- Дигенетические сосальщики
-

В этой статье не хватает ссылок на источники информации. Информация должна быть проверяема, иначе она может быть поставлена под сомнение и удалена.
Вы можете отредактировать эту статью, добавив ссылки на авторитетные источники.
Эта отметка установлена 10 июня 2012.Дигенетические сосальщики 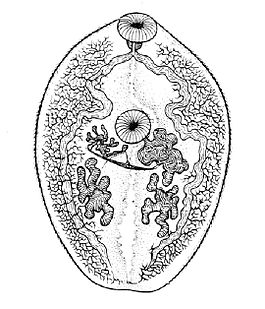
Марита Paragonimus westermaniiНаучная классификация промежуточные рангиДомен: Эукариоты Царство: Животные Подцарство: Эуметазои Без ранга: Двусторонне-симметричные Без ранга: Первичноротые Надтип: Platyzoa Тип: Плоские черви Класс: Дигенетические сосальщики Международное научное название Digenea Carus, 1863
Синонимы Отряды - Echinostomatida
- Plagiorchida
- Opistorchida
- Schistosomatida
- Strigeidida
- Azygiida
Систематика
на Викивидах
Изображения
на ВикискладеITIS 55189 NCBI 6179 Дигенетические сосальщики, или трематоды (лат. Digenea) — класс паразитических плоских червей[1]. Их жизненный цикл, за редкими исключениями, протекает в нескольких хозяевах и сопровождается закономерным чередованием не менее трёх поколений. Описано около 7200 видов. Некоторые трематоды (около 40 видов) — паразиты человека, вызывающие опасные заболевания — трематодозы, например, шистосомоз и описторхоз. К трематодам относятся: печёночная двуустка (Fasciola hepatica), кошачья двуустка (Opisthorchis felineus), шистосома (Schistosoma), лейкохлоридий парадоксальный (Leucochloridium paradoxum).
В отличие от отечественной научной традиции, в современной англоязычной научной литературе название Trematoda используют для обозначения таксона, объединяющего аспидогастров (Aspidogastrea) и собственно дигенетических сосальщиков, за которыми сохраняется название Digenea. Это противоречие носит сугубо номенклатурный характер.
Содержание
Жизненный цикл
Трематоды обладают сложным жизненным циклом с гетерогонией: закономерным чередованием амфимиктной (обычно гермафродитной) и партеногенетических стадий. Всех представителей класса характеризует наличие стадий, паразитирующих в беспозвоночных (обычно в брюхоногих моллюсках). У большинства видов в жизненном цикле также присутствует, по меньшей мере, ещё один хозяин — позвоночное.
Партеногенетические поколения
Из яйца выходит особь первого партеногенетического поколения — мирацидий, который способен плавать с помощью биения ресничек. Для дальнейшего развития мирацидий должен заразить следующего хозяина, в котором будут паразитировать все партеногенетические стадии. В роли этого хозяина практически у всех трематод выступают брюхоногие моллюски. Лишь некоторые группы в качестве хозяина используют двустворчатых моллюсков (например, трематоды сем. Bucephalidae) или многощетинковых червей (трематоды сем. Aporocotylidae). Эту стадию характеризует высокая специфичность к хозяину: как правило, мирацидии заражают и развиваются в моллюсках лишь одного вида (или нескольких близкородственных видах).
Выход мирацидия из яйца у разных видов происходит либо после заглатывания яйца хозяином (пассивное заражение), либо во внешней среде (активное заражение; подразумевает поиск хозяина). Последний вариант может реализоваться только у тех видов, яйца которых для развития должны попасть в воду. В то же время, заражение даже водных моллюсков некоторыми видами трематод осуществляется пассивно.
При обнаружении хозяина мирацидий проникает через покровы и претерпевает метаморфоз, обычно сводящийся к сбрасыванию ресничных покровов и формированию нового эпителия — тегумента. У небольшого числа видов превращение носит более кардинальный характер, и их мирацидии теряют почти все органы, а следующие стадии развиваются из небольшого числа клеток. Претерпевший метаморфоз мирацидий называют материнской спороцистой. В материнской спороцисте из партеногенетических яйцеклеток развивается следующее поколение — дочерние партениты, в зависимости от строения называемые редиями или дочерними спороцистами. Дочерние партениты в размножения отрождают либо следующие поколения дочерних партенит такого же строения, либо церкарий — личинок амфимиктного поколения.
Дочерние партениты некоторых трематод могут обитать в моллюске в течение нескольких лет, не приводя к его гибели, подавляя, однако, его репродуктивную активность. Паразитизм других видов, характеризующихся массовым выходом церкарий, закономерно приводит к гибели хозяина.
Амфимиктное поколение
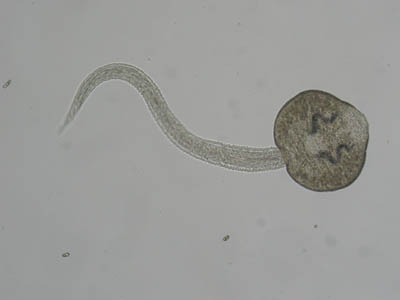
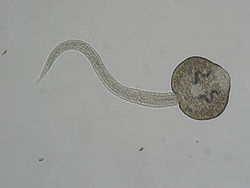 Церкария трематоды Fascioloides magna
Церкария трематоды Fascioloides magna
Поведение вышедшей из хозяина церкарии (расселительной личинки) специфично для разных видов. Выделяют три основных стратегии:
- прямое заражение окончательного хозяина через наружные покровы или при случайном заглатывании;
- проникновение и инцистирование в промежуточном хозяине (стадия метацеркарии), при этом промежуточный хозяин служит пищей для окончательного хозяина или следующего промежуточного хозяина (встраивание в трофическую цепь);
- инцистирование во внешней среде на неживых предметах, водных растениях или раковинах моллюсков (стадия адолескарии) с последующим пассивным заражением окончательного хозяина.
Существуют трематоды (например, лейкохлоридий парадоксальный), церкарии которых не выходят из тела хозяина, а заражают следующего хозяина при поедании им предыдущего.
За исключением единичных случаев, окончательный хозяин, в котором развиваются половозрелые особи амфимиктного поколения (мариты), — позвоночное. В организме хозяина они могут локализоваться в кишечнике, паренхиме и протоках печени, жёлчном пузыре, органах выделительной системы, дыхательных путях, кровеносном русле или даже под веком. Образующиеся после скрещивания марит яйца выводятся во внешнюю среду с калом или мочой хозяина.
Мариты
Внешнее строение
Размеры марит — особей гермафродитного поколения — варьируют от десятков микрон до нескольких сантиметров. В отдельных случаях длина может достигать нескольких десятков сантиметров.
Тело зачастую листовидной формы, уплощённое по дорсо-вентральной оси.
Как правило, имеются две присоски:- ротовая — на переднем конце тела; на её дне находится ротовое отверстие;
- брюшная (ацетабулюм) — расположена на брюшной стороне тела и обыкновенно смещена к переднему концу.
В теле марит выделяют два отдела: локомоторный, или преацетабулярный (перед брюшной присоской); и генеративный, или постацетабулярный (после неё), которые несколько различаются по морфологии.Однако в разных семействах существуют различные отклонения от такой общей схемы внешнего строения:
- Сем. Echinostomatidae вместо ротовой присоски имеется воротничок — складка на дорсальной стороне, частично заходящая на брюшную сторону.
- Сем. Paramphistomatidae: ротовая присоска полностью вторично редуцирована; терминально образуется передний орган (за счёт модификации самого переднего отдела пищеварительной системы). Брюшная присоска сильно смещена назад.
- Сем. Bivisiculidae: нет присосок вообще.
- Сем. Sanguinicolidae: нет брюшной присоски.
- Сем. Notocotylidae: также нет ацетабулюма, но развиваются вторичные прикрепительные образования (группы секреторных клеток).
- У высших фуркоцеркных трематод ротовая присоска очень маленькая, брюшная — почти редуцитрована; функцию прикрепления берёт на себя орган Брандеса.
- Сем. Bucephalidae: присоски отсутствуют; ротовое отверстие — на вентральной стороне, приблизительно посередине; терминально спереди — передний орган.
Кожно-мускульный мешок
Покровы трематод представлены типичным тегументом: сплошная наружная синцитиальная пластинка связана цитоплазматическими мостиками с погружёнными ядросодержащими частями — цитонами. В цитоплазме синцитиальной пластинки зачастую имеются актиновые шипики. Поверхностные структуры (такие как микровилли и микротрихии) у большинства представителей отсутствуют.
Под синцитиальной пластинкой расположены ряды кольцевых, продольных и диагональных мышц. Необходимо отметить, что хорошо выраженные мышечные слои имеются только в локомоторном отделе тела мариты; в генеративном же мышечных волокон значительно меньше и расположены они неупорядоченно. Строение мышечной ткани у трематод достаточно архаично: мышечная клетка представлена миоцитоном и отходящими от него отростками, которые содержат сократительные волокна. Каждый миоцитон имеет от двух до десяти отростков.
Паренхима у трематод фактически отсутствует. То, что ранее называли клетками паренхимы, оказалось либо цитонами тегумента, либо миоцитонами.
Нервная система
Нервная система трематод является типичным примером ортогона. Имеются три пары продольных стволов (дорсальные, латеральные и вентральные), которые связаны поперечными комиссурами (связками). Из трёх пар стволов, как правило, наиболее развиты брюшные. На переднем конце имеется трапецивидный ганглий. Вперёд от него отходит некоторое количество нервных окончаний, иннервирующих мышцы ротовой присоски и органы чувств. От вентральных стволов к брюшной присоске также отходит система нервных стволиков, образующих в её районе сплетение.
В генеративном отделе тела решётка ортогона утрачивает свою правильность, нервные стволы сильно утончаются, комиссуры между ними практически исчезают, количество нейронов резко падает.Протонефридиальная система
У марит трематод в отличие от остальных плоских червей имеется мочевой пузырь с эпителизованными стенками и своей мускулатурой. Его отверстие расположено субтерминально на заднем конце тела и снабжено сфинктером. От мочевого пузыря к переднему концу тела идут два главных собирательных канала, по ходу часто образующие узелки — сильно извитые и плотно упакованные участки. Далее главные собирательные каналы делятся на передние и задние продольные собирательные каналы. От этих каналов идут множественные ответвления, от которых в свою очередь отходят от трёх до девяти тончайших капилляров, на концах которых расположены циртоциты — мерцательные клетки.
Основной функцией протонефридиальной системы, по-видимому, является осморегуляция. Выделительная же функция у трематод осуществляется аммонотелическим путём (то есть продуктом азотистого обмена является аммиак, который выделяется во внешнюю среду через покровы).
Кроме того, у многих трематод формируется вторичная (резервная, лакунарная) выделительная система: на поздних этапах развития мариты возникают множественные анастомозы между правой и левой частями протонефридиальной системы, а также дополнительные выросты.
Функции вторичной выделительной системы — распределительная (в её сосудах обнаруживается большое количество аминокислот, простых сахаров) и выделительная (на стадии инцистировавшейся метацеркарии она работает как почка накопления).Дыхательная система
Специализированных органов дыхания нет. Паразиты используют анаэробный обмен.
Пищеварительная система
Пищеварительная система трематод замкнутая. Ротовое отверстие расположено на переднем конце тела на дне ротовой присоски (если таковая имеется). Оно ведёт в перфарингиальный отдел, после которого расположена мускулистая глотка. Далее идёт пищевод, который перед брюшной присоской разделяется на две слепозамкнутые ветви кишечника.
Кроме того, у многих крупных трематод (например, Fasciola hepatica) наблюдается дополнительное ветвление пищеварительной системы, поскольку она берёт на себя распределительную функцию.
Совершенно иначе устроена пищеварительная система у представителей семейства Bucepalidae:
- У них ротовое отверстие расположено на брюшной стороне тела. Оно ведёт в небольшую щелевидную полость, после которой идёт глотка. За глоткой следует мешковидная кишка.
- Таким образом семейство Bucepalidae относят к подклассу Gasterostomata и противопоставлют всем остальным трематодам (подкласс Prosostomata).
Половая система
Большинство марит трематод являются синхронными облигатными гермафродитами, то есть имеют и женскую, и мужскую половую систему. Однако среди трематод есть и раздельнополые: в семействах Schistosomatidae и Didymozoidae.
Женская часть половой системы
Как правило имеется один яичник (хотя он бывает вторично сегментирован). От него отходит яйцевод, в котором расположено множество различных клапанов, препятствующих прохождению обратно в яичник зрелых яйцеклеток, а также сперматозоидов. В стенке яйцевода имеется вырост — семяприемник (служит для хранения спермы). В яйцевод впадает проток желточника (желточник обычно представлен множественными фолликулами).
Также от яйцевода отходит так называемый Лауреров канал, который открывается отверстием на брюшной или спинной стороне тела. Его функция состоит в выведении из яйцевода избытков желточных клеток и сперматозоидов.После места впадения Лаурерова канала яйцевод резко расширяется и образует крупную овальную камеру — оотип. Оотип имеет сильно утолщённые стенки, к которым примыкают одноклеточные скорлуповые железы. Совокупность оотипа и скорлуповых желёз называют тельцем Мелиса.
Мужская часть половой системы
Обыкновенно в развитии закладываются два семенника, которые впоследствии могут фрагментироваться. Исключением являются представители семейства Monorchidae, имеющие один семенник.
От семенников отходят семявыносящие каналы, идущие вперёд до брюшной присоски и объединяющиеся там в семяпровод. На семяпроводе могут иметься два вздутия — семенные пузырьки. Дальше семяпровод впадает в сумку цирруса — копулятивного органа.
Мирацидий
 Строение мирацидия: 1 — хоботок; 2 — апикальная железа; 3 — латеральные железы; 4 — глазки; 5 — мозговой ганглий; 6 — протонефридий; 7 — эпителиальная пластинка; 8 — зародышевые клеткиДополнительные сведения: Спороциста
Строение мирацидия: 1 — хоботок; 2 — апикальная железа; 3 — латеральные железы; 4 — глазки; 5 — мозговой ганглий; 6 — протонефридий; 7 — эпителиальная пластинка; 8 — зародышевые клеткиДополнительные сведения: СпороцистаМирацидий является личинкой партеногенетического поколения трематод.
Условно всех мирацидиев трематод можно разделить на две группы:
- Многоклеточные — 60–80 клеток, размеры: 100–160 микрон. Такие личинки всегда имеют период активного существования во внешней среде. Активно, перкутанно заражают промежуточного хозяина. Продолжительность жизни — 12–14 часов.
- Малоклеточные — 12–18 клеток, размеры: 12–14 микрон. Мирацидии этого типа не покидают яйцевых оболочек во внешней среде. Заражение происходит, если промежуточный хозяин съедает яйцо с мирацидием: личинка выходит из оболочек яйца в пищеварительной системе хозяина и внедряется в эпителий кишки. Продолжительность жизни малоклеточных мирацидиев может достигать нескольких месяцев.
Форма тела мирацидиев обычно каплевидная. На переднем конце располагается хоботок — орган для внедрения в ткани хозяина.
Покровы
Тело мирацидиев покрыто рядами эпителиальных пластинок — это крупные клетки, несущие множество ресничек, использующихся для локомоции. Количество и расположение эпителиальных пластинок весьма стабильно и используется в качестве систематического признака.
Эпителиальные пластинки не контактируют друг с другом: между ними проходят гиподермальные гребни, образующие синцитиальную сеть в промежутках между эпителиальными пластинками. Цитоны гиподермы погружены в глубь тела, под мышечные слои. Гиподермальные гребни не связаны между собой под эпителиальными пластинками; сплошную синцитиальную пластинку они образуют только на переднем конце мирацидия, где расположен хоботок.
Мускулатура
Мускулатура тела представлена слоями продольных и кольцевых мышечных волокон. При движении во внешней среде она используется только для осуществления поворотов (само движение осуществляется исключительно за счёт ресничек).
Хоботок имеет свою специализированную мускулатуру.Протонефридиальная система
Имеются одна или две пары циртоцитов. Главные собирательные каналы открываются латерально. Мочевой пузырь отсутствует.
Пенетрационный аппарат
В передней части тела мирацидия расположена крупная четырёхъядерная железа — апикальная. Протоки этой железы открываются на вершине хоботка. По бокам от неё расположены латеральные железы, протоки которых открываются у основания хоботка.
Апикальная железа и латеральные железы в комплексе представляют собой пенетрационный аппарат — то есть служат для внедрения в ткани хозяина, поскольку их секрет обладает цитолитическим и гистолитическим действием.
Нервная система
В передней трети тела мирацидия расположен крупный мозговой ганглий, иннервирующий эпителиальные пластинки, мышечные волокна и сложный набор сенсорных структур (до 17 типов). Все поверхностные сенсиллы мирацидиев расположены только на гиподерме (на эпителиальных пластинках они отсутствуют).
Органы чувств мирацидиев можно разделить на следующие группы:
- сенсиллы, связаные с хоботком;
- парные пигментированные глазки;
- непарный непигментированный глазок;
- сенсиллы между первым и вторым рядами эпителиальных пластинок (ниже чувствительные элементы отсутствуют).
Социальная организация у трематод
В 2010 году американские исследователи показали, что партениты одного из видов рода Himasthla, паразитирующие в улитках Cerithidea californica, чётко разделяются на две группы, характеризующиеся различными размерами, строением и поведением. Более крупные редии ведут малоподвижный образ жизни и способны к размножению, более многочисленные мелкие редии, названные первооткрывателями «солдатами», не размножаются и активно атакуют как трематод других видов, так и неродственных им представителей того же вида[2].
См. также
Примечания
- ↑ БСЭ, 1969-1978.
- ↑ 1 2 Hechinger, R. F., Wood, A. C., Kuris, A. M. (2011). Social organization in a flatworm: trematode parasites form soldier and reproductive castes. Proceedings of the Royal Society B 278: 656-665. DOI:10.1098/rspb.2010.1753 (англ.)
Категории:- Животные по алфавиту
- Трематоды
- Классы животных

Wikimedia Foundation. 2010.