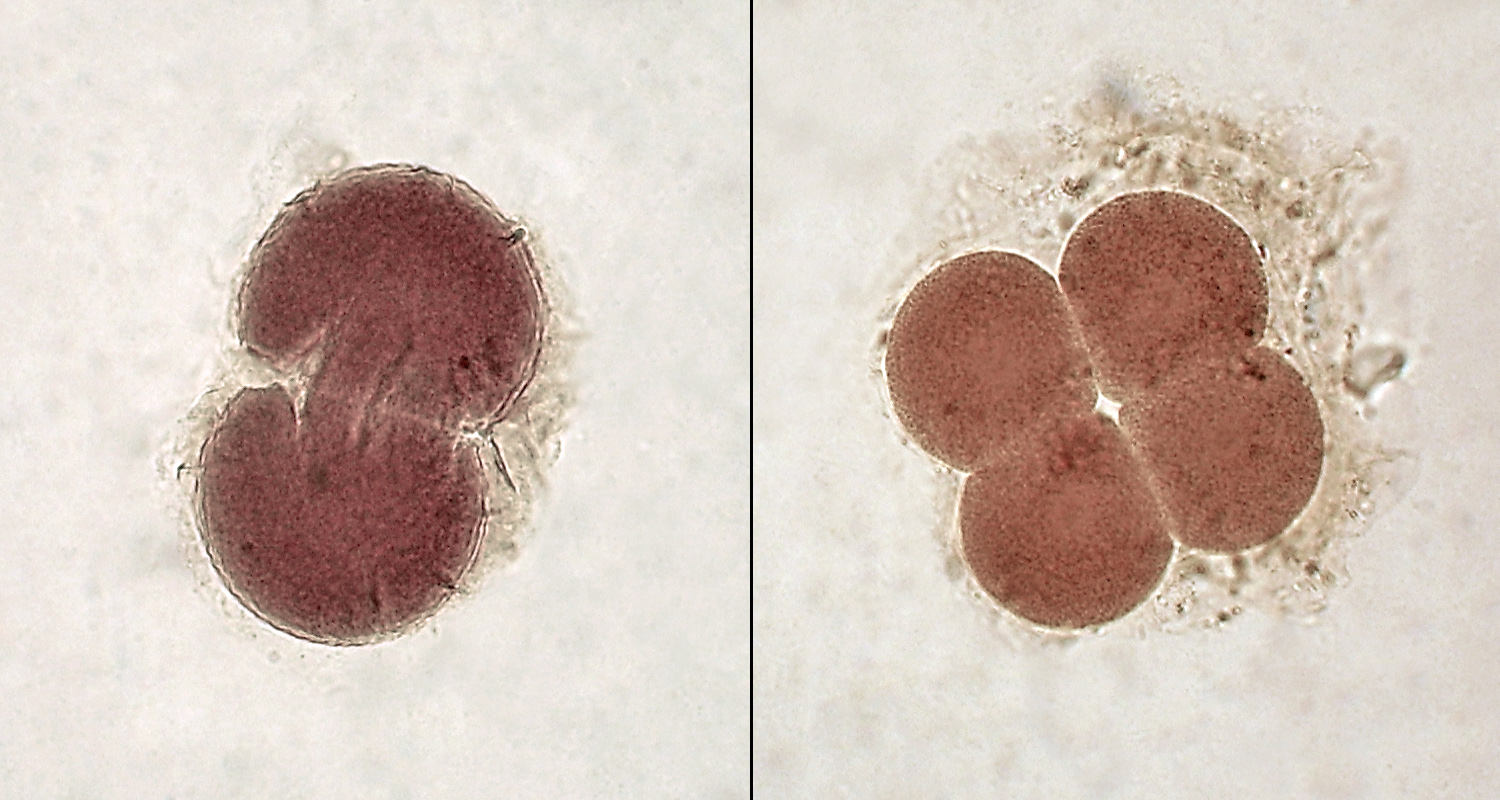
 Эмбрионы ланцетника на стадии двух и четырёх бластомеров
Эмбрионы ланцетника на стадии двух и четырёх бластомеров
Дробление. Благодаря малому количеству желтка, первый период онтогенеза проходит легко — дробление голобластическое (полное), бластомеры почти одинаковой величины (не вполне равномерное дробление), симметрия дробящегося яйца радиальная с элементами билатеральной симметрии. Редко можно наблюдать слабые признаки спирального дробления.[6]
Обычно борозда первого деления проходит через анимальный и вегетативный полюса и через середину мезодермального серпа. Таким образом яйцо разделяется на две симметричные половины. Вторая борозда дробления тоже меридиональная, перпендикулярна борозде первого деления и отделяет два анимальных (заднебрюшных) бластомера от двух вегетативных (переднеспинных), при чём первые содержат материал мезодермального серпа и несколько меньше вторых.[6] Третье деление проходит чуть выше экватора и образует четыре анимальных и вегетативных микромера, при этом два задних несколько мельче передних. При четвёртом, приблизительно меридиональном делении проявляется билатеральность — из восьми вегетативных бластомеров два передних и два задних мельче оставшихся боковых. Пятое деление — широтное, шестое деление происходит в меридиональном направлении. После шестого деления бластомеры располагаются неправильно, а уже при седьмом они делятся в разных направлениях.
Дробление становиться не синхронным и не правильным с восьмого деления. Медленне всего делятся крупные энтодермальные бластомеры, а быстрее всего — мелкие мезодермальные.
В итоге образуется целобластула, которой характерна большая полость и однослойная бластодерма. Бластоцель появляется на стадии 4—8 бластомеров. Изначально он открыт на обоих полюсах яйца, но потом эти отверстия замыкаются. Увеличение бластулы и расширение бластоцеля происходит на счёт набухания студенистого вещества в последнем. Позже, при максимальной величине бластоцеля, это вещество разжижается практически до плотности морской воды.
Гаструляция. Внутренний слой клеток образуется путем впячивания внутрь полости бластулы клеток её стенки — инвагинации. Постепенно происходит смыкание краёв бластопора и удлинение зародыша за счёт выпячивания тела в передне-заднем направлении и уменьшения поперечника гаструлы. Эмбрион становится билатерально симметричным.
Нейруляция. По краю нервной пластинки — образования клеток эктодермы — нарастают направленные вверх складки (нервные валики), центр же опускается, формируя нервный желобок.
Вместе с этим происходит расщепление мезодермы в форме небольших карманообразных выростов по двум сторонам энтодермы. Помимо продольных желобков от переднего конца первичной кишки (гастроцеля) по порядку отделяются дополнительные две пары целомических мешков. Это означает, что стадия характеризуется тремя парами сегментов и указывает на филогенетическую родственную связь рода ланцетников с трёхсегментарными личинками полухордовых и иглокожих.[5]
Образование целома — отшнуровка от первичного кишечника — происходит энтероцельно. Мезодермальные тяжи полностью распадаются на сегменты — происходит дифференцировка сомитов (спинная часть) и спланхнотом (брюшная). Сомиты, из которых развивается туловищная мускулатура, остаются обособленными друг от друга, спланхнотомы же сливаются на каждой стороне, образуя левую и правую полости, которые затем объединяются под кишечной трубкой в общую вторичную полость тела — целом.[5]
Эмбрион ланцетника, с одной стороны, обладает чертами типичных позвоночных:
- присущее им расположение зачатков органов при гаструляции,
- отделение хорды из спинной стенки гастроцеля,
- образование нервной пластинки из эктодермы с дорсальной стороны зародыша.
С другой стороны, у них наблюдаются признаки беспозвоночных вторичноротых:
- присутствие целобластулы,
- образование гаструлы путём впячивания внутрь (инвагинация),
- трёхсегментная стадия,
- энтероцельный способ образования целома.
Постэмбриональное развитие
Через 4—5 дней[5] после оплодотворения из икры выходит личинка, имеющая очень малые размеры, которая начинает самостоятельно питаться. Некоторое время личинка плавает при помощи покрывающих тело ресничек, затем опускается на дно, где развивается с метаморфозом. Личинки, достигшие длины в 3,5—5,2 мм, поднимаются ночью к верхним слоям воды, а днём опускаются ко дну. Ланцетники пребывают в личиночной стадии около трёх месяцев.[8][9]
Личинки ланцетников особенно интересны, так как обладают некоторыми любопытными особенностями : иногда они могут вырасти в так называемых гигантских «амфиоксидов» (англ. giant «amphioxides» form), а также обладают право-левой асимметрией (в то время как большинству животных присуща лево-правая асимметрия[10]). Кроме того, личинки ланцетников составляют значительную часть запаса зоопланктона Атлантического океана[11] и, соответственно, являются важным пищевым источником для морских животных.
Передвижение личинок
 Парящая четырёхдневная личинка. (Масшатабная линия — 0,5 мм.)
Парящая четырёхдневная личинка. (Масшатабная линия — 0,5 мм.)Передвижение двухдневных эмбрионов осуществляется спиральными, ресничными плавательными движениями.[12]
Месячные дометаморфные личинки проводят большинство времени практически неподвижно вися в средних слоях воды. Тело каждой парящей личинки ориентировано под углом примерно в 60° по горизонтали, при этом тело наклонено так, что передний конец и брюшная сторона ориентированы по направлению к поверхности воды.[13] Движущая сила парения — метахрональные волнообразные колебания эпидермальных ресничек. Ресничные волны проходят от переднего до заднего конца тела со скоростью 0,3 мм/с.[13] «Парящие» личинки поддерживают им характерное наклонённое положение в темноте и умеренном свете. Однако, они проявляют некоторую направленную светочувствительность: они медленно ориентируют головы от смещенного в сторону от центра источника света. Поскольку полная ориентация требует около 20 минут, трудно понять какую (если таковая вообще имеются) функцию может иметь это явление. Однако, такая светочувствительность указывает на то, что личинки имеют направительные фоторецепторы. Эти фоторецепторы могут представлять собой нейроны, связанные с пигментным пятном на переднем конце мозгового пузырька.[14]
Как показали исследования Стоукса и Холланда[13], не все личинки ланцетников передвигаются за счёт волнообразных колебаний ресничек. Они способны плавать при помощи мускульной ундуляции в течение нескольких секунд и с редкими интервалами, или даже ползать по дну с помощью эпидермальных ресничек. Но такие типы передвижения скорее всего связаны с плохими условиями содержания, нежели с особенностями развития.[13]
В отличие от волнообразного (ундуляторного) передвижения, «парение», энергетически эффективнее: количество энергии — примерно 10−9—10−11 W (в зависимости от возраста личинки), в то время как волнообразное движение требует около 10−6—10−8 W.[15] Ресничное передвижение благоприятствует непрерывной фильтрации. Также неподвижные личинки менее привлекательны для хищников, что обеспечивает обеспечивает ланцетникам большую выживаемость на ранних стадиях личиночного развития. Большинство ресничек личинки теряют во время метаморфоза.[16]
Гигантские личинки-амфиоксиды
Гигантские личинки ланцетников — феномен, редко встречающийся в морском зоопланктоне.[17] Личинки-амфиоксиды распространены во всех открытых частях Индийского, Атлантического и Тихого океанов, даже над большими, до 3000 м, глубинами, но только в тех районах, куда они могут попасть с течением.[8]
Первоначально их принимали за взрослых особей и выделяли в отдельный род (лат. Amphioxides), но в настоящее время они отнесены к личинкам.[18] Так, например, Amphioxides Gill, 1895 является на самом деле гигантской личинкой Epigonichthys Peters, 1876.[19]
Личинки, которые переживают метаморфоз, обычно, в зависимости от окружающей среды, 3,5—5 мм длиной и имеют 12—14 жаберных щелей. Личинки-амфиоксиды в этот же период намного больше — примерно 11 мм, и имеют большее количество жаберных щелей — до 35.[18] Амфиоксиды не встречаются у дна, как взрослые ланцетники, а только в планктоне. Ротовая полость у личинок-амфиоксидов расположена на левой стороне тела, практически нет ротовых щупалец, атриальная полость отсутствует, половые железы не развиты — это характерные для личинок черты строения.[8]
Чтобы выяснить факторы, которые вызывают продолжительное личиночное существование, были проведены исследования. Например, велись наблюдения за метаморфозом у личинок ланцетников в различных концентрациях тиоурацила — гормона, вызывающего метаморфоз у позвоночных, насыщенных растворах и суспензиях тироксина и трийодотиронина, а также в обычной морской воде.[17] Были сделаны выводы, что метаморфоз — вызываемый и самоподдерживающийся процесс.[17] Таким образом, однажды начавшись, он может существенно замедлится или ускориться. Это может объяснить феномен гигантских личинок.
Примечания
- ↑ Holland and Holland (1992).
- ↑ См. описание на сайте «JRank». (англ.)
- ↑ Описано в учебнике Н. П. Наумова и Н. Н. Карташёва (1979).
- ↑ См., например, учебное пособие Е. А. Веселова и О. Н. Кузнецовой (1979).
- ↑ 1 2 3 4 5 6 7 Данные из методического указания Г. Т. Масловой и А. В. Сидорова (2005).
- ↑ 1 2 3 4 5 6 По Ивановой-Казас (1978).
- ↑ Из лекции на виртуальном сервере по эмбриологии и цитологии «WEBEMBRYO».
- ↑ 1 2 3 Информация из книги «Жизнь животных» (1971).
- ↑ Сведения из учебного пособия С. П. Гапонова (2004).
- ↑ Levin (2004).
- ↑ Jefferies (1986).
- ↑ Bone (1958).
- ↑ 1 2 3 4 Stokes and Holland (1995b).
- ↑ Lacalli et al. (1994).
- ↑ Stokes (1997).
- ↑ Stokes and Holland (1995a).
- ↑ 1 2 3 Wickstead (1967).
- ↑ 1 2 Из публикации о семействе ланцетниковых на сайте «ICES». (англ.)
- ↑ Таксономия на сайте ITIS (англ.)
Литература
- Веселов Е. А., Кузнецова О. Н. Практикум по зоологии: учеб. пособие для с.-х. вузов / Е. А. Веселов, О. Н. Кузнецова. — З-е изд., доп. — М.: Высшая школа, 1979. — С. 177—183.
- Гапонов С. П. Оболочники. Бесчерепные. Развитие хордовых // Пособие по курсу «Зоология позвоночных»: учеб. пособие для студентов 2-го курса биолого-почвенного факультета / С. П. Гапонов. — Воронеж: Изд-во ВГУ, 2004. — С. 26.
- Жизнь животных. Т. 4. Рыбы / Под ред. проф. Т. С. Расса. — М.: Просвещение, 1971. — Ч. 1. — С. 12—14.
- Иванова-Казас О. М. Сравнительная эмбриология беспозвоночных животных: Низшие хордовые. — М.: Наука, 1978. — С. 4—133.
- Маслова Г. Т., Сидоров А. В. Развитие позвоночных. Ланцетник: метод. указания по курсу «Биология индивидуального развития». — Мн.: БГУ, 2005. — С. 13.
- Наумов Н. П., Карташёв Н. Н. Низшие хордовые, бесчелюстные, рыбы, земноводные // Зоология позвоночных: учебник для биолог. спец. ун-тов / Н. П. Наумов, Н. Н. Карташёв. — М.: Высш. школа, 1979. — Ч. 1. — С. 33—42.
- Bone Q. Observations upon the living larva of amphioxus // Pubbl. Staz. Zool. Napoli. — 1958. — T. 30. — P. 458—441. (англ.)
- Holland L. Z. and Holland N. D.. Early Development in the Lancelet (= Amphioxus) Branchiostoma floridae from Sperm Entry through Pronuclear Fusion: Presence of Vegetal Pole Plasm and Lack of Conspicuous Ooplasmic Segregation// The Biological Bulletin. — 1992. — Vol. 182. Issue 1. — P. 77—96. (англ.)
- Jefferies R. P. S. The Ancestry of the Vertebrates. — London: British Museum (Natural History), 1986. — 376 p. (англ.)
- Lacalli T. C., Holland N. D., West J. E. Landmarks in the anterior central nervous system of amphioxus larvae // Philosophical Transactions: Biological Sciences. — 1994. — Vol. 344. — P. 165—185. (англ.)
- Levin M. The embryonic origins of left-right asymmetry // Critical Reviews in Oral Biology and Medicine. — 2004. — Vol. 15. No. 4. — P. 197—206. (англ.)
- Stokes M. D. Larval locomotion of the lancelet Branchiostoma floridae // Journal of Experimental Biology. — 1997. — Vol 200. — P. 1661—1680. (англ.)
- Stokes M. D., Holland N. D. Ciliary hovering in larval lancelets (=amphioxus) // Biol. Bull. — 1995b. — Vol. 188. — P. 23l—233. (англ.)
- Wickstead J. H. Branchiostoma Lancelatum larvae: some experiments on the effect of thiouracil on the metamorphosis // J. mar. biol. Ass. U. K. — 1967. — Vol. 47. — P. 49—59. (англ.)
- Stokes M. D., Holland N. D. Embryos and larvae of a lancelet, Branchiostoma floridae, from hatching to metamorphosis: growth in the laboratory and external morphology // Acta Zool. Stockh. — 1995a. — Vol. 76. — P. 105—120. (англ.)
См. также
Ссылки
- «Lancelets: Cephalochordata — Physical Characteristics, Behavior and Reproduction, Lancelets and Vertebrates, Florida Lancelet (Branchiostoma floridae): Species Account — Geographic Range, Habitat, Diet, Conservation Status» — описание флоридского вида ланцетника на сайте «JRank» (англ.)
- «Chordata, Sub-Phylum Acrania (=Cephalochordata), Family: Branchiostomidae (By J. H. Wickstead) 1967» — публикация о семействе ланцетниковых на сайте «ICES» (Дания) (англ.)

 Категория:
Категория:- Размножение

Wikimedia Foundation. 2010.