- Птицы
-
Запрос «Птица» перенаправляется сюда; см. также другие значения.
Птицы 
18 видов птицНаучная классификация промежуточные рангиДомен: Эукариоты Царство: Животные Подцарство: Эуметазои Без ранга: Двусторонне-симметричные Без ранга: Вторичноротые Тип: Хордовые Подтип: Позвоночные Инфратип: Челюстноротые Надкласс: Четвероногие Класс: Птицы Международное научное название Подклассы - Бескилевые (Paleognathae)
- Новонёбные (Neognathae)
Систематика
на Викивидах
Изображения
на ВикискладеITIS 174371 NCBI 8782 EOL 695 Пти́цы (лат. Aves) — класс теплокровных яйцекладущих позвоночных животных, представители которого характеризуются тем, что тело их покрыто перьями и передние конечности видоизменены в органы полёта — крылья (виды, которые не летают, имеют недоразвитые крылья)[1] Изначально строение тела птиц приспособлено к полёту, хотя в настоящее время существует много видов нелетающих птиц. Ещё одним отличительным признаком птиц является также наличие клюва. На сегодняшний день на Земле обитает более 9792 различных видов[2], в том числе на территории России — 657 гнездящихся видов, а всего на территории страны отмечено 789 видов, включая 125 видов, гнездование которых не показано, и 7 вымерших видов[3], что делает их наиболее разнообразной группой надкласса четвероногих. Птицы населяют все экосистемы Земного шара от Арктики до Антарктики. Наука, изучающая птиц, называется орнитология.
Содержание
Этимология
Русское слово «птица» восходит к праслав. *ръtica, которое является суффиксальным производным от *ръtа, которое родственно латыш. putns «птица», лит. putýtis «пташка», лат. putus «дитя», putila «птенец», оскск. рuсlо- «дитя», др.-инд. पुत्रः (putráḥ IAST) «дитя, сын», др.-инд. पोतः (рótаḥ IAST) «детеныш животного», и восходит в свою очередь к пра-и.е. *pōu-/*pəu-/*pu-/*pū- «маленький»[4][5].
Латинское слово avis восходит к пра-и.е. *h2ewis (в традиционной реконструкции *awis) «птица», от которого происходят также др.-инд. विः (viḥ IAST) «птица», др.-греч. ἀετός «орёл», арм. haw «птица», валл. hwyad «утка»[6][7].
Краткий обзор класса
 Красный лори (Eos bornea)
Красный лори (Eos bornea)
Птицы представляют собой хорошо обособленную группу теплокровных позвоночных животных. Одним из наиболее характерных признаков представителей которой является покров из перьев, предохраняющий тело от неблагоприятных изменений температуры и играющий важную роль при полёте. Способность к полёту является главнейшей особенностью птиц (отсутствие такой способности у некоторых современных птиц — вторичное явление)[8]. Все птицы откладывают яйца. Верхние конечности приобрели форму крыльев, и птицы, как правило способны летать, хотя есть и исключения — страусы, пингвины и ряд островных эндемиков в ходе эволюции утратили способность к полёту. Птицы обладают особым строением органов дыхания и пищеварения, что тесно связано с их способностью летать. Некоторые птицы, в особенности вороны и попугаи, обладают способностью к быстрому обучению, другие используют подручные «инструменты», а третьи передают накопленные ими навыки из поколения в поколение.
Многие виды птиц совершают регулярные длительные перелёты из одного региона Земли в другой, а ещё большее количество ведут кочевой образ жизни, постоянно перемещаясь на небольшие расстояния в пределах ареала. Птицы являются социальными животными, общаясь между собой с помощью визуальных и звуковых сигналов, и выполняют общественные действия — такие как коммунальное гнездование, совместная охота и защита от хищников. Для большинства видов характерная «социальная моногамия» — пары формируются на один или реже на несколько сезонов размножения, у некоторых групп птиц на всю жизнь. Среди брачных форм также встречаются полигиния (когда самец сожительствует с несколькими самками) и в редких случаях полиандрия (одна самка и несколько самцов). Как правило, яйца откладываются в гнездо и насиживаются обоими членами пары. Большинство птиц длительное время ухаживают за потомством после его появления на свет.
В связи с тем, что полёт требует больших затрат мышечной энергии, уровень обмена веществ у птиц чрезвычайно высок и потребность в пище велика: дневная норма пищи составляет 12—28 % общей массы тела[8]. Температура тела птиц в среднем составляет 42,2 °С, а у мелких видов достигает 45,5 °С[8]. Такая высокая температура тела имеет большое физиологическое значение, являясь одним из важнейших условий высокой интенсивности обмена веществ и всех физиологических процессов в организме.
Птицеводство является одна из основных отраслей народного хозяйства, производящее для человека необходимые продукты питания — мясо, яйца и жир, а также перья в качестве набивочного материала. Многие виды птиц содержат в домашних условиях в качестве декоративных животных. Вследствие человеческой деятельности многие птицы получили своё дальнейшее развитие (а некоторые стали синантропами), но в то же время огромное их число оказалось на грани полного исчезновения — только с VII столетия вымерло 86 видов птиц[2]. В настоящее время около 1200 видов в той или иной степени подвержены риску вымирания и охраняются национальными и международными законодательствами.
Размеры
 Африканский страус — наиболее крупная современная птица
Африканский страус — наиболее крупная современная птицаСамым маленьким представителем этого класса считается обитающая на Кубе и острове Хувентуд колибри-пчёлка (Mellisuga helenae) — длина её самца не превышает 5,7 см, а самым крупным — африканский страус (Struthio camelus) высотой до 270 см и массой до 156 кг[9]. Таким образом, масса самой маленькой птицы в 66 000 раз меньше массы самой крупной из ныне живущих[10].
Андский кондор (Vultur gryphus) принадлежит к числу наиболее крупных летающих птиц — масса тела может достигать 11,4 кг, при размахе крыльев до 3,1 метров[11]. Длина взрослых птиц варьирует в пределах от 117 до 135 см[12][13].
Странствующий альбатрос (Diomedea exulans) достигает длины тела до 117 см и обладает самым большим среди птиц размахом крыльев, достигающим до 3,25 м[14][15].
Эволюция и систематика
Систематика
Первую попытку систематизации животных в IV веке до н. э. предпринял греческий учёный Аристотель — в своих сочинениях «О частях животных» и «О возникновении животных» он выделил всех известных ему птиц в «высший» род Ornithes[16][17]. Несмотря на очевидное несовершенство этой системы, вплоть до второй половины XVII века новых попыток классификации животного мира не предпринималось. В 1676 году вышла в свет рукопись Ornithologiae libri tres, написанная английским биологом Френсисом Виллоуби и уже после его смерти оформленная и опубликованная его другом и учёным Джоном Реем[18][19]. Это первая известная работа, в которой авторы попытались классифицировать птиц на основании внешних, морфологических признаков. Позднее, в 1758 году шведский натуралист Карл Линней активно использовал этот труд при создании своей Системы природы, в которой ввёл иерархические категории и биноминальную номенклатуру при обозначении видов, которые и используется вплоть до настоящего времени[20].
В системе Линнея все животные были разбиты на шесть основных категорий — классов, одну из которых, наряду с млекопитающими, амфибиями, рыбами, червями и насекомыми, заняли птицы, или Aves (позднее появились категории и более высокого порядка). Альтернативная филогенетическая систематика рассматривает группу Aves в качестве клады тероподных динозавров[21]. Согласно теории происхождения птиц от древних ящеров, Aves и её сестринская клада Crocodilia (крокодилы) считаются единственными современными ветвями клады рептилий Archosauria (архозавров). Все современные птицы имеют единого предка, которым некоторые учёные считают археоптерикса (Archaeopteryx lithographica) — животного, населявшего Землю в конце юрского периода 150—155 млн лет назад, который считается самой древней известной птицей планеты[22]. Другие учёные полагают, что археоптерикс не был предком современных (веерохвостых) птиц. Ряд известных динозаврологов и последователей системы ФилоКод, таких как Жак Готье, Луис Киаппи и их последователей, под категорией Aves понимают лишь современных птиц, не включая туда группы, известные лишь по ископаемым останкам — археоптерикса, энанциорниса, конфуциусорниса, патагоптерикса и некоторых других. Однако эти учёные ввели новую категорию Avialae, или птиц в широком смысле, которая объединяет живущие в наше время виды и их ископаемых предшественников[23].
Все современные группы птиц относятся к инфраклассу Neornithes, или веерохвостых птиц, который в свою очередь разделяется на две категории — Paleognathae, или бескилевые птицы (сюда входят, как правило, нелетающие птицы вроде страуса), и Neognathae, новонёбные птицы (включающий все остальные виды). Обычно эти две категории имеют ранг надотряда, хотя в работах Лайвези и Зуси рассматриваются как когорты[21]. В зависимости от системы классификации насчитывают от 9800[2] до 10 050[24] современных видов птиц.
Происхождение
 Археоптерикс — самый примитивный и ранний из всех известных представителей птиц[К 1]. Берлинский экземпляр.
Археоптерикс — самый примитивный и ранний из всех известных представителей птиц[К 1]. Берлинский экземпляр. Конфуциусорнис (Confuchisornis sanctus) — примитивная ископаемая птица[25] из раннего мелового периода Китая.
Конфуциусорнис (Confuchisornis sanctus) — примитивная ископаемая птица[25] из раннего мелового периода Китая.Наиболее распространена гипотеза, что птицы произошли от тероподных динозавров из группы манирапторов, куда помимо прочего входят дромеозавры и овирапторы[26]. По мере того, как учёные обнаруживают всё большее количество ископаемых останков нелетающих тероподов и тем не менее имеющих родство с пернатыми, точная граница между птицами и не-птицами становится размытой. Если раньше одним из определяющих признаков птиц было наличие перьевого покрова, то ряд открытий конца XX — начала XXI веков в провинции Ляонин на северо-востоке Китая показывает, что многие мелкие тероподы имели перья, внося свой вклад в эту неопределенность[27]. Однако группа учёных Университета штата Орегон США показала, что некоторые особенности дыхательной системы птиц не позволяют с определённостью утверждать, что их предками были тероподные динозавры[28], поскольку малоподвижность бедра птицы определяет в конечном итоге её способность летать, а бедренные кости динозавров подвижны. Кроме того, возраст некоторых обнаруженных останков птиц превосходит возраст останков динозавров, их теоретических предков.
Динозавры и птицы
В последние годы среди палеонтологов достигнут консенсус, что ближайшими родственниками птиц являются дейнонихозавры (Deinonychosauria) — инфраотряд, включающий в себя семейства дромеозавров (Dromaeosauridae) и троодонтид (Troodontidae). Вместе эти три категории были объединены в единую группу, получившую название Paraves. Центральное место в семействе дромаеозавров занимают микрорапторы (Microraptor gui) — четырёхкрылые мелкие хищники, которые были способны летать либо планировать. Тот факт, что дейнонихозавры в основном были очень маленького размера, дал основание предполагать, что предок птиц вёл древесный образ жизни и мог перемещаться планированием[29][30].
Теория эволюции птиц от рептилий возникла после открытия в 1860 году окаменелых останков археоптерикса — животного, жившего около 150 млн лет назад в верхней юре. Археоптерикс обладал характеристиками типичного пресмыкающегося — особым строением таза и рёбер, зубами, когтистыми лапами и длинным, как у ящериц, хвостом. В то же время найденные окаменелости продемонстрировали хорошо сохранившиеся отпечатки маховых крыльев с перьями, аналогичных тем, что имеются у птиц. В отличие от предыдущих исследований, это существо уже не рассматривают в качестве общего прародителя всех современных птиц, однако оно считается древнейшим и наиболее примитивным представителем класса Aves или Avialae, и вероятно имеет близкое родство с настоящим предком[31]. Среди возможных ближайших родственников птиц рассматривается также авимим — маленький хищный динозавр с большими глазами и крупным мозгом, населявший Землю в меловом периоде около 75 млн лет назад[32].
Альтернативные версии происхождения
За обозримый период времени было выдвинуто несколько научных версий происхождения и родственных связей птиц, причём в течение более чем 100 лет они были чисто гипотетическими и основывались лишь на одном единственном экземпляре археоптерикса, найденом в Германии в середине XIX века[33]. Наряду с теорией динозавров существует гипотеза, что птицы могли эволюциионировать из мелких и более примитивных псевдозухий (Pseudosuchia) из группы текодонтов в поздней перми или раннем триасе. Среди последователей «динозавровой» версии также имелись расхождения, какая группа могла быть наиболее вероятным предком всех позднейших птиц — птицетазовые или ящеротазовые тероподы[34]. Несмотря на то, что птицетазовых ящеров и птиц объединяет схожая форма лобковых костей таза, полагают что последние всё-таки произошли от ящеротазовых и развили аналогичное строение самостоятельно[35]. На самом деле, похожая на птичью структура таза также развивалась у третьей части специфической группы тероподов, известных как теризинозавры.
Ряд учёных, среди которых такие известные специалисты как Ларри Мартин и Алан Федучия, продолжают настаивать, что птицы произошли не от динозавров, а от более ранних архозавров типа лонгисквам (Longisquama). В большинстве своих публикаций эти учёные приводят доводы в пользу того, что сходство между птицами и манираптором есть результат конвергентной эволюции, и они никак между собой не связаны. В конце 1990-х годов, когда эта наследственная связь стала почти очевидной, Федучия и Мартин выдвинули модифицированную версию гипотезы, согласно которой манирапторы являлись нелетающими потомками ранних птиц[36], а сами птицы всё-таки произошли от лонгисквам. Таким образом, в этой версии ни одна группа современных птиц по сути динозаврами не является, а манираптор есть нелетающая птица-архозавр[37]. Эта теория оспаривается большинством палеоорнитологов[38]. Те особенности, которые автор приводил в качестве доказательства нелетаемости животного, было интерпретировано как экзаптация, или «пре-адаптация», которую манирапторы наследовали от их общего с птицами предка.
Наконец, ещё одну версию происхождения птиц отстаивает известный российский палеонтолог Е. Н. Курочкин. По его мнению, предком настоящих пернатых является протоавис (Protoavis texensis), существо на 75 млн лет более древнее, чем археоптерикс[33][39]. Останки этого животного были обнаружены в 1991 году в Техасе, США. По мнению Курочкина, протоавис имел ряд прогрессивных особенностей, гораздо более близких к современным птицам, чем у археоптерикса. Критики этого предположения утверждают, что останки плохо сохранились, подверглись тщательной реконструкции и вообще могут являться «химерой», то есть принадлежать разным видам животных. Большинство специалистов склоняются к версии, что череп находки принадлежит очень ранней форме целурозавров[40].
Ранняя эволюция птиц
Aves Pygostylia Confuciusornithidae
Ornithothoraces Enantiornithes
Ornithurae Neornithes
Филогения по Chiappe, 2007[41] С точки зрения современной науки, ранняя эволюция птиц выглядит очень запутанной и сложной. Диверсификация, или эволюционное разделение на множество разнообразных форм, началось в меловом периоде[41]. Многие из этих форм, в том числе и предки современных веерохвостых птиц, на первоначальном этапе сохраняли доставшиеся им от динозавров крылья с когтями и зубы, которые впоследствии были редуцированы. Археоптерикс, джехолорнис (Jeholornis) и другие древнейшие птицы обладали длинным хвостовым отделом позвоночника[41]. У других более развитых групп, таких как представителей клады пигостилевых (Pygostylia), 4—6 задних хвостовых позвонков были слиты воедино, образовав кость, известную как пигостиль.
Первой широко распространённой и разнообразной группой короткохвостых птиц стали энанциорнисы (Enantiornithes), или «противоположные птицы», названные так потому, что строение их плечевых костей было противоположно строению таковых у современных птиц. Внешне похожие на современных птиц и развивавшиеся вместе с ними в меловом периоде, энанциорнисы заняли всевозможные экологические ниши от обитателей песчаных морских побережий и охотников за рыбой до ведущих древесный образ жизни и питающихся семенами[41]. Более развитые ихтиорнисы (Ichthyornithes), слегка напоминающие чаек, обитали на морских просторах и также питались рыбой[42]. Морские птицы из отряда гесперорнисообразных из мезозойской эры настолько приспособились к охоте за рыбой, что в процессе эволюции утратили способность к полёту и полностью перешли к водному образу жизни. Несмотря на такую узкую специализацию, гесперорнисообразные представляют собой одних из ближайших родственников современных птиц[41].
Эволюция современных птиц
Обнаруженные на архипелаге Вега в Западной Антарктиде останки древней птицы, напоминающей крупную утку и названной впоследствии вегавис (Vegavis iaai), показали, что по крайне мере одна эволюционных линия веерохвостых птиц получила развитие в начале мелового периода[43] и впоследствии разделилась на две основные формы, известные сегодня как подклассы бескилевых и новонёбных птиц. Бескилевые представляют относительно небольшую группу, куда относят страусообразных и обитающих в Южной и Центральной Америке скрытохвостых. Следующим расхождением от остальных новонёбных птиц стали представители надкласса Galloanserae — категории, куда включают гусеобразных (лебеди, гуси, утки и паламедеи) и курообразных (тетерева, фазаны, большеноги, краксы и им подобные). Период, когда началось это расхождение, является предметом научных споров — большинство специалистов сходятся во мнении, что веерохвостые получили своё развитие в меловое время, а ветвь Galloanserae отделилась от остальных новонёбных до начала биологической катастрофы третичного периода, погубившей динозавров. Тем не менее, у этой версии имеются и противники, полагающие что радиация могла произойти уже после (и благодаря) гибели большей части животного мира[44]. Разногласие отчасти вызвано различными методами исследований: молекулярная генетика указывает на основную диверсификацию в середине мелового периода, в то время как анализ окаменелостей говорит о начале палеогена. Попытки согласовать результаты этих двух методик ставят специалистов в тупик[44][45].
Классификация птиц до сих пор находится в стадии формирования. Одна из ключевых работ в этом направлении — молекулярно-эволюционное исследование «Филогения и классификация птиц» (Phylogeny and Classification of Birds), проведённое американцами Чарльзом Сибли и Джоном Алквистом и опубликованое в 1990 году[46]. Результаты этого труда часто дискутируются в научных кругах либо подвергаются ревизии. Большинство авторов соглашается, что расположение отрядов указано точно[47], однако не соглашается с родственными отношениями между отрядами; сравнительный анализ анатомии птиц, ископаемых окаменелостей и ДНК не привёл к согласию по этому вопросу. Множественные находки последнего времени и молекулярные исследования рисуют всё более и более ясную картину эволюции современных птиц.
Распространение и места обитания
Большая общая подвижность и высокий уровень метаболизма и гомойотермия, развитие органов чувств, сложное поведение, способность приспосабливаться к меняющимся условиям среды, а также широкий спектр питания позволили птицам широко расселиться по всему земному шару. На Земле нет ни одного района, где бы ни встречались те либо иные виды птиц. Каждому крупному географическому району свойствен свой состав фауны, включающей как эндемичные виды, так и виды, населяющие соседние области. Птицы населяют практически все наземные биомы включая и внутренние районы Антарктиды — например, снежный буревестник (Pagodroma nivea) гнездится в глубине этого материка на расстоянии до 440 км от берега[48]. По краям Антарктического щита местами гнездятся пингвины (императорский, Адели), снежный и гигантский буревестники, южный поморник. Населяют птицы и самые безводные пустыни и горы вплоть до границы вечных снегов: на высотах до 5500 — 6500 метров гнездятся бородач, грифы, некоторые виды колибри, клушица, гималайская завирушка и др., а во время миграций стаи пролетающих гусей, журавлей и иногда наблюдали на высоте 7000 — 9000 м н.у.м. Заселяют птицы и наиболее удаленные от материков океанические острова. На вновь появившихся вулканических островах, некоторые морские птицы поселяются ещё до появления там растений и других животных. Несколько семейств птиц приспособились к жизни на море, и некоторые виды морских птиц достигают берега лишь с целью размножения[49]. Птицы обычно не ныряют глубже 50—60 м, но отдельные виды пингвинов ныряют на глубину до 300 м.[50]. Наибольшего биоразнообразия птицы достигают в тропических широтах: здесь встречается около 85 % ныне живущих видов и подвидов, тогда как в умеренных и холодных широтах — лишь 15 %. Многие семейства, характерные для тропических широт, либо совсем не проникают в умеренные широты, либо представлены там немногими видами.
Только единичные виды птиц характеризуются очень широким распространением, встречаясь в подходящих биотопах на всех материках (кроме Антарктиды) и на многих островах. Это сокол-сапсан и скопа. Несколько меньше ареал сипухи, морского зуйка, деревенской ласточки и некоторых других видов. Значительно больше видов, ареалы которых простираются на два-три континента (Евразия и Северная Америка, Евразия и Северная Африка и т. п.). Преобладающее большинство видов имеет гнездовые ареалы, охватывающие лишь часть континента, но перелётные виды с такими ареалами на зимовках часто встречаются на других континентах. В пределах своего ареала каждый вид птиц распространен неравномерно (говорят о «кружеве ареала»). Существуют виды с небольшими ареалами. Некоторые виды колибри свойственны лишь отдельным горам в Андах. Только на Ямале и Таймыре гнездится краснозобая казарка. Другой эндемик нашей — кулик-лопатень, который гнездится только в приморской полосе Чукотского полуострова. Ареал одной из американских славок Dendroica kirtlandi занимает площадь 140 на 160 км в Северном Мичигане.
Вследствие интродукции человеком многие птицы успешно обосновались в дикой природе регионов, где они никогда ранее не обитали. Во многих случаях такая интродукция была сознательной: например, обыкновенный скворец попал на американский континент в конце XIX века благодаря поклонникам Уильяма Шекспира, пожелавшим расселить в Центральном парке в Нью-Йорке всех птиц, воспетых этим драматургом[51]. В других случаях попадание в чужеродную среду оказалось случайным: так, содержащийся в домашних условиях попугай-монах из клетки попал на волю и успешно обосновался в нескольких городах Северной Америки[52]. Наконец, ареал некоторых видов, таких как египетской цапли[53], химахимы (Milvago chimachima)[54] и розового какаду[55] расширился далеко за пределы первоначальных территорий обитания, поскольку сельскохозяйственное использование земель создало новую подходящую среду обитания.
Особенности строения
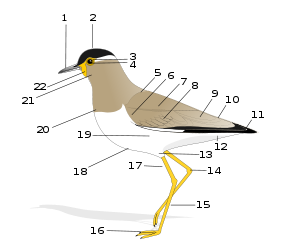 Общее строение птицы:
Общее строение птицы:
1. Клюв 2. Голова 3. Радужная оболочка 4. Зрачок 5. Спина 6. Малые кроющие крыла 7. Плечо 8. Кроющие второстепенных маховых 9. Кроющие первостепенных маховых 10. Надхвостье 11. Первостепенные маховые 12. Подхвостье 13. Бедро 14. Предплюсневой сустав 15. Плюсна 16. Пальцы 17. Голень 18. Брюхо 19. Бок 20. Грудь 21. Горло 22. СерёжкаВнешняя морфология вид птиц отражает их приспособленность к полёту. Существует относительно небольшое количество (около 60 видов) нелетающих либо почти нелетающих птиц, однако все они в процессе эволюции так или иначе утратили это качество, которое имели их предки[56]. Способность к полёту определяет особенности этого класса животных, включая общий план строения[57].
Туловище птиц обтекаемой яйцеобразной формы, отличающееся компактностью[57]. Шея у большинства видов тонкая и гибкая. На голове вперед выдается клюв, образованный надклювьем и подклювьем. Для полета служат видоизмененные передние конечности — крылья. Их несущую плоскость образуют большие маховые перья. Ноги птиц принимают всю тяжесть тела, при передвижении по земле, взлёте и посадке, лазании по деревьям. Ноги птиц имеют четыре отдела: бедро, голень, цевку и пальцы. Обычно ноги четырёхпалые, но иногда число пальцев сокращается до трех и даже двух (африканский страус). Из четырёх пальцев в преобладающем большинстве случаев три являются направленными вперед, а одни — назад[57]. Передвижение по земле производится обычно при помощи ног. При этом бедренные кости малоподвижны, поэтому при перемещении по земле они практически не смещаются из горизонтального положения. Именно такая фиксированная позиция кости позволяет поддерживать брюшной воздухоносный мешок на вдохе, что определяет работу дыхательной системы птиц и позволяет им летать[28]. Это одна из основных характеристик птиц; все прочие животные, передвигающиеся по земле, имеют подвижный тазобедренный сустав.
Движение в воздухе осуществляется при помощи передних конечностей, преобразованных в крылья, и хвоста. Крылья служат как аэродинамическими поверхностями, удерживающими птицу в воздухе, так и источниками тяги для движения вперёд[58].
Другой особенностью, отличающей птиц от других хорошо летающих позвоночных, являются хорошо развитые массивные мышцы ног, особенно заметные при сравнении птиц с рукокрылыми и птерозаврами, с их почти редуцированными ногами[59].
Оперение и роговой покров
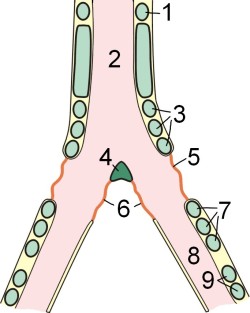
См. также: Оперение (биология) и Перо Перо, 1 — Опахало. 2 — Ствол. 3 — Обычная поверхность. 4 — Пуховая часть. 5 — Очин (стержень).
Перо, 1 — Опахало. 2 — Ствол. 3 — Обычная поверхность. 4 — Пуховая часть. 5 — Очин (стержень). Типы перьев у птиц: 1 — контурное, типичное маховое перо, 2 — рулевое перо (перо хвоста), 3 — покровное перо, 4 — нитевидное перо, 5 — кистеобразное перо, 6 — пуховое перо.
Типы перьев у птиц: 1 — контурное, типичное маховое перо, 2 — рулевое перо (перо хвоста), 3 — покровное перо, 4 — нитевидное перо, 5 — кистеобразное перо, 6 — пуховое перо. Перья крыла птицы: 1 — маховые перья 1-го порядка; 2 — большие кроющие перья; 3 — крылышко; 4, 8 — маховые перья 2-го порядка; 5 — оперение плеча; 6 — средние кроющие перья; 7 — кроющие перья крыла.
Перья крыла птицы: 1 — маховые перья 1-го порядка; 2 — большие кроющие перья; 3 — крылышко; 4, 8 — маховые перья 2-го порядка; 5 — оперение плеча; 6 — средние кроющие перья; 7 — кроющие перья крыла. Структура пера сине-жёлтого ара
Структура пера сине-жёлтого араКожные покровы птиц тонкие, эластичные. В соединительнотканном слое имеются обильные пучки гладких мышц, прикрепляющиеся к очинам контурных перьев и изменяющие их положение. Кожные железы отсутствуют, единственной кожной железой у птиц является копчиковая железа, которая находится над хвостовыми позвонками (отсутствует у бескилевых птиц, у некоторых дроф, голубей, попугаев и др.). Она секретирует маслянистый секрет, который птицы выдавливают клювом и смазывают им оперение, что способствует сохранению эластичности пера[60].
Для всех видов птиц характерным является наличие перьевого покрова, который не встречается у других современных животных[57]. Перья покрывают все тело птицы, за исключением клюва и дистальных частей задних конечностей. У некоторых птиц, у как например индеек и американских грифов, оперение на голове и шее либо отсутствует вовсе, либо выражено очень слабо[60].
Начальные стадии эмбрионального развития пера сходны с развитием чешуи. Поэтому можно говорить, что перья возникли в результате эволюционных преобразований чешуи. Эволюционное происхождение перьев можно проследить начиная с хищных динозавров синозавроптерикса и дилонга, покрытых волокнистым пухом[61] У каудиптерикса, синорнитозавра и микрораптора можно наблюдать настоящие перья[61].
Контурные перья, покрывают все тело птицы и имеют хорошо развитый плотный стержень, основание которого — полый очин — охватывается находящейся в коже перьевой сумкой. Глубина перьевой сумки больше у крупных перьев. От стержня отходят упругие бородки, которые несут бородочки с крючочками, которые сцепляются с крючочками соседних бородок, образуя опахало пера. В самой нижней части пера бородки обычно являются более мягкими и длинными, а их бородочки не имеют крючочков — этот участок называют пуховой частью опахала. Особенности строения перьев могут варьировать у разных групп птиц. Так виды, живущих в суровых температурных условиях, обычно обладают более сильно развитой пуховой частью опахала[60]. У большинства птиц имеются пуховые перья (стержень мягкий) и пух (стержень совсем редуцирован), мягкие и длинные бородки которых несут мягкие бородочки, лишенные крючочков, из-за чего сцепленного опахала не образуется. Между типичным пером, пуховым пером и пухом существуют разнообразные промежуточные типы. Пуховые перья обычно располагаются по птерилиям. Пух относительно равномерно покрывает всё тело (веслоногие,гусеобразные, многие хищные птицы и др.), либо имеется только на аптериях (цапли, куриные, совы, многие воробьиные и др.), либо — только на птерилиях (тинаму). Обычно пуховые перья и пух прикрыты контурными перьями. Только у немногих птиц (грифы, марабу и др.) голова и часть шеи покрыты только пухом. Нитевидные перья расположены под контурными перьями, обладают длинным тонким стержнем и редуцированными бородками. Видимо, они выполняют осязательную функцию[60].
Зимнее перо характеризуется тем, что его пуховая часть опахала и добавочный стержень обычно развиты значительно лучше, чем в летнем пере. У козодоев и особенно у сов хорошо выряжена бархатистость (ворсистость) всех перьев, в том числе маховых и рулевых; гася завихрение воздушных потоков, ворсистость обеспечивает бесшумность полета. У водоплавающих птиц относительно короткие и сильно изогнутые перья плотно прилегают друг к другу, предотвращая намокание оперения; при этом водостойкость оперения обеспечивается и особой формой бородок и бородочек, способствующей возникновению на поверхности пера водной пленки[60].
Общее число перьев у крупных видов больше, чем у мелких. Напрмиер, у колибри около 1000 перьев, у мелких воробьиных 1,5 — 2 500, у чаек 5 — 6 000, у уток 10—12 000, у лебедей 25 000 и т. п. Общее число перьев, их размеры и особенности строения в варьируют и у близких групп в зависимости от их экологической специализации[60].
Оперение делает форму тела обтекаемой и увеличивают площадь крыльев и хвоста, что в немалой степени способствует лётным качествам, а также обеспечивает терморегуляцию. Кроме того, с помощью перьев птицы передают друг другу сигналы, маскируются от хищников[62].
Перья представляют собой роговые накожные образования, растущие из расположенных рядами углублений кожи, называемых птерилиями. Лишь у немногих не летающих птиц, как например, у пингвинов, птерилии не выражены, а перья растут равномерно по всему телу[63]. Расположение и форма птерилий часто служат систематическим признаком. В пределах одного вида окрас и форма перьев может различаться в зависимости от возраста, пола[64] или социального статуса птицы[65]. Во время формирования перьев в ороговевающих клетках откладываются пигменты, обусловливающие характерную для каждого вида окраску. Наиболее распространеныными являются пигменты двух типов: меланины и липохромы. Меланины обусловливают окраску перьев в разные оттенки чёрного, коричневого, красновато-бурого и жёлтого цветов. Липохромы обеспечивают более яркую окраску: красную, зелёную, жёлтую, синюю и др. Сочетание в одном пере разных пигментов усложняет окраску. Белая окраска создается полным отражением света от наполненных воздухом прозрачных полых роговых клеток пера при отсутствии пигментов[60].
Функции оперения весьма многообразны. Оно обеспечивает возможность полёта, образуя несущие плоскости (крылья, хвост) и создавать обтекаемость тела. Перья защищают кожные покровы от механических повреждений. Очень эффективна водозащитная и теплозащитная функции оперения: черепицеобразно прилегающие друг к друга вершины контурных перьев противостоят намоканию, а переплетение пуховых частей опахал контурных перьев, пуховых перьев и пуха удерживает около кожных покровов неподвижный слой воздуха, что уменьшает потери тепла[60].
Периодически птицы линяют: старые перья выпадают, а на их месте вырастают новые. Обычно линька происходит один раз в году, реже — два и совсем редко, как например у морянки (Clangula hyemalis) — три раза в год. Крупные хищные птицы способны линять один раз в несколько лет. В случае повторной линьки различают гнездовой и зимний наряды, а также пуховой наряд для неоперившихся птенцов. Как правило, смена маховых и рулевых, необходимых для полёта перьев происходит в определённой последовательности, так что птицы и в процессе линьки сохраняют свои лётные качества. Из этого правила есть и исключения — например, у утиных все маховые опадают одновременно, в результате чего те теряют способность летать[66]. Перед насиживанием у самок большинства видов на брюхе образуется так называемое наседное пятно — неоперённый участок кожи с развитыми кровеносными сосудами, которым птица прижимается к яйцам и согревает их[67].
Перья требуют тщательного ухода, и птицы ежедневно тратят около 9 % времени на чистку своего оперения, купание и пылевые ванны[68]. Быстролетающие птицы — стрижи, ласточки и крачки на лету окунаются в воду. Другие встряхиваются на мелководье либо смачивают перья в росе или дождевой воде[63].
Ухаживают за перьями птицы с помощью клюва. Лишь одна птица не может этого делать - колибри-мечеклюв обладает настолько длинным клювом, что ей приходится чистить свои перья не клювом, а лапкой[69]. С помощью клюва из перьев удаляются чужеродные предметы, после чего тело покрывается жиром, выделяемым из копчиковой железы у основания хвоста. Чтобы нанести жир на голову, птицы сначала наносят его на ноги, а затем лапами растирают голову. Выделения придают оперению эластичность и действуют как противомикробное средство, препятствуя размножению разрушающих перья бактерий[70]. Наконец, многие виды специально копаются в муравейниках с целью нанесения на тело муравьиной кислоты — такой процесс, известный как «муравление», также избавляет птиц от паразитов.
Маховые и рулевые перья при линьке выпадают попарно, благодаря чему правильность движений не нарушается. У большинства птиц птенцовые маховые перья не меняются в течение года, и полной линьки не бывает столько же времени. Но у куриных, начинающих летать очень рано, птенцовые маховые быстро изнашиваются и рано заменяются новыми. Утиные птицы (утки, гуси, лебеди), утрачивают сразу маховые и потому долгое время не могут летать.
Скелет и мускулатура
 Скелет птицы на примере голубя. 1 — череп, 2 — шейные позвонки, 3 — вилочка 4 — клювовидный отросток, 5 — крючковидные отростки ребер, 6 — киль, 7 — коленная чашечка, 8 — цевка, 9 — пальцы, 10 — большая берцовая кость (тибиотарсус), 11 — малая берцовая кость (тибиотарсус) 12 — бедро, 13 — седалищная кость, 14 — лобковая кость, 15 — подвздошная кость, 16 — хвостовые позвонки 17 — пигостиль, 18 — сложный крестец, 19 — лопатка, 20 — поясничные позвонки, 21 — плечевая кость, 22 — локтевая кость, 23 — лучевая кость, 24 — запястья, 25 — кисть, 26 — пальцы, 27 — крылышко.
Скелет птицы на примере голубя. 1 — череп, 2 — шейные позвонки, 3 — вилочка 4 — клювовидный отросток, 5 — крючковидные отростки ребер, 6 — киль, 7 — коленная чашечка, 8 — цевка, 9 — пальцы, 10 — большая берцовая кость (тибиотарсус), 11 — малая берцовая кость (тибиотарсус) 12 — бедро, 13 — седалищная кость, 14 — лобковая кость, 15 — подвздошная кость, 16 — хвостовые позвонки 17 — пигостиль, 18 — сложный крестец, 19 — лопатка, 20 — поясничные позвонки, 21 — плечевая кость, 22 — локтевая кость, 23 — лучевая кость, 24 — запястья, 25 — кисть, 26 — пальцы, 27 — крылышко. Разнообразие форм клюва, которые обеспечивают у одних схватывание добычи, иногда её расчленение, у других — долбление, рытьё и т. д., а также различные сложные манипуляции.
Разнообразие форм клюва, которые обеспечивают у одних схватывание добычи, иногда её расчленение, у других — долбление, рытьё и т. д., а также различные сложные манипуляции.В строении скелета птиц отчетливо видны адаптивные особенности, связанные с полётом как основным средством передвижения, в некоторых случаях у специализированных видов — с модифицированными приспособлениями к нырянию, плаванию, горизонтальному бегу или лазанию по стволам или ветвям деревьев, скалам и другим вертикальным поверхностям[10][57].
Скелет предельно упрощён и состоит из лёгких и прочных костей. Некоторые кости имеют наполняемые воздухом полости, называемые «пневматическими», связанные с органами дыхания[71].
Кости черепа слиты воедино и не имеют черепных швов[62]. Глазницы большие и разделены между собой костной перегородкой. Череп соединяется с позвоночником при помощи одного затылочного мыщелка, нижняя челюсть прикрепляется к черепу посредством квадратной кости, играющей роль подвеска.
Очень важная особенность черепа птиц — подвижность надклювья, усиливает силу укуса и увеличивает размеры зева, а подвижное нёбо помогает проталкиванию пищевого комка в пищевод. Основание надклювья у многих птиц покрыто восковицей. Форма и длина клюва, особенности его рогового чехла, характер подвижности, размеры ротового отверстия у различных птиц варьируют в широких пределах, отражая пищевую специализацию видов[60].
Позвоночный столб делится на шейный, грудной, поясничный и хвостовой отделы. Для всех отделов позвоночника, кроме шейного, характерно сращение многих позвонков между собой. Эта особенность позвоночного столба обусловливает неподвижность туловища птиц, что имеет большое значение при полете. Для птиц характерна длинная подвижная шея. Её образуют 11—25 шейных позвонков. Шейный отдел очень длинный и отличается большой гибкостью, однако подвижность значительно уменьшается в грудном и полностью отсутствует в поясничном отделе[72]. В шейном отделе может быть различное число позвонков (9—25). Кроме атланта и эпистрофея все остальные шейные позвонки у птиц гетероцельного типа и имеют седлообразные суставные поверхности. Это обеспечивает относительную подвижность в горизонтальной и вертикальной плоскостях, но не вокруг оси. Вращение головы вокруг своей оси обеспечивается особым строением первого (атлант) и второго (эпистрофей) шейных позвонков. В грудном отделе 3—10 позвонков, которые у большинства птиц срастаются, образуя спинную кость. Все поясничные, крестцовые и часть хвостовых позвонков вместе с тазом объединены в единую кость, называемую сложным крестцом[62]. Число свободных хвостовых позвонков составляет от 5 до 9. Последние хвостовые позвонки образуют копчиковую кость (пигостиль). Ребра состоят из двух частей — позвоночной и грудинной. У птиц (кроме паламедей) рёбра снабжены крючковидными отростками, к которым прикрепляются межрёберные мышцы. Грудина у летающих птиц и у пингвинов, имеет киль, к которому прикрепляется мощная сильно развитая летательная мускулатура (подключичные и большие грудные мышцы); У бегающих птиц (страусов, казуаров и др.) киля нет. Плечевой пояс состоит из мощных удлинённых коракоидов, сросшихся ключиц (вилочки) и длинных узких лопаток, которые лежат над рёбрами. Кости пясти и некоторые кости запястья срастаются в единое образование — пряжку. Свободными остаются только две проксимальные кости запястья.
У птиц передние конечности превратились в крылья, которые состоят из хорошо развитой плечевой кости, прямой и более тонкой лучевой кости и изогнутой более толстой локтевой кости и кисти, в которой произошла редукция ряда костей и слияние некоторых из них между собой. Пястный отдел кисти представляет собой две кости, соединенные своими концами, образующие «пряжку». Сохранились рудименты трех первых пальцев, наиболее развитым из которых является второй. В результате этих изменений передняя конечность потеряла способность к передвижению по земле.
Таз птиц открытый: лобковые и седалищные кости своими концами не соединяются внизу с соответствующими костями противоположной стороны (лобковое сочленение имеется только у страусов), что позволяет откладывать крупные яйца[60]. Большая поверхность тазовых костей и их прочное соединение с осевым скелетом создают опору для задних конечностей, обеспечивая место прикрепления мощной мускулатуры. Округлая головка относительно короткого бедра, входит в вертлужную впадину тазового пояса почти под прямым углом к его оси, что обеспечивает относительно вертикальное положение задней конечности. Коленный сустав спереди прикрывает коленная чашечка, особенно хорошо развитая у ныряющих птиц[60]
Большая берцовая кость срастается с проксимальными костями предплюсны (пяточной и таранной) в одну кость — беговую, или большеберцово-заплюсневую (тибиотарзус). Малая берцовая кость редуцирована. Дистальный ряд костей предплюсны и все кости плюсны срастаются в единое костное образование — цевку, что дает дополнительный сустав на ногах. опора пальцев, одновременно увеличивающая длину шага. Особенно удлинена цевка у околоводных птиц (куликов, голенастых, журавлей)[60].
Пальцев чаще всего четыре, фаланговая формула обычно 2-3-4-5. У подавляющего числа птиц развито четыре пальца: первый направлен назад, три остальных — вперёд. Пальцы обычно очень подвижны и способны совершать разнообразные движения: хватание, обхват веток, цепляние и т. п[60]. У совообразных, бананоедов, скопы и некоторых других второй палец может произвольно быть направлен либо вперед, либо назад. У ряда древесных птиц (попугаи, кукушки, дятлы) два пальца являются направленными вперед, два (первый и четвёртый) — назад. Длина пальцев, развитие когтей определяются экологической специализацией. У околоводных птиц длинные пальцы позволяют двигаться по вязкому грунту; у наземных бегающих птиц пальцы укорачиваются и утолщаются, а задний палец либо исчезает, либо сохраняется в сильно уменьшенном виде[60]. На подошвенной части глубокого сгибателя пальцев имеются поперечные выступы. Благодаря этому сухожилие прочно фиксируется; когда птица спит, пальцы крепко охватывают ветку даже при расслаблении мышц.
Последняя фаланга каждого пальца прикрыта хорошо развитым когтем. Когти служат опорой при ходьбе, помогают птице при лазанье, чистке оперения, постройке гнезда. Ряд видов когтями удерживает или умерщвляет добычу. Слабо развитые рудиментарные коготки имеются у некоторых птиц на первом (некоторые хищные птицы, гуси, паламедеи) и на втором пальцах крыла (гоацин, казуары, киви). Крупные костные выросты, покрытые роговым чехлом — шпоры — развиваются на цевке у многих фазановых[60].
Мускулатура птиц характеризуется развитием грудных и подключичных мышца, приводящих в движение крылья. Мощно развиты также мышцы ног, выполняющие большую работу при хождении птицы и передвижении по земле, веткам деревьев, во время взлета и посадки.
Пищеварительная и выделительная система

Пищеварительная система птиц начинается ротовой полостью. Зубы у современных птиц отсутствуют[57] — их отчасти заменяют острые края клюва, которым птицы захватывают, удерживают и порой размельчает пишу. В ротовой полости располагаются слюнные железы. У некоторых птиц липкая слюна обеспечивает приклеивание мелкой добычи к языку (дятлы и др.). Слюнные железы слабо выражены или отсутствуют совсем у видов, питающихся достаточно влажным кормом. Длинный пищевод у некоторых птиц (куриные, дневные хищники, голуби, попугаи и др.) имеет чётко выраженное расширение — зоб, служащий для временного хранения пищи. В нём корм, подвергшийся обработке слюной, набухает и размягчается. Своеобразие птиц — разделение желудка на два отдела: железистый (где выделяются пищеварительные ферменты) и мускульный. Из пищевода пища попадает в железистый отдел желудка, а из него переходит в мускулистый отдел. Его стенки образованы мощными мышцами, а в полости, обычно, находятся гастролиты. Они и складки желудка при сокращении мышц стенок перетирают и размельчают содержимое, компенсируя отсутствие зубов[57][73]. Не поддающиеся перетиранию компоненты (шерсть, перья, хитин, осколки костей и т. п.) у многих птиц (совы и дневные хищники, чайки, некоторые воробьиные и др.) спрессовываются в мускульном желудке в погадку и через пищевод и ротовую полость выбрасываются наружу[60].
Кишечник у птиц относительно короткий: у разных видов он длиннее туловища в 3 — 12 раз, редко больше. Относительно длинный он у видов, питающихся грубой растительной пищей. В нём выделяют более длинный тонкий отдел и менее протяженный толстый отдел. Обычно парные слепые кишки у большинства птиц малы и выполняют роль лимфоидных органов. Однако у некоторых растительноядных видов (страусы, куриные, многие гусеобразные и др.) слепые кишки характеризуются большими размерами, и в них идет активное переваривание пищи[60]. Прямая кишка у птиц является не развитой, фекалии не накапливаются в кишечнике, что облегчает массу птицы. Кишечник заканчивается расширением — клоакой, в которую также открываются мочеточники и протоки половых желез[57].
Для компенсации энергозатрат во время полёта требуется быстрый обмен веществ, и по этой причине у большинства птиц пищеварительная система адаптирована к быстрому процессу переваривания[69]. Процессы пищеварения у птиц протекают очень быстро: у свиристели ягоды рябины проходят весь кишечник за 8—10 мин.[57], а у утки через 30 мин. после проглатывания карася длиной 6 см, его остатков уже нельзя обнаружить в кишечнике[57]. Энергетические резервы накапливаются в виде гликогена в печени и особенно в виде жира — подкожного и внутреннего. В период миграций запасы жира у некоторых видов может достигать 30 — 50 % от общей массы тела[60].
Также как и пресмыкающиеся, птицы, как правило, урикотеличны, то есть их почки извлекают азотсодержащие отходы из кровотока и выделяют их в виде мочевой кислоты, а не мочевины или аммиака, как у млекопитающих. Крупные метанефрические почки лежат в углублениях тазового пояса. Мочеточники открываются в клоаку, мочевой пузырь отсутствует[74][75]. Некоторые птицы, как например колибри, составляют исключение — их азотистые отходы могут выделяться в виде аммиака, то есть птицы по сути являются аммонотеличными[76]. Ещё одним конечным продуктом обмена веществ является креатин (у млекопитающих эту функцию выполняет креатинин)[62].
Все эти вещества скапливаются в кишечнике и затем выделяются из клоаки птицы[77][78].
У морских птиц (трубконосые, чайки, чистики, пеликанообразные и др.) и некоторых пустынных птиц (африканский страус, пустынная курочка и др.) добавочными органами солевого обмена являются надорбитальные железы, способные выделять избыток солей в виде капелек секрета, стекающего из ноздрей[60].
Дыхательная система
 Эритроциты птиц, в отличие от круглых безъядерных эритроцитов млекопитающих, имеют элиптическую форму и содержат ядра[79].
Эритроциты птиц, в отличие от круглых безъядерных эритроцитов млекопитающих, имеют элиптическую форму и содержат ядра[79].Дыхательная система птиц также характеризуется признаками приспособления к полёту, во время которого организм нуждается в усиленном газообмене. Эта система органов у птиц считается одной из самых сложных среди всех групп животных[62]. От глотки отходит длинная трахея, делящаяся в грудной полости на два бронха. На месте бифуркации трахеи имеется расширение — нижняя гортань, в которой расположены голосовые связки; стенки её имеют костные кольца. Нижняя гортань представляет собой голосовой аппарат и наиболее сильно развита у птиц, поющих и издающих громкие звуки. Легкие птиц являются малыми по объёму, малоэластичными и прирастают к ребрам и позвоночному столбу[60]. Они характеризуются трубчатым строением и очень густой капиллярной сетью. С легкими связано 5 пар воздушных мешков — тонкостенных, легко растяжимых выростов вентральных ответвлений крупных бронхов, находящиеся среди внутренних органов, между мышцами и в полостях трубчатых костей крыльев. Эти мешки играют большую роль в процессе дыхания птиц во время полета[57]. Наряду с функцией дыхания воздушные мешки несут добавочные функции: облегают массу тела птицы, и примыкая к крупным группам мышц участвуют в терморегуляции (рассеивание лишнего тепла)[60].
Лёгкие устроены таким образом, что воздух проходит через них насквозь. При вдохе только 25 % наружного воздуха остаётся непосредственно в лёгких, а 75 % проходит через них и попадает в специальные воздушные мешки. При выдохе воздух из воздушных мешков опять идёт через лёгкие, но уже наружу, образуя так называемое двойное дыхание. Таким образом, лёгкие постоянно насыщаются кислородом как во время вдоха, так и выдоха[80]. В состоянии покоя дыхание птицы осуществляется путем расширения и сжатия грудной клетки. При полёте, когда движущимся крыльям нужна твердая опора, грудная клетка птиц остается практически неподвижной и прохождение воздуха сквозь лёгкие обусловливается путём расширения и сжатия воздушных мешков[57]. Чем быстрее машущий полет, тем интенсивнее дыхание. При подъеме крыльев они растягиваются и воздух самостоятельно засасывается в легкие и в воздушные мешки. При опускании крыльев происходит выдох и через легкие проходит воздух из мешков[57].
Голосовой аппарат
 Схема строения сиринкса — «нижней гортани»
Схема строения сиринкса — «нижней гортани»
1 — последнее свободное хрящевое кольцо трахеи; 2 — тимпанальный орган; 3 — первая группа колец сиринкса; 4 — козелок; 5' — боковая тимпанальная мембрана; 6 — средняя тимпанальная мембрана; 7 — вторая группа колец сиринкса; 8 — бронх; 9 — хрящевые кольца бронхаГолосовой аппарат птиц имеет не одну гортань, а две — верхнюю (larynx) и нижнюю (syrinx). Главная роль в образовании звуков принадлежит нижней, устроенной очень сложно. Само её наличие — отличие птиц от других животных. Располагается она в нижней части трахеи там, где трахея разветвляется на два главных бронха[81].
Нижняя гортань имеет два или четыре вибратора, работающих независимо друг от друга, что позволяет птице изображать из себя дуэт или квартет. Также она позволяет использовать в качестве сильнейшего резонатора трахею. У многих птиц последняя сильно увеличивается в длину и диаметре, увеличиваются также и бронхи, имеющие каждый по независимому источнику звука. Птица способна значительно изменять форму сложной системы голосового аппарата при помощи движений тела и натяжения специальных мышц. Это позволяет ей управлять высотой и тембром своего голоса[81]. Ритмические характеристики звука определяются рефлекторным содружеством нижней и верхней гортани. Верхняя в выступает в роли стоп-крана на пути звукового потока[81]. Голосовой аппарат занимает значительную часть тела, что особенно характерно для маленьких птичек, у которых в процесс пения вовлекается весь организм[81].
Кровеносная система и кровообращение
Кровеносная система имеет два круга кровообращения. Сердце четырёхкамерное с полным разделением артериальной и венозной крови. В отличие от млекопитающих, у птиц главной артерией является правая (а не левая[К 2][57]) дуга аорты, с которой начинается большой круг кровообращения[62]. Проходящая через конечности кровь далее попадает в почечно-портальную систему, а затем в нижнюю полую вену[57]. От дуги аорты отходят парные безымянные артерии, которые затем делятся на сонные артерии, мощные грудные и подключичные артерии, идущие к грудным мышцам и крыльям. От спинной аорты ответвляются артерии к туловищу птиц и к ногам. Венозная система птиц в основном сходна с таковой у пресмыкающихся[57].
Высока у птиц частота сердечных сокращений, она также выше у более мелких видов. В полёте, по сравнению с покоем, пульс заметно увеличивается. Так, у серебристых чаек в покое частота сердечных сокращений достигает 250 в минуту, а при беспокойстве возрастал до 450 уд\мин, в планирующем полёте — 200—300 уд\мин[60]. У воробья в покое пульс составляет около 460 ударов в минуту, а в полёте — около 1000 уд\мин[60].
Нервная система
Нервная система в связи с приспособлением к полету и обитанием в разнообразных условиях развита у птиц значительно лучше, чем у пресмыкающихся[57]. Головной мозг намного превосходит по размерам мозг пресмыкающихся. Продолговатый мозг развит очень хорошо: здесь находятся центры, регулирующие дыхание, кровообращение, пищеварение и другие жизненно важные функции, которые у птиц осуществляются особенно интенсивно. Масса головного мозга составляет у бескилевых птиц и пингвинов — 0,04—0,09 % от массы тела, а у остальных летающих птиц — от 0,2 до 5—8 % от массы тела[82].
Мозжечок велик, так как его роль при полёте, связанном с активной работой мышц и необходимостью постоянного сохранения равновесия тела, очень значительна. Средний мозг покрыт сверху сильно развитым передним мозгом и виден только по бокам. Окончания зрительных нервов, находящиеся в этом отделе мозга, связаны с корой переднего мозга, поэтому зрительная память у птиц хорошо развита. Передний мозг сильно развит, это самая большая часть головного мозга[62].
Кора переднего мозга у птиц выражена лучше, чем у пресмыкающихся, но её слой довольно тонок. Хорошо развиты полосатые тела — главная часть переднего мозга, в которых расположены центры регуляции зрения, движения и так далее. Обонятельные доли малы и тесно примыкают к большим полушариям, поэтому способность птиц к восприятию запахов ограничена[57]. У птиц, питающихся падалью, обонятельные доли развиты гораздо лучше. Полушария передней части мозга гладкие, без извилин и по сравнению с млекопитающими относительно невелики. В их функцию входят управление поведением, ориентация в пространстве, употребление пищи, спаривание и способность строить гнёзда. Высшая нервная деятельность осуществляется не в неокортексе, как у млекопитающих, а в гиперстриатуме[83]. За координацию движений отвечает мозжечок, расположенный в задней части головного мозга. У подавляющего большинства видов плохо развито чувство обоняния, они практически не различают запахи. Среди исключений — киви[84], американские грифы[85] и трубконосые[86].
Спинной мозг в связи с отсутствием хвоста является относительно коротким. Хорошо выражены утолщения в плечевой и поясничной областях. Отходящие от них спинномозговые нервы при выходе из позвонков образуют плечевое и поясничное сплетения, ветви которых иннервируют мускулатуру соответствующих конечностей[57].
Интеллект птиц
Хорошо развитый головной мозг позволяет птицам вырабатывать сложные формы поведения и приспосабливаться к самым разным ситуациям. Наиболее яркой демонстрацией умственных способностей птиц считается обучение певчих птиц пению, повторение человеческой речи попугаями, приемы добывания корма у ряда видов и умение вороновых решать сложные задачи, предлагаемые им в специальных экспериментах.
Так, например, в Англии синицы, обитавшие около загородных домов, научились снимать алюминиевые крышки с выставленных бутылок с молоком и продырявливать крышки и склевывать сливки[69]. Описало для птиц и использование «инструментов». Дятловый древесный вьюрок (Camarhynchus pallidus), обитающий на Галапагосских островах держа в клюве крупную иглу кактуса или сухую веточку, ковыряет ею в трещинах коры для извлечения насекомых из глубоких расщелин в древесине. Перелетая с дерева на дерево, вьюрок часто переносит колючку с собой. В Африке обыкновенный стервятник, найдя страусовое яйцо, скорлупу которого он не в состоянии разбить клювом, разыскивает камень (иногда массой до 0,5 кг при массе птицы 2—2,5 кг) — и, многократно бросая его клювом на яйцо, раскалывают скорлупу. Иногда вороны и крупные чайки, схватив твердую добычу (моллюски, крабы и др.), взлетают и затем бросают добычу на землю. Прием повторяется многократно, пока раковина или панцирь не расколется. Также поступают и некоторые хищные птицы с черепахами (стервятники) или с крупными костями (бородач). Различные виды дятлов используют щели в стволах деревьев для закрепления раздалбливаемых ими шишек.
Органы чувств
Из органов чувств глаза являются основным органом ориентировки птиц. Многие птицы хорошо видят вдаль (сокол-сапсан способен увидеть небольшую птицу на расстоянии более 1 км). У некоторых видов поле зрения достигает почти 360°. Зрение у птиц заметно более острое, нежели чем у других групп позвоночных — это объясняют значительно большим количеством светочувствительных клеток в сетчатке глаза. Глаза у большинства видов малоподвижны, поэтому птицам приходится часто двигать головой, чтобы хорошо рассмотреть окружающую среду (исключением является большой баклан)[87] Как правило, зрение монокулярное, зато с очень большим полем зрения — у обыкновенного и американского вальдшнепов его угол может достигать 360° в горизонтальной плоскости и 180° в вертикальной, охватывая, таким образом, всю сферу[88]. Постоянное бинокулярное зрение, как у человека, имеется только у совоообразных[89]. Глаза нырковых птиц имеют особые гибкие хрусталики, приспособленные к зрению как в воздухе, так и под водой[62]. Важной особенностью зрения птиц является его четырёхкомпонентность — сетчатка глаза способна улавливать не только цветовую модель, состоящую из красного, зелёного и синего цветов, но также лучи ближнего ультрафиолета[90]. Кроме того, криптохром, служащий для восприятия синего цвета, воспринимает магнитное поле Земли, позволяя его буквально видеть; молекула быстро переключается между сигнальным и неактивным положением благодаря присутствию супероксида, содержащегося в чуть более высокой концентрации, чем у млекопитающих[91].

Веки неподвижны, мигание осуществляется с помощью особой перепонки («третьим веком»), которая располагается в переднем углу глаза и двигается по горизонтали[92] .У многих водных птиц перепонка полностью закрывает глаза и под водой выполняет функцию контактной линзы[62].
Ушные раковины отсутствуют, уши прикрыты перьями. В лабиринте внутреннего уха имеется один завиток улитки, отходящий от нижнего мешочка. Среднее ухо содержит одну косточку, которая передает в лабиринт звуковые волны; барабанная перепонка лежит в небольшом углублении, обрамление у некоторых птиц наружного слухового отверстия перьями создает как бы ушную раковину. У ушастых сов, филинов и со’вок (Otus) эти перья, называемые кроющими, вытянуты и внешне напоминают уши[93].. Прекрасно развито у птиц восприятие изменений равновесия тела, благодаря хорошему развитию соответствующих частей лабиринта.
Органы обоняния и вкуса сравнительно слабо развиты. Органы осязания рассеяны в разных местах поверхности кожных покровов
Половая система
Наружные половые органы отсутствуют. Органы размножения самца представлены парными семенниками бобовидной формы, лежащими в брюшной полости. От них отходят семяпроводы, открывающиеся в клоаку. Размеры семенников изменяются в течение года: в период размножения их вес может увеличиваться в 250—300 раз. У большинства птиц внутреннее оплодотворение обеспечивается тем, что отверстие клоаки самца прижимается к клоаке самки в которую впрыскивается порция спермы. У некоторых птиц (страусообразные, тинаму, гусеобразные и краксы из курообразных) от стенки клоаки обособляется непарный совокупительный орган, имеющий фиброзные тела; при совокуплении он выдвигается и вводится в клоаку самки[57][60].
Органы размножения самок представлены обычно только одним, левым, яичником, лежащим вблизи почки. У самок некоторых попугаев и хищных птиц иногда бывает и правый яичник. Яичник имеет зернистое строение. Формирование фолликулов заканчивается в первые месяцы постэмбриональной жизни (у галки образуется около 26 000 фолликулов). Ежегодно к началу размножения начинается рост немногих фолликулов (в 5—10 раз больше нормального числа откладываемых яиц), яичник приобретает гроздевидную форму. Яйцевод представляет собой длинную трубку, открывающаяся воронкой в полость тела около яичника; другой конец яйцевода открывается в клоаку[60]. Вышедшее из яичника яйцо попадает в непарный яйцевод, в верхней части которого и происходит оплодотворение. Пройдя по яйцеводу, яйцо приобретает свою белковую оболочку, а попав в более широкую матку, покрывается скорлупой. Через влагалище — конечный отдел половых путей самки — яйцо попадает в клоаку, а оттуда уже выводится наружу[57].
Образ жизни
Жизнь птиц слагается из многократно повторяющихся явлений, среди которых можно различить суточные и годичные периодические явления. Суточные представляют собой чередование периодов деятельности и отдыха, годичные гораздо сложнее, отражают на себе приспособление организма к жизни в известных условиях и часто могут быть поняты лишь путём сравнительного изучения. Таковы явления гнездостроения, линьки и странствования птиц.
Питание
Спектр питания птиц довольно является широким и включает разнообразные растительные и животные корма. По разнообразию используемых кормов птиц разделяют на три группы[94]. Полифаги (всеядные) питаются разнообразными растительными и животными кормами. К этой группе можно отнести примерно треть всех семейств, причем в пределах каждого семейства всеядность сильнее выражена у более крупных видов. Примером наиболее типичных птиц-полифагов могут быть крупные вороновые птицы, крупные чайки, журавли и др.
Противоположной группой являются стенофаги — птицы, потребляющие однородные корма и использующие однообразные приемы ловли добычи. Стенофагия встречается среди птиц редко. К стенофагам следует относятся стрижи и многие козодоев, питающихся только летающими насекомыми, и ласточек, которые ловят насекомых в воздухе, но могут и склевывать их на лету с растений. К этой же группе относятся типичные падальщики — грифы и марабу, а также виды, питающиеся только крупной рыбой — пеликаны, скопа, бакланы, орланы и др. Африканский пальмовый гриф питается преимущественно плодами нескольких видов пальм; американский коршун-слизнеед поедает практически лишь моллюсков. Нектаром цветов ограниченного числа растений питаются некоторые виды мелких колибри, медососов, нектарниц и гавайских цветочниц[95]. К стенофагам относятся и клесты, питающиеся семенами хвойных деревьев.
Большинство птиц относится к промежуточной группе — они питаются довольно широким набором различных кормов. Таковыми являются многие воробьинообразные, питающиеся различными насекомыми и семенами. Некоторые колибри, медососовые и нектарницевые питаются не только нектаром, но и тычинками цветков и насекомыми. Рыбой и разнообразными, крупными водными беспозвоночными питаются бакланы, поганки и многие другие птицы; зелеными частями растений, семенами, ягодами и различными беспозвоночными — курообразные, туканы, птицы-носороги и др.
По составу корма птиц, также разделяют на ряд экологических групп. Виды, питающиеся преимущественно растительной пищей (фитофаги). Почти исключительно листьями питается гоацин; гуси, лебеди, некоторые утки, лысухи питаются преимущественно разнообразной прибрежной и водной растительностью, попутно поедая водных животных. Зеленые части растений, ягоды, семена, почки — основа рациона питания курообразных. Преимущественно семенами питаются многие воробьинообразные — ткачиковые, вьюрковые, жаворонковые и др. Семенами, ягодами и плодами питаются многие попугаи, голуби, турако, птицы-мыши, многие трогоновые, райские птицы. Однако большинство таких птиц выкармливает своих птенцов преимущественно животными кормами.
Виды, питающиеся преимущественно животной пищей, называют зоофагами, хотя многие из них в небольшой степени поедают и растительные корма. Почти треть всех семейств птиц являются исключительно или преимущественно насекомоядными (энтомофаги). Однако, насекомыми той или иной степени питаются почти все птицы. Многие водные и околоводные птицы питаются преимущественно рыбой, попутно поедая и водных беспозвоночных.
Многие хищные птицы и совы питаются преимущественно мелкими грызунами. Немногие хищные птицы, например сокол сапсан, ястребы и чеглок, болотный лунь и некоторые другие питаются главным образом другими птицами. Небольшое число видов птиц относится питаются преимущественно моллюсками (например пастушковый журавль арама). К герпетофагам, птицам питающимся амфибиями и рептилиями относятся орел-змееяд, птица-секретарь, кариамы, некоторые крупные зимородки (кукабары) и др. Некоторые чайки и качурки относятся к копрофагам — они могут поедать помёт китообразных и ластоногих. Гарпии ловят обезьян и ленивцев, красногорлый сокол и коршун Macheiramphus alcinus охотятся преимущественно на летучих мышей. Малайский орел-яйцеед (Ictinaetus malayensis) таскает яйца из гнезд других птиц.
В связи с сезонностью появления разных типов кормов для многих видов птиц характерно сезонное изменение питания. Степень его изменчивости определяется характером пищевой специализации. Часто переход с одного пищевого режима на другой зависит от доступности определённого корма, что особенно характерно для зимующих и перелётных птиц северного полушария. Так, обыкновенная иволга в период размножения питается главным образом гусеницами и прочими насекомыми, а в остальное время года переключается на ягоды и сочные плоды растений[96] Большой пёстрый дятел на Британских островах зимой питается древесными насекомыми, а в более холодном климате Финляндии семенами хвойных деревьев.[94].
У многие птицы, обитающие на больших ареалах, характеризуются выраженной географической изменчивостью рациона питания, определяемой наличием и доступностью определённого корма. Отличия в питании могут быть заметными как между популяциями, населяющими удалённые районы, так и обитающими на соседних территориях.
Некоторым видам птиц свойственно запасание корма. Наиболее примитивна форма запасания пищи присуща филину и некоторым крупным совам: поймав крупную добычу, хищник относит её в укрытие и сидит на ней, пока не съест. Гораздо более сложно выражено запасание пищи у воробьиного сычика и мохноногих сычей. Осенью они складывают излишки пойманных мышевидных грызунов в дупла. После выпадения снега, когда отлов добычи становится затруднительным, птицы используют эти запасы. Кедровки расклевывают шишки кедровой сосны и собирают орешки, затем пряча свою добычу в дуплах и т. п. При урожае желудей сойки прячут их среди опавших листьев, мха, в прогнивших пнях, дуплах и т. п. Желудевый дятел (Melanerpes formicivorus) из США выдалбливает в стволах деревьев и деревянных столбах маленькие отверстия, в которые забивает собираемые им жёлуди. В одной жёлтой сосне было обнаружено до 60 тыс. спрятанных таким образом жёлудей[69]. Подобные запасы используются птицами в зимнее время.
Пение птиц
Звуковые сигналы имеют в жизни птиц исключительно важное значение. Ими обеспечиваются охрана своей территории от вторжения чужаков (то есть, по сути, обеспечение пищевых ресурсов), привлечение самки для выведения потомства, предупреждение сородичей и птенцов о грозящей опасности. В языке птиц насчитываются десятки звуковых сигналов (бедствия, предостережения, пищевые, ухаживания, спаривания, агрессивные, стайные, гнездовые и так далее)[97].

Разнообразие и музыкальность голосов птиц широко используются в музыкальном творчестве. Однако человек не может расслышать многие высокие обертоны в голосе птиц (уходящие в ультразвуки), равно как и быстрые трели (модуляции, громкости и высоты звуков, превышающие 10 изменений в секунду). Птичье щебетание часто состоит из модуляций порядка 100—400 изменений звука в секунду[98].
Ультразвуковые обертоны голоса у певчих птиц простираются до 50 000 колебаний в секунду. Пение некоторых видов целиком состоит из ультразвуков. Возможности птичьего голосового аппарата в плане ритмических модуляций в 50—100 раз превышают возможности слуха человека.
Также птицы используют другие способы звукоизвлечения: с применением клюва, лап, крыльев, хвоста или посторонних предметов (как, например, дятлы).
Песню птицы принято делить на отдельные элементы, называемые нотами или слогами. Большинство песен состоит из 1—2 слогов, однако бывает и больше (13—24 слога у пеночки-теньковки). Более сложные части, состоящие из нескольких слогов, называются мотивами, фразами или коленами (как у соловья)[97].
Птичья песня иногда имеет широкий смысл и понимается другой птицей по ситуации. Одна и та же песня в период размножения может служить сигналом привлечения самки, а в период высиживания птенцов — сигналом занятости территории. Сигнальное значение несут не все элементы песни; часто фразы прибавляются для самовыражения птицы[97].
Важно то, что птичья песня видоспецифична, что обеспечивает биологическую устойчивость и сохранность вида. Но особи одного вида, находящиеся на слишком большом расстоянии друг от друга, нередко вырабатывают свои особенности языка и теряют способность понимать сородичей. Благодаря индивидуальности и хорошему слуху птицы различают близких сородичей по характеру песни (например, супруга). Наряду с видоспецифичностью песни птиц нередко имеют и межвидовые сигналы. Примером этого служит крик синицы, обнаружившей сову — за ним следует сбор к этому месту множества других птиц для выражения агрессии по отношению к сове[97].
В вокализации выделяют пение и голосовые сигналы, отличие между которыми основывается на модуляции, длине и контексте звуков. Пение или песня длинная и более сложная и ассоциируется с брачным и территориальным поведением, тогда как голосовые сигналы или призывы выполняют функции предупреждения или удержание стаи вместе[99][100]. Пение наиболее развито у птиц отряда воробьинообразных. Большей частью пение свойственно самцам, а не самкам, хотя встречаются исключения. Пение чаще всего издается, когда птица сидит на каком-либо субстрате, хотя некоторые виды способны издавать его во время полета. Некоторые группы птиц практически молчаливы, издают лишь механические звуки, например аист, лишь щелкают клювами. У некоторых манакинов (Pіprіdae) самцы развили несколько механизмов образования таких звуков, включая стрекотание, характерное для насекомых[101].
Образование звуков механическими средствами в отличие от сиринкса носит название инструментальной музыки (по определению Чарльза Дарвина) или механических звуков[102] и в работах современных авторов, сонации[103]. Термин сонация означает как акт образования невокальных звуков, которые образовываются с определенным назначением, и являются коммуникационными сигналами, которые образовываются неголосовыми структурами, такими как клюв, крылья, хвост и перья[103].
Многие птицы способны перенимать песни других видов и имитировать голос человека (включая индивидуальные особенности его тембра, как это делают скворцы, попугаи, вороны и др.)[81]. Название пересмешник (Mіmus) было дано этой птице за способность копировать звуки других птиц и воссоздавать их. Другим известным видом, способным к копированию, является обыкновенный скворец (Sturnus vulgarіs). Хорошими способностями для имитации языка человека владеют попугай жако (Psіttacus erіthacus), австралийские попугаи, такие как большой желтохохлый какаду и южноамериканские амазоны (Amazona). В Европе случаи способности имитировать голос человека были известны среди некоторых представителей семейства врановых, такие как галка (Corvus monedula), сорока (Pіca pіca) и ворон (Corvus corax)[104].
Также существуют случаи присвоения голосовых сигналов, а не пения птиц. Например, толстоклювая эуфония (Euphonia laniirostris) часто издаёт сигнал угрозы других видов во время приближения хищника к гнезду. В других случаях имитация используется для заманивания жертвы, например панамский черноспинный лесной сокол (Micrastur mirandollei) способен имитировать призывы своих жертв, а потом ловит птиц, которые прилетели в ответ на призыв[105].
Передвижение
 Золотистая щурка (Merops apiaster)
Золотистая щурка (Merops apiaster) Колибри (Archilochus colubris) может зависать на одном месте, производя до 52 взмахов крыльями за секунду
Колибри (Archilochus colubris) может зависать на одном месте, производя до 52 взмахов крыльями за секунду Императорский пингвин (Aptenodytes forsteri)
Императорский пингвин (Aptenodytes forsteri)Полёт является основным способом передвижения большинства видов птиц, помогающий им искать пищу, мигрировать и спасаться от хищников. Только небольшое количество видов вторично утратили эту способность. По ветвям, по земле и воде птицы передвигаются при помощью задних конечностей. Только у стрижей все четыре пальца лапы направлены вперёд, заканчиваются острыми изогнутыми коготками и служат только для цепляния и опоры; передвигаться по земле и по ветвям они не могут.
Полёт является наиболее специфической формой передвижения птиц, определившей основные черты организации этого класса. Способность птиц к полёту вызвала ряд адаптаций для обеспечения полёта, взлёта и посадки, ориентирования в пространстве и навигации. Характеристики полёта птиц в значительной мере зависят от величины птицы и её экологической ниши. Хотя определённые биологические адаптации (к примеру, уменьшение массы) характерны для всех птиц, другие (например, форма крыльев) характерны только для отдельных групп. Полёт птиц принято разделять на два основных типа[106]:
- активный, или машущий
- пассивный, или парящий
Машущий полёт состоит из двух отдельных типов движения: рабочего хода и обратного хода. Во время рабочего хода крыло двигается вперёд и вниз, а обратный ход возвращает крыло в начальную позицию. При этом внутренняя часть крыла в первую очередь генерирует подъёмную силу, тогда как кисть генерирует тягу, которая толкает птицу вперёд. Во время рабочего хода маховые перья первого порядка сведённые вместе, формируют плотную обтекаемую поверхность крыла. Наоборот, во время обратного хода маховые перья первого порядка многих, особенно небольших, птиц поворачиваются вокруг своей оси, обеспечивая движение воздуха между ними. Крупные птицы или длиннокрылые маленькие птицы полностью или частично сгибают крылья, приближая их к туловищу[107]. Парящим называется полёт без активных затрат энергии со стороны птицы, который осуществляется или за счёт потери скорости или высоты, или за счёт использования движения воздуха для получения энергии. Различают парение динамическое и статическое. В основе статического парения птиц лежит использование потоков обтекания или воздушных термических потоков. Ещё один тип парения связан с использованием птицами термиков — потоков восходящего воздуха, которые возникают вследствие нагрева воздуха возле поверхности земли. Этим методом пользуется много крупных птиц, удерживаясь в центральной части термика за счёт кружения на месте. Такое поведение характерно для многих хищных птиц, в частности грифов, коршунов, канюков. Этим методом также пользуются и аисты, пеликаны и другие птицы, которые не являются хищными.
Птицы обычно используют не один тип полёта, а комбинируют их. За взмахами крыльев следуют фазы, когда крыло не совершает движений: это скользящий полёт, или парение. Такой полёт характерен преимущественно для птиц средних и крупных размеров, с достаточной массой тела[106].
Зависание на месте относительно окружающего воздуха (англ. hovering) является сложной задачей для птиц. Большинство птиц или совсем неспособны к нему или способны лишь на протяжении очень незначительного промежутка времени. Фактически колибри являются единственной группой птиц, приспособленных к этому. Отличительными являются зависания относительно поверхности за счёт полёта против ветра со скоростью ветра (англ. windhovering). Такое поведение характерно для многих хищных птиц и птиц, которые специализируются на охоте и ловле рыбы: буревестник, скопа, крачки, поморники, некоторых зимородков.
Некоторые виды птиц, прежде всего обитающие на изолированных островах, где отсутствуют наземные хищники, потеряли способность к полёту. Это является доказательством того, что, несмотря на большие преимущества полёта, он требует больших затрат энергии, и поэтому, при отсутствии хищников, может стать ненужным[60][108].
Большинство мелких древесно-кустарниковых птиц по земле, и в ветвях передвигаются прыжками. Птицы средних размеров (голуби, вороны, и др.) и некоторые мелкие птицы (трясогузки, и др.) по земле ходят и бегают, попеременно переставляя ноги. Ряд видов чередуют ходьбу и прыжки. У птиц наземного образа жизни (фазановые, тинаму, дрофы, рябки и др.) крепкие пальцы относительно короткие и толстые, задний палец часто мал и не служит для опоры. По земле они ходят и бегают, попеременно переставляя ноги; на ветви садятся лишь немногие виды[60].
 Оляпка — одна из немногих воробьиных птиц, которая может нырять, часто погружаясь в воду и бегая по дну, хорошо плавая, гребя под водой крыльями.
Оляпка — одна из немногих воробьиных птиц, которая может нырять, часто погружаясь в воду и бегая по дну, хорошо плавая, гребя под водой крыльями.Любая птица в воде не тонет и при необходимости, плывет[60]. Могут присаживаться на воду дрозды и голуби[60], довольно часто плавают пастушки и многие кулики, иногда садятся на воду и плавают цапли и т. д. Но у птиц, постоянно связанных с водой, развиваются приспособления облегчающие плавание, а у многих видов — и ныряние[60]. Низкий удельный вес птиц, облегчая плавание, затрудняет ныряние. Некоторые птицы при ловле добычи способны нырять с пикирования: птица летит над водой и, заметив у добычу, полускладывает крылья и резко пикирует, погружаясь в воду на небольшую глубину за счет инерции. К такому нырянию прибегают птицы, которые могут плавать — трубконосые, многие веслоногие (фрегаты, фаэтоны, чайки, крачки, поморники, олуши, американский бурый пеликан,). Также ныряют птицы, обычно не садящиеся на воду — скопа, орланы, рыбные филины, многие зимородки. По-настоящему погружаться на большую глубину и активно передвигаться в толще воды может относительно небольшое число видов хорошо плавающих птиц. Обычно птицы могут нырять на глубину нескольких метров. Некоторые нырковые утки (гаги, морянки и др.), кайры, гагары, поганки, пингвины ныряют иногда на глубину 50—60 м и крайне редко—несколько глубже[60]. Зарегистрированное погружение императорского пингвина на глубину 265 м.[60] видимо, является пределом для ныряющих птиц. Продолжительность погружения обычно длится не более к 1—1,5 мин[60], но птицы могут находиться под водой даже до 10—15 минут (пингвины, гагары)[60]. Из воробьиных птиц ныряют оляпки. Сохраняя типичный облик воробьиных, они отличаются имеют более плотным оперением, сильно развитым пухом, укороченными крыльями и хвостом. Способ ныряния оляпок совершенно не похож на ныряние других птиц. Опустившаяся на воду оляпка так держит крылья, что течением её прижимает ко дну, она бежит, переворачивая камешки и склевывая добычу под ними. К такому своеобразному нырянию в быстро текущую воду и беганью по дну способны также вилохвостки (Enicurus) из семейства дроздовых и водяные печники (Cinclodes)[60].
Размножение
Характерная черта размножения птиц — яйцекладка. Яйцеклетка птиц увеличивается и превращается в яйцо, которое отличается большим размером и содержит запас питательных веществ, необходимый для развития зародыша. Половые органы птиц расположены внутри, открываясь напрямую в клоаку. Оплодотворение происходит внутри, после яйцекладки для продолжения развития зародышу необходимо тепло, поэтому родители согревают его теплом своего тела на протяжении нескольких недель или даже месяцев. В зависимости от продолжительности и сложности эмбрионального развития, птицы подразделяются на два класса — выводковые и птенцовые:
- Выводковые птицы — птицы, птенцы которых вылупляются из яйца вполне сформированными, одетыми пухом и способными отыскивать корм. Они тут же покидают гнездо, хотя ещё долгое время следуют за своими родителями, которые их защищают и помогают отыскивать корм.
- Птенцовые птицы — птицы, птенцы которых вылупляются из яйца несформированными, голыми, слепыми и беспомощными. Они долго остаются в гнезде. Родители не только защищают их, но также и кормят из клюва.
- Полувыводковые птицы — смешанный тип развития, при котором птенцы появляются отчасти сформированными, но долгое время остаются в гнезде и получают пищу от родителей.
За несколько дней до вылупления практически сформировавшиеся птенцы устанавливают с родителями звуковую связь. В этом общении выделено около десятка сигналов. Таким образом птенец усваивает сигналы матери и основные ситуации внешней среды, что подготавливает его к жизни после вылупления[109].
Забота о потомстве

Этот раздел статьи ещё не написан. Согласно замыслу одного из участников Википедии, на этом месте должен располагаться специальный раздел.
Вы можете помочь проекту, написав этот раздел.Гнездование
Гнездостроение представляет собой чрезвычайно сложное явление. Вообще, птицы устраивают гнездо, чтобы поместить в нём яйца и воспитать птенцов, но, с одной стороны, многие птицы совсем не устраивают гнезда, с другой — очень многие не воспитывают в гнезде птенцов, оставляющих его сейчас же после вылупления из яиц. Этим обуславливается то, что те птицы, которые принадлежат к группе птенцовых, вовсе не всегда строят и наиболее искусные гнезда. Но многочисленные факты указывают, что не только строение гнёзд, но даже и откладываемые птицами яйца находятся в самом тесном соотношении с теми условиями, среди которых протекает жизнь птиц. Таким образом, одна и та же птица в разных условиях строит и более или менее различные гнезда; и кроме того молодые особи вообще являются худшими строителями гнёзд, нежели старые. Из этого следует, что строение гнёзд никак не является исключительно инстинктивным актом.
Некоторые из чистиков кладут своё единственное яйцо на голом выступе скалы, откуда яйцо не сносится ветром благодаря своей конической форме. Авдотка и козодой откладывают яйца прямо на почву, безо всякой подстилки, но выбирают такие местечки, что яйца совершенно сливаются с ними благодаря своей окраске. Киви откладывает своё непропорционально большое яйцо между корнями папоротников. Самки страусов откладывают яйца в песок, устраивая род общественного гнезда, забота о котором падает на самца. Некоторые из сорных кур зарывают яйца в песок, предоставляя вывод птенцов солнечной теплоте, другие зарывают их в кучи гниющих веществ. Гагары устраивают плавучие гнезда. Голуби довольствуются возведением на деревьях лёгкого помоста из веток. Береговая ласточка, зимородок вырывают норки, где помещают гнёзда в самой отдалённой части такого хода. Ласточки и стрижи лепят гнёзда из грязи, а саланганы из комочков быстро затвердевающей на воздухе слюны. В гнездостроении большинства воробьиных слюна играет очень важную роль, так как при помощи её птица не только размачивает, но и склеивает разные волоконца, стебельки и веточки.
Многие птицы, как иволги, обыкновенный ремез и др., строят висячие гнезда, прикрепляя их самым различным способом к ветвям и даже отдельным листьям (колибри). Поползни и дятлы гнездятся в дуплах, так называемая печная птица Южной Америки устраивает из глины огромное шаровидное гнездо на корнях мангровых деревьев. Многие птицы, как грачи, розовые скворцы, щурки, гнездятся колониально, но особенно замечателен в этом отношении южноафриканский общественный ткач. Последний строит на акациях гнездо рядом с гнездом, наваливая огромную кучу растительного материала, под которым, наконец, дерево рушится. Замечательные австралийские беседковые птицы устраивают беседки (Ptilorhynchus, Chlamydera, Sericulus), куда самцы сходятся, чтобы привлечь к себе самок, другие (Amblyornis — Новая Гвинея) с той же целью устраивают сады. Обыкновенная кукушка представляет собой любопытный пример паразитизма, так как несёт яйца в гнёздах других птиц.
Гнездование характеризует собой в жизни птиц один из самых важных периодов, так называемый брачный, то есть период спаривания самца с самкой, устройства гнезда, вывода и воспитания молодых. Большинство птиц гнездится в течение года один раз, многие по два и более. Перелётные птицы на зимовках не гнездятся. Спаривание сопровождается пением самцов, а у полигамных птиц, каковы, например, тетерева, так называемым токованием. Последним названием определяется собрание многих самцов на одном месте, токовище, где они состязаются друг с другом из-за обладания самками. Часто токование сопровождается ожесточёнными битвами соперников. Нормально у полигамных птиц, что самец не принимает участия в воспитании детей. Напротив, у моногамов он всецело разделяет заботы самки.
У птиц-носорогов после того как самка отложит яйца, самец заделывает отверстие дупла с гнездом глиной, оставляя лишь узкую щель, через которую проходит клюв «пленницы». Самка насиживает яйца и продолжает оставаться в дупле ещё несколько недель после вылупления птенцов. Всё это время самец обеспечивает семейство кормом. Такие меры безусловно защищают самку и птенцов от хищников, однако они же доставляют самке проблемы с уходом за гнездом[110].
Когда птенцы выведены и воспитаны, птицы приступают к линьке, то есть к смене старых перьев на новые.
Экология
Этот раздел статьи ещё не написан. Согласно замыслу одного из участников Википедии, на этом месте должен располагаться специальный раздел.
Вы можете помочь проекту, написав этот раздел.Взаимоотношения с другими организмами
См. также: Муравление и Ant-follower Чёрный дронго (Dicrurus macrocercus) во время муравления
Чёрный дронго (Dicrurus macrocercus) во время муравленияБолее чем 250 видов птиц занимаются муравлением[111]. Они отдыхают на муравьиных гнёздах, где муравьи, забравшись к ним на крылья и перья, удаляют эктопаразитов. Другие птицы «купаются» в муравейниках или раздавливают муравьёв и смазывают ими оперение, преследуя аналогичные цели[112][113][114]. При этом птицы выбирают муравьёв (из подсемейств формицины и долиходерины), которые выбрасывают струи муравьиной кислоты, обладающей инсектицидными свойствами.
Кочевые муравьи, ведущие кочевой образ жизни, известны своими «рейдами»[115]. Подобные скопления муравьёв вида эцитон Бурчелли привлекают птиц, следующих за муравьями, таких как древолазовые или полосатые муравьеловки, которые используют муравьёв как загонщиков насекомых и других мелких членистоногих[116]. Наиболее известные представители принадлежат к семейству полосатых муравьеловок (Thamnophilidae), хотя подобное поведение также описано для представителей семейств древолазовых, мухоловковых, кардиналовых, кукушковых, дроздовых, совиных, бюльбюлевых и некоторых других.
Защита
Почти все птицы при нападении на них активно защищаются. Но специальные приспособления в этом направлении развиты не столько против внешних врагов, сколько против особей того же вида. Большая часть этих приспособлений выработалась в процессе драк самцов или самок в брачный период. Например, шпоры — костные и роговые наросты на плюсне у куриных, на сгибе крыла у паламедей и пастушков[117].
При защите большинство птиц пользуется теми орудиями, которые используются ими при добывании пищи. Во многих случаях эта защита сопровождается устрашающим поведением: гуси злобно шипят, вертишейка ворочает вытянутой шеей и головой, напоминая змею, буревестники и качурки изрыгают на нападающего вонючую маслянистую жидкость, некоторые хищные птицы забрызгивают нападающего своими экскрементами, выпь и филин, поднимая перья, увеличивают размеры тела и принимают угрожающую позу.[117]
 Оборонительная поза исполинского белонога.
Оборонительная поза исполинского белонога. Взятая в руки вертишейка вытягивает и вращает шею, за что получила своё название
Взятая в руки вертишейка вытягивает и вращает шею, за что получила своё названиеБолее пассивные способы защиты сводятся главным образом к покровительственной окраске, широко свойственной как яйцам, птенцам и многим взрослым птицам. Иногда нормальная защитная поза имитирует какой-нибудь предмет, например, высиживающий яйца cерый лесной козодой (Nyctibius griseus) сидит почти вертикально с вытянутой шеей, становясь чрезвычайно похожим с основанием обломанной ветви. Не менее оригинальную защитную позу занимает исполинский белоног (Podargus strigoides) из Австралии[117].
Отдельные виды птиц используют химическую защиту от хищников. Некоторые трубконосые способны выделять из желудка неприятную маслянистую жидкость против агрессора[118]. Обитающие в Новой Гвинее дроздовые мухоловки (Pitohui) выделяют на кожу и перья сильнодействующий нейротоксин[119].
Рябинники защищают своё гнездо от хищников свойственным для них способом: птицы, с резкими звуками «ра-ра-ра…» набрасываются на хищника (млекопитающего, птицу), меча на него жидким помётом[120][121][122].
Естественные враги
Роль в экосистемах
Классификация
Классификация современных птиц
Все современные птицы относятся к иногда выделяемой как подкласс группе веерохвостых птиц — Ornithurae, или Neornithes, которая противопоставляется ящерохвостым птицам — Saururae, или Archaeornithes. К последней группе относят лишь археоптерикса.
Веерохвостые птицы разделяются на 2 подкласса, объединяющих 33 отряда (6 из них только с ископаемыми формами) и 213 семейств (из них 42 — ископаемые). Всего известно около 9000 видов, группируемых в 2000—2400 родов. Распространены практически по всему земному шару.
 Каролинский дятел (Melanerpes carolinus, отряд дятлообразных)
Каролинский дятел (Melanerpes carolinus, отряд дятлообразных)- Подкласс Paleognathae — Бескилевые
- Отряд Казуарообразные (Casuariiformes)
- Отряд Кивиобразные (Apterygiformes)
- Отряд Нандуобразные (Rheiformes)
- Отряд Страусообразные (Struthioniformes)
- Отряд Тинамуобразные, или Скрытохвостые (Tinamiformes)
- † Отряд Эпиорнисообразные (Aepyornithiformes)
- Подкласс Neognathae — Новонёбные
- Отряд Аистообразные, или Голенастые (Ciconiiformes, Gressores)
- Отряд Буревестникообразные, или Трубконосые (Procellariiformes, Tubinares)
- Отряд Воробьинообразные (Passeriformes)
- Отряд Гагарообразные (Gaviiformes)
- † Отряд Гесперорнисообразные (Hesperornithiformes)
- Отряд Голубеобразные (Columbiformes)
- Отряд Гусеобразные (Anseriformes)
- † Отряд Диатримообразные (Diatrymiformes или Gastornithiformes)
- Отряд Дятлообразные (Piciformes)
- Отряд Журавлеобразные (Gruiformes)
- † Отряд Ихтиорнисообразные (Ichthyornithiformes)
- Отряд Козодоеобразные (Caprimulgiformes)
- Отряд Колибриобразные (Trochiliformes)
- Отряд Кукушкообразные (Cuculiformes)
- Отряд Курообразные (Galliformes)
- Отряд Пеликанообразные, или Веслоногие (Pelecaniformes, Steganopodiformes)
- Отряд Пингвинообразные (Sphenisciformes)
- Отряд Поганкообразные (Podicipediformes)
- Отряд Попугаеобразные (Psittaciformes)
- Отряд Птицы-мыши (Coliiformes)
- Отряд Ракшеобразные (Coraciiformes)
- Отряд Ржанкообразные (Charadriiformes)
- Отряд Рябкообразные (Pteroclidiformes)
- Отряд Совообразные (Strigiformes)
- Отряд Соколообразные, или Дневные хищные птицы (Falconiformes)
- Отряд Стрижеобразные, или Длиннокрылые (Apodiformes)
- Отряд Трогонообразные (Trogoniformes)
- Отряд Туракообразные (Musophagiformes)
- Отряд Фламингообразные (Phoeriicopteri)
Птицы и человек
Использование птиц человеком
 Куры
КурыЧеловеком одомашнено несколько видов птиц с целью получения от них яиц, мяса, пера и пуха, а также для использования в религиозных обрядах, для развлечения и в целях коммуникации (почтовый голубь).
Разведение домашних птиц составляет отдельную отрасль сельского хозяйства — птицеводство.
Ловля рыбы бакланами (Phalacrocorax capillatus) практикуется местным населением в 13 районах Японии, а также в ряде мест Китая. Проглотить крупную рыбу птицам мешает надетое на горло веревочное кольцо. Как только рыбак видит, что зоб птицы наполнился рыбой, он вытаскивает баклана из воды за поводок, а затем достаёт рыбу из зоба птицы. В Китае же подход к процессу несколько иной — там бакланов дрессируют, чтобы те сами доставляли рыбу к лодке и выплевывали её из зоба[123][124].
Хозяйственное значение
Антропогенное воздействие на птиц
В последние годы описано множество случаев локальной массовой гибели птиц при использовании человеком различных ядов для борьбы с насекомыми-вредителями и грызунами, при применении некоторых гербицидов, случаи гибели водных птиц при загрязнении нефтью и другими веществами в местах сброса отходов различных производств и т. п. Много птиц гибнет, разбиваясь о маяки и линии электропередач. Большой является разорение гнезд и гибель птенцов луговых птиц во время сенокоса и других сельскохозяйственных работах. В окрестностях крупных городов из-за частого вспугивания птиц нарушается нормальный ход насиживания и выкармливания птенцов. С 1600 года по настоящее время вымерло 94 вида птиц. Примерно четверть из них вымерла по естественным причинам, а гибель остальных связана с деятельностью человека: непосредственным истреблением, уничтожением мест обитания, завозом хищников и т. п. Из этих видов только 9 обитали на материках, а остальные 85 видов обитали на островах Новой Зеландии, Гавайских и ряде других[125].
Охрана
В Красную книгу СССР 1978 года было включено 26 видов птиц, находящихся под угрозой исчезновения, и 37 видов, сохранение популяции которых вызывает сейчас серьезные опасения.
Охота на птиц
 Российская почтовая марка с изображением охоты на кряковых уток
Российская почтовая марка с изображением охоты на кряковых утокПтицы, собирательно называемые в охотничьей практике «пернатая дичь», являются наиболее популярным объектом любительской спортивной охоты. Это обусловлено тем, что птицы более многочисленны, чем четвероногая дичь, они встречаются практически во всех биотопах, охота на них обычно не представляет большой сложности и доступна всем категориям охотников[126][127]. В фауне бывшего СССР к пернатой дичи относятся около 250 видов птиц[126]. «Охотничьей» как правило считается птица, обладающая вкусным мясом, хотя причисление того или иного биологического вида к дичи носит достаточно условный характер и зачастую вызвано традицией — так, во Франции и Италии дрозды считаются ценной дичью, а в России к дичи вообще не относятся. В русскоязычной терминологии пернатая дичь делится на боровую (рябчик, глухарь, вальдшнеп и др.), полевую (перепел, коростель, серая куропатка), болотную (бекас, дупель, гаршнеп и другие кулики, кроме вальдшнепа) и водоплавающую (утки и гуси). Иногда выделяется также горная дичь — кеклики, улары и т. п.[126]. Существует много видов птиц, на которых нет специальной охоты, но которые могут добываться попутно вместе с другой дичью — серая цапля, лысуха, султанская курица и др. Как правило, любительская охота на птицу проводится с помощью гладкоствольного ружья. Стрельба ведётся дробью. Лишь иногда, при больших расстояниях и при охоте на очень крупную птицу (глухарь, дрофа, гуси) охотники применяют нарезное оружие малого калибра, но это разрешено далеко не во всех странах. При охоте часто используют охотничьих собак — легавых, спаниелей, лаек и т. д. Встречаются также любители и энтузиасты охоты на пернатую дичь с использованием специально обученных ловчих птиц. Такая охота традиционно именуется соколиной, хотя, кроме собственно соколов, могут использоваться и другие птицы — кречеты, беркуты и др. В России соколиная охота была особенно популярной среди знати в XVII веке. В царствование Алексея Михайловича для царских соколиных охот существовала развитая инфраструктура, обеспеченная многочисленным и хорошо организованным штатом обслуги. Только в двух сёлах, Коломенском и Семёновском, содержалось более 3 тыс. разных ловчих птиц[128]. В Средние века соколиная охота была весьма широко распространена также в Западной Европе. В настоящее время она сохранилась в качестве дорогостоящего традиционного развлечения преимущественно в некоторых странах Ближнего Востока. Для пополнения запасов пернатой дичи во многих странах (преимущественно в Европе) существует практика её разведения на специальных фермах с последующим выпуском в охотничьи угодья.
Птицы в культуре
В мифологии и религии