- Биолюминесценция
-
 Биолюминесценция обыкновенного светляка
Биолюминесценция обыкновенного светляка
Биолюминесце́нция — способность живых организмов светиться, достигаемая самостоятельно или с помощью симбионтов. Название происходит от греческого слова «биос», что означает жизнь, и латинского «люмен» — свет. Свет создаётся у более высоко развитых организмов в специальных светящихся органах (напр., в фотофорах рыб), у одноклеточных эукариот — в особых органоидах, а у бактерий — в цитоплазме. Биолюминесценция основывается на химических процессах, при которых освобождающаяся энергия выделяется в форме света. Таким образом, биолюминесценция является особой формой хемилюминесценции.
Содержание
История исследований
 Свечение моря, обусловленное динофлагеллятами (гребень волны)
Свечение моря, обусловленное динофлагеллятами (гребень волны)Свечение живых организмов отмечалось ещё античными авторами — Плиний Старший в своей «Естественной истории» упоминал свечение морских организмов[1], многие авторы описывали свечение моря. Однако изучение природы биолюминесценции берёт своё начало в 1668 г., когда Роберт Бойль, крупнейший представитель пневмохимии, изучавший процессы горения, обнаружил сходство между процессами горения угля и свечением гнилушек — Бойль, используя построенный им вакуум-насос, продемонстрировал, что в обоих случаях свечение исчезает, если удалить воздух (то есть кислород).
Пионером в исследовании механизмов биолюминесценции стал Рафаэль Дюбуа, в 1887 г. , поставивший эксперимент с экстрактами из светящихся жуков Pyrophorus — он обнаружил, что экстракт тканей фотофоров светляков, полученный гомогенизацией в холодной воде, светится в течение нескольких минут, однако экстракт, приготовленный в горячей воде, не светится. Вместе с тем Дюбуа обнаружил, что если добавить к потухшему холодному экстракту порцию несветящегося горячего экстракта, то свечение возобновляется. Таким образом, за свечение были ответственны две фракции: устойчивая к нагреву низкомолекулярная, и белковая, теряющая активность при нагревании; свечение in vitro возникало только в присутствии обеих фракций и в присутствии кислорода. Аналогичные результаты Дюбуа получил и при эксперименте со светящимися двустворчатыми моллюсками Pholas dactylus. Такое поведение типично для систем фермент — субстрат, поэтому Дюбуа назвал низкомолекулярную фракцию люциферином, а белковую — люциферазой и постулировал ферментативную природу реакций, вызывающих биолюминесценцию[2][3].
Работы Дюбуа положили основу для дальнейших работ в исследовании биолюминесценции, оказалось, что у различных групп организмов существует множество систем люциферин — люцифераза.
Эдмунд Ньютон Харви (Edmund Newton Harvey) в Принстонском университете начал работы по изучению биолюминесценции ракообразных. В 1920 г. Харви показал различие люциферазных субстрат-ферментных систем различных таксонов: люциферин моллюсков Pholas не светился под действием люциферазы ракообразных Cypridina и наоборот, люцифераза Pholas была неактивна по отношению к люциферину Cypridina.
В 1957 г. был выделен и охарактеризован люциферин светляков, оказавшийся производным тиазола[4].
 Медуза Aequorea victoria
Медуза Aequorea victoriaВ конце 1950-х — начале 1960-х Осаму Симомура в университете Нагоя исследовал механизм свечения остракод Cypridina hilgendorfii, которые использовались во время Второй Мировой Войны японцами как природный люминофор: высушенные рачки при смачивании снова начинали светиться. Ему удалось выделить из них в чистом кристаллическом состоянии новый люциферин, отличающийся от люциферина светляков[5]. В качестве объекта дальнейших исследований биолюминесценции в Принстоне он избрал медузу Aequorea victoria, фотофоры которой излучают зелёный свет. Симомура выделил из медуз экворин — белок, содержащий имидазопиразин целентеразин и показал, что биолюминесценция экворина инициируется ионами кальция, при этом, в отличие от классической биолюминесценции, для излучения света экворином кислород не требовался. Это стало открытием нового класса биолюминесцентных систем — фотопротеинов, в которых светоизлучающий фрагмент является не свободным субстратом — люциферином, а простетической группой, прочно связанной с белком.
Симомура также обнаружил, что выделенный из медузы и очищенный экворин in vitro излучает синий свет, в то время как живая медуза светится зелёным. Дальнейшие исследования показали, что за зелёное свечение ответственен другой белок — GFP (англ. green fluorescent protein — зелёный флуоресцентный белок), флуоресцирующий зелёным светом под действием голубого излучения экворина; и экворин, и GFP в дальнейшем вошли в лабораторную практику молекулярной биологии, первый — как индикатор присутствия ионов Ca2+, второй — в качестве флуоресцентной метки для изучения экспрессии клеточных белков. За работы по GFP Симомура был удостоен нобелевской премии по химии 2008 года.
Физико-химические механизмы биолюминесценции
Хемилюминесценция возникает при многих химических реакциях — например, при рекомбинации свободных радикалов или в реакциях окисления (при свободнорадикальном окислении паров белого фосфора в газовой фазе, окислении люминола в водном растворе и т. п.). В этом случае, как и в реакциях биолюминесценции, выделяющаяся энергия не рассеивается в виде тепла, как это происходит в ходе большинства экзотермических химических реакций, а расходуется на образование одного из продуктов реакции в возбуждённом электронном состоянии. Для излучения света в ходе хемилюминесцентной реакции необходимо выполнение, как минимум, двух условий: во-первых, энергия, выделяющаяся в ходе реакции должна превышать ~41-71,5 ккал/моль и, во-вторых, разница энергий основного и возбуждённого состояния продукта реакции должна быть ниже энтальпии химической реакции.
При соблюдении этих условий возможно образование с достаточно высоким выходом окисленной формы люциферина в возбуждённом состоянии и дальнейший переход в основное состояние с испусканием фотона видимого спектрального диапазона. Отношение числа излученных фотонов к общему числу элементарных актов реакции называется квантовым выходом реакции, квантовые выходы биолюминесценции, в отличие от большинства хемилюминесцентных реакций, очень высоки и достигают значений 0,1-1. Такие квантовые выходы для реакций, протекающих в водных растворах при нейтральных значениях pH необычны для хемилюминесцентных процессов и обусловлены специфичной ферментативной природой окислительных реакций биолюминесценции, катализируемых люциферазными комплексами.
Длина волны излучаемого при биолюминесцентных процессах света зависит от разности энергий основного и возбуждённого состояний окислённых форм люциферинов и связанна с ней отношением
 , полуширина полосы излучения составляет обычно ~50 нм. Поскольку процесс перехода возбуждённое — основное состояние обратим, то спектры флуоресценции оксилюциферинов близки к спектрам биолюминесценции: в обоих случаях излучает молекула оксилюциферина, переведённая в возбуждённое состояние либо вследствие химической реакции (биолюминесценция), либо вследствие поглощения достаточно энергетичного фотона.
, полуширина полосы излучения составляет обычно ~50 нм. Поскольку процесс перехода возбуждённое — основное состояние обратим, то спектры флуоресценции оксилюциферинов близки к спектрам биолюминесценции: в обоих случаях излучает молекула оксилюциферина, переведённая в возбуждённое состояние либо вследствие химической реакции (биолюминесценция), либо вследствие поглощения достаточно энергетичного фотона.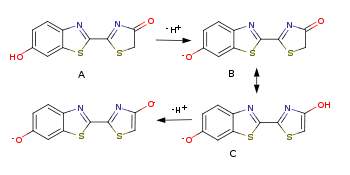 Различные формы оксилюциферина насекомых:
Различные формы оксилюциферина насекомых:
A — нейтральная кетоформа λmax = 618 нм
B — анион (фенолят) кетоформы
C — анион енольной формы, λmax = 587 нм
D — енолят-дианион, λmax = 556 нмВместе с тем, максимум в спектре излучения в биолюминесцентных процессах может изменяться в зависимости от условий протекания реакции. Например, несмотря на то, что химизм биолюминесценции жуков-светляков одинаков и структуры люциферина и оксилюциферина различных видов идентичны, цвет свечения может варьировать от зелёного до красного, то есть максимум в спектре излучения может меняться от 490 до 622 нм. Более того, у личинок бразильских жуков-фенгонид рода Phrixothrix есть несколько органов-фотофоров, испускающих свет различных оттенков — красного фотофоров головы и желто-зеленого фотофоров брюшка[6]. Такое изменение спектра излучения возможно, когда оксилюциферин может существовать в нескольких формах с различной энергией основного состояния, что, в свою очередь, соответствует различающимся энергиям перехода из возбуждённого состояния и, вследствие этого, к различным максимумам в спектре излучения при переходе из возбуждённого состояния в основное.
Оксилюциферин светляков способен к кето-енольной таутомерии и в растворах существует в виде смеси кетонной и енольной форм. Отношение количеств кето- и енольного таутомеров зависит от pH среды: в слабощелочных условиях (pH 7.5 — 7.8 и выше) преобладает енольная форма, при этом максимум в спектре биолюминесценции приходится на 587 нм, то есть на желто-зелёную область, при закислении среды (pH < 6) преобладающей становится кетонная форма и максимум в спектре излучения сдвигается в длинноволновую область до 618 нм, то есть в красную область. При подщелачивании среды образуется енолят-анион оксилюциферина, и максимум в спектре смещается в коротковолновую область до 556 нм. При промежуточных значениях pH в растворе присутствует смесь обеих форм и спектр излучения оказывается бимодальным, воспринимаемый глазом промежуточный оттенок получается вследствие аддитивного смещения желто-зелёного и красного света[7].
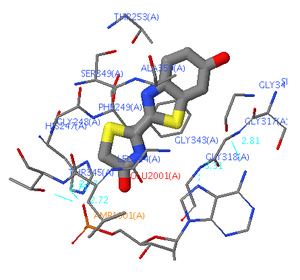 Микроокружение молекулы оксилюциферина в люциферазе светляков Photinus[8].
Микроокружение молекулы оксилюциферина в люциферазе светляков Photinus[8].Другим фактором, влияющим на спектр биолюминесценции, является микроокружение молекулы оксилюциферина в основном и возбуждённом состояниях. На значения энергетических уровней основного и возбуждённого состояний молекулы оксилюциферина в среде оказывает влияние и энергия их взаимодействия с растворителем (энергия сольватации), и образование водородных связей: чем сильнее возбуждённая молекула ассоциирована с микроокружением и чем выше его поляризуемость, тем ниже энергия возбуждённого состояния, тем меньше энергия испускаемого фотона и тем сильнее сдвиг максимума спектра излучения в длинноволновую область.
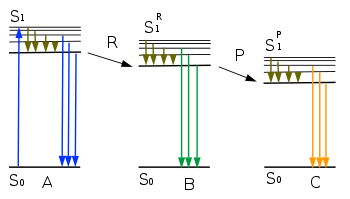 Диаграмма Яблонского для сдвига λmax оксилюциферина:
Диаграмма Яблонского для сдвига λmax оксилюциферина:
A — возбуждённая молекула оксилюциферина в микроокружении молекулы — предшественницы
R — релаксация сольватной оболочки и белкового окружения
B — возбуждённая молекула оксилюциферина в релаксировавшем микроокружении
P — протонирование или таутомеризация
C — таутомер оксилюциферина
Энергии S1 > S1R > S1P, максимумы излучения λAmax < λBmax < λCmaxТретьим фактором, влияющим на энергию возбуждённого состояния оксилюциферина и, соответственно, спектральный максимум, являются релаксационные процессы микроокружения. При отщеплении CO2 от 1,2-диоксетанового предшественника оксилюциферина светляков происходит очень быстрая перестройка электронной структуры молекулы и резкое изменение её дипольного момента, при этом возбуждённая молекула оказывается в сольватной оболочке молекулы — предшественницы. Время жизни молекулы осилюциферина в возбуждённом синглетном состоянии составляет ~ 10−9−10−8 секунды, и если за это время молекулы растворителя или окружающие активный центр белковые цепи люциферазы не успевают переориентироваться в новое равновесное состояние, то энергия возбуждённого состояния оксилюциферина оказывается максимальной, а максимум спектра сдвинут в коротковолновую область, то есть длина волны излучаемого света оказывается зависимой от скорости релаксации микроокружения — и в том числе от подвижности белковых цепей люциферазы[7].
И, наконец, особым случаем, ведущим к изменению спектра биолюминесценции, является переизлучение энергии, выделяемой при окислении люциферинов, флуоресцентными белками — такой механизм наблюдается у некоторых люминесцирующх бактерий и медуз и приводит к смещению спектрального максимума в длинноволновую область. У бактерий, в клетках которых присутствует жёлтый флуоресцентный белок (YFP, англ. yellow fluorescent protein) предполагается индуктивно-резонансный межмолекулярный перенос энергии (механизм Фёрстера) от люциферин-люциферазного комплекса к флуоресцентному белку. Этот механизм может играть весьма существенную роль и становиться основным механизмом биолюминесценции: было показано, что in vitro при добавлении к целентеразиновой люциферин-люциферазной системе полипов-альционарий Renilla reniformis, излучающей с максимумом 480 нм, зелёного флуоресцентного белка Renilla квантовый выход люминесценции на длине волны GFP 510 нм повышается в три раза[9].
Типы люциферин-люциферазных систем
Как уже упоминалось, необходимым условием биолюминесценции является высокая энтальпия реакции окисления люциферина: энергия, выделяющаяся в ходе реакции должна превышать ~41-71.5 ккал/моль, — что соответствует энергиям электромагнитного излучения в видимом диапазоне ~400-700 нм, эта энергия соизмерима с энергией связи C-C в алканах (~79 ккал/моль). Такой энергетический эффект значительно превышает энергетические эффекты большинства биохимических реакций — в том числе и с участием макроэргических соединений — носителей энергии в живых системах; так, например, энергия, высвобождающаяся при гидролизе АТФ до АМФ составляет 10.9 ккал/моль.
 Наиболее распространенный реакционный механизм биолюминесценции: отщепление CO2 от диоксетанона — промежуточного продукта окисления люциферина ведёт к образованию оксилюциферина в возбуждённом состоянии, который переходит в основное состояние с излучением света.
Наиболее распространенный реакционный механизм биолюминесценции: отщепление CO2 от диоксетанона — промежуточного продукта окисления люциферина ведёт к образованию оксилюциферина в возбуждённом состоянии, который переходит в основное состояние с излучением света.Энергия, соответствующая энергиям видимого спектра, в живых системах может быть получена только в реакциях одностадийного окисления с участием молекулярного кислорода (или активных форм кислорода), поэтому большинство люцифераз относятся к классу ферментов — оксигеназ, катализирующих реакции, в которых происходит присоединение кислорода к субстрату-люциферину (за немногими исключениями люцифераз кольчатых червей, обладающих пероксидазоподобной активностью) и, соответственно, все светящиеся организмы являются аэробами.
Большинство люциферинов при окислении образуют циклические напряжённые промежуточные пероксиды — диоксетаноны, в которых валентные углы в четырёхчленном цикле существенно отличаются от нормальных валентных углов, такие соединения далее распадаются с выделением молекулы углекислого газа и образованием возбуждённого кетона-люциферина. Такой механизм реакции характерен для окисления люциферина насекомых и целентеразинов — люциферинов многих морских организмов.
В настоящее время известно пять классов люциферинов различной химической природы: альдегид-флавиновая система бактерий, альдегидные люциферины червей, тетрапирролы динофлагеллят и некоторых ракообразных, имидазопиразолы различных морских организмов и люциферин насекомых — производное тиазола.
Биологические функции
Биолюминесценция выполняет следующие биологические функции:
- привлечение добычи или партнёров
- коммуникация
- предупреждение или угроза
- отпугивание или отвлечение
- маскировка на фоне естественных источников света
Во многих случаях функция биолюминесценции в жизни отдельных светящихся организмов выяснена не до конца, либо вообще не изучена.
См. также
- Люминесценция
- Флуоресценция
- Фосфоресценция
- Сонолюминесценция
- Электрофосфоресценция
- Хемилюминесценция
Примечания
- ↑ C. Plinius Secundus. Naturalis Historia, Liber IX, XLIII (de pisce qui noctibus lucet)
- ↑ Dubois. Note sur las physiologie des pyrophores. C. R. Seances Soc. Biol.2:559-562 (1885)
- ↑ R. Dubois. Note sur la fonction photogenique chez la Phpolas Dactilus. C. R. Seances Soc. Biol. 39:564-566 (1887)
- ↑ B. Bilter, W. D. McElroy. Preparation and properties of crystalline firefly luciferin. Arch. Biochem. Biophys. 72:358-368 (1957)
- ↑ Shimomura, Osamu; Toshio Goto, Yoshimasa Hirata (1957). «Crystalline Cypridina Luciferin». Bulletin of the Chemical Society of Japan 30 (8): 929-933. DOI:10.1246/bcsj.30.929. ISSN 0009-2673. Проверено 2010-01-06.
- ↑ Viviani, Vadim R.; Etelvino J. H. Bechara, Yoshihiro Ohmiya (1999). «Cloning, Sequence Analysis, and Expression of Active Phrixothrix Railroad-Worms Luciferases: Relationship between Bioluminescence Spectra and Primary Structures†,‡». Biochemistry 38 (26): 8271-8279. DOI:10.1021/bi9900830. Проверено 2010-01-05.
- ↑ 1 2 Ugarova, N. N.; L. G. Maloshenok, I. V. Uporov, M. I. Koksharov (2005). «Bioluminescence Spectra of Native and Mutant Firefly Luciferases as a Function of pH». Biochemistry (Moscow) 70 (11): 1262-1267. DOI:10.1007/s10541-005-0257-2. ISSN 0006-2979. Проверено 2010-01-05.
- ↑ Crystal structure of the thermostable japanese firefly luciferase (PDB id: 2d1r) complexed with oxyluciferin and AMP // PDBsum
- ↑ H Morise, O Shimomura, FH Johnson, J Winant: Intermolecular Energy Transfer in Bioluminescent systems of aequorea. Biochemistry 13 (1974) 2656-62.
Литература
- Shimomura Osamu Bioluminescence: chemical principles and methods. — World Scientific. — ISBN 9812568018, 9789812568014
Ссылки
- Bioluminescence // American Society for Photobiology
- С.Б. Ланда, Петербургский институт ядерной физики им. Б. П. Константинова РАН 188350 Гатчина, Орлова роща Коммуникация у светляков. Проверено 25 декабря 2009.
Категории:- Источники света
- Биофизика
- Физиология
- Люминесценция
- Биолюминесценция







Wikimedia Foundation. 2010.