- Rhynchonelliformea
-
Rhynchonelliformea 
Acanthothiris
Stethothyris toorlooensis
Dagnochonetes supragibbosa
Mucrospirifer thedfordensis
Раковины ископаемых RhynchonelliformeaНаучная классификация промежуточные рангиДомен: Эукариоты Царство: Животные Подцарство: Эуметазои Без ранга: Двусторонне-симметричные Без ранга: Первичноротые Надтип: Спиральные Тип: Плеченогие Подтип: Rhynchonelliformea Международное научное название Rhynchonelliformea A. Williams, Carlson, Brunton, Holmer & Popov, 1996
Классы - † Chileata
- † Obolellata
- † Kutorginata
- Rhynchonellata
- † Strophomenata
Геохронология 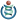
Систематика
на Викивидах
Поиск изображений
на ВикискладеRhynchonelliformea (лат.) — подтип плеченогих (брахиопод).
Для представителей данного подтипа характерна кальцитовая раковина, состоящая из брюшной и спинной створок. Эти створки, как правило, сочленяются посредством зубов, зубчиков или пластин. Кишечник оканчивается слепо, анальное отверстие отсутствует.

Rhynchonelliformea ведут неподвижный образ жизни на морском дне. Питаются они скоплениями бактериальных клеток и молекулами органических веществ, отфильтровывая их из воды.
Как правило, это раздельнополые животные. После оплодотворения яйцеклетки начинается радиальное дробление бластомеров. Гаструляция обычно осуществляется путём инвагинации, а мезодерма образуется энтероцельным способом. В процессе развития личинки бластопор закрывается. Личинка не способна к самостоятельному питанию. Прожив от нескольких часов до нескольких суток, она опускается на субстрат, прикрепляется к нему и переживает метаморфоз. Вторичный рот формируется на месте бластопора.
Представители подтипа широко распространены в морях с нормальной солёностью. Разные виды встречаются на различных глубинах, но обычно не более 300 м.
Rhynchonelliformea известны начиная с атдабанского века раннего кембрия. Они были весьма многочисленны и разнообразны в геологическом прошлом.
Содержание
История изучения
Из родов, входящих в состав данной группы и признаваемых в настоящее время, самым первым был описан род Terebratula. Его описание было опубликовано в 1776 году О. Ф. Мюллером[1].
В качестве самостоятельного подтипа Rhynchonelliformea были впервые выделены в 1996 году Альвином Уильямсом, Сандрой Дж. Карлсон, Говардом Брантоном, Ларсом Холмером и Л. Е. Поповым[2]. Эти пять авторов участвовали в написании томов по брахиоподам для нового издания фундаментального труда «Treatise on Invertebrate Paleontology» и объединили свои усилия для разработки новой классификации данной группы животных[3]. В 1996 году они опубликовали статью, в которой предлагалась новая система брахиопод, основанная на данных анатомии, эмбриологии и молекулярной биологии[2][3]. В последующие годы эта система была принята в «Treatise on Invertebrate Paleontology» и ряде других работ.
Особенности строения
Раковина
Раковина состоит из двух створок — брюшной и спинной[4]. Её размеры обычно составляют несколько сантиметров[5]. Крупнейшими раковинами, достигавшими размера около 40 см, обладали представители палеозойского рода Gigantoproductus[5]. Из современных представителей подтипа самые крупные относятся к виду Magellania venosa, длина их раковин достигает 8,4 см[5].
Край раковины, на котором находится макушка, называется задним краем, а противоположный ему — передним краем. У представителей классов Rhynchonellata и Strophomenata створки обычно сочленяются посредством замкового сочленения — двух зубов под макушкой на заднем крае брюшной створки, входящих в соответствующие им ямки на спинной створке. У некоторых форм (подотряд Productidina) замковое сочленение вторично утрачено[6]. Сочленение створок отсутствует у многих древнейших форм — у отряда Chileida, а также некоторых представителей класса Obolellata[6]. У прочих Obolellata сочленение осуществлялось зубчиками брюшной створки и ямками спинной створки[6]. У представителей отряда Dictyonellida[7] и класса Kutorginata сочленение осуществлялось посредством борозд на замочных краях створок[8][9].
Раковина сформирована из кальцита и покрыта снаружи тонким органическим слоем — периостракумом. Кальцитовая составляющая обычно двух- или трехслойная, редко состоит из одного слоя. Первичный (наружный) слой образован мелкими кристаллами кальцита, часто имеющими игольчатую форму и направленными перпендикулярно поверхности раковины. Вторичный слой имеет фиброзную (волокнистую), либо пластинчатую или перекрещенно-пластинчатую микроструктуру. Третичный слой, если он развит, состоит из многоугольных призм кальцита, направленных перпендикулярно поверхности.
Мускулатура
Имеются мускулы открыватели створок (дидукторы) и закрыватели створок (аддукторы), а также ножные мускулы.
Пищеварительная система
У современных представителей кишечный тракт оканчивается слепо, анальное отверстие отсутствует[10]. У представителей вымершего класса Kutorginata возможно имелось анальное отверстие [11].
Жизненный цикл
Размножение
Большинство современных видов раздельнополы. Встречаются, однако, и гермафродитические виды, которые в целом отличаются в целом меньшими размерами[12].
Половой диморфизм отсутствует у современных форм, за исключением Lacazella mediterranea[12].
Оплодотворение яйцеклеток происходит в мантийной полости самки или во внешней среде.
У некоторых видов (особенно часто у гермафродитических) развитие личинок происходит на внутренней поверхности мантии родительской особи, либо в пределах её лофофора. У представителей рода Lacazella для этого служат выводковые камеры, которые формируются на лофофоре, а у Argyrotheca и Gwynia capsula — выводковые камеры в полости тела[13].
Индивидуальное развитие
Об индивидуальном развитии Rhynchonelliformea судят по современным представителям этой группы, из которых эмбриология лучше всего изучена у Calloria inconspicua, Notosaria nigricans, Terebratulina septentrionalis, Terebratulina retusa и Terebratalia transversa[14].
Развитие эмбриона
 Дробление зиготы Terebratalia transversa
Дробление зиготы Terebratalia transversa
 Гаструляция Terebratalia transversa (в разрезе)
Гаструляция Terebratalia transversa (в разрезе) Формирование личинки Terebratalia transversa (вид сбоку)
Формирование личинки Terebratalia transversa (вид сбоку)После оплодотворения яйцеклетки сперматозоидом образуется зигота[14]. Обычно она покрывается оболочкой оплодотворения, препятствующей проникновению других сперматозоидов[15].
Дробление зиготы на клетки-бластомеры радиальное[16]. После четвертой фазы дробления, на которой число клеток достигает 16, дальнейшее деление часто идёт хаотично и несинхронно. В результате формируется бластула с внутренней полостью — бластоцелем[17].
Затем бластула превращается в гаструлу. Как и у прочих брахиопод, это превращение (гаструляция) обычно происходит путём инвагинации (впячивания стенки бластулы в бластоцель). Участок, на котором происходит впячивание, называется вегетативным полюсом. На месте впячивания образуется первичный рот — бластопор, впяченная часть стенки превращается в энтодерму, а остальная — в эктодерму. Известно, однако, и исключение: у особей вида Lacazella mediterranea гаструляция происходит путём деляминации (расслоения бластулы на два слоя клеток, формирующих энтодерму и эктодерму)[17].
На стадии поздней бластулы или ранней гаструлы на поверхности эмбриона развиваются реснички, и он становится способен к передвижению в толще воды[17]. На анимальном полюсе тела[18] (противоположном вегетативному, то есть бластопору) формируется апикальный пучок длинных неподвижных ресничек, предположительно выполняющий сенсорные функции[17][19]. Тогда же, возможно, происходит и прорыв оболочки оплодотворения с выходом эмбриона наружу, однако этот процесс плохо изучен[17].
В ходе дальнейшего развития гаструла приобретает вытянутую форму. При этом бластопор из округлого также становится вытянутым, а сторона тела, на которой он расположен, становится брюшной стороной. Анимальный полюс с апикальным пучком ресничек несколько смещается к одному из концов бластопора и становится передним концом тела[18].
Мезодерма образуется энтероцельным путем, то есть из энтодермы[17]. Участок энтодермы на спинной стороне утолщается и даёт начало слою мезодермальных клеток. Разрастаясь в боковых направлениях, этот слой формирует целомические полости[20]. Сообщения о формировании мезодермы у некоторых представителей схизоцельным способом (из клеток на границе эктодермы и энтодермы) не подтвердились[17].
После возникновения мезодермы бластопор постепенно исчезает, зарастая от заднего конца к переднему[20]. Его окончательное исчезновение рассматривается как превращение эмбриона в личинку[14].
Развитие личинки
Сформировавшаяся личинка — лецитотрофная, то есть она не способна питаться[21]. Тело личинки поперечными пережимами разделено на передний, туловищный и задний (ножной) отделы[20]. На переднем отделе иногда имеются парные глазки, которых у разных видов насчитывается от нескольких штук до нескольких десятков[21]. У некоторых представителей вдоль заднего края переднего отдела развиты несколько десятков гранул, называемых везикулярными тельцами. Функции этих телец неясны[19]. Туловищный отдел формирует складку, которая охватывает задний отдел. Часто на краю данной складки присутствуют 4 пучка щетинок — два боковых и два спинных[22]. Эти щетинки выполняют осязательную функцию, а также служат средством защиты[23]. У зрелых личинок реснички сохраняются в основном на переднем отделе, причём, у многих видов по периметру переднего отдела развит поясок двигательных ресничек; кроме того, реснички сохраняются на брюшной стороне туловищного отдела, где они образуют продольную полоску[19].
Плавание личинки продолжается от нескольких часов до нескольких суток, после чего личинка оседает на дно или другой подходящий субстрат. Задний отдел выделяет клейкую или слизистую субстанцию и прикрепляется к субстрату[24].
Дальнейшее развитие
Перед оседанием личинки на субстрат или сразу после этого она утрачивает апикальный пучок ресничек и глазки[25].
В процессе оседания или несколько позже личинка претерпевает метаморфоз. Края складки туловищного отдела поднимаются вверх и охватывают передний отдел, образуя мантию[26]. На наружной поверхности мантии начинает формироваться раковина[25]. Иногда формирование раковины начинается даже до метаморфоза[27].
После прикрепления личинки в субстрату её задний отдел превращается в ножку[28].
На месте бывшего бластопора образуется новое ротовое отверстие. Вокруг него из тканей переднего отдела формируется лофофор[25].
Образ жизни
Во взрослом возрасте представители Rhynchonelliformea ведут практически неподвижный образ жизни.
Экологические типы
По соотношению с субстратом представители данной группы относятся к трём экологическим типам[29].
- Якорный экологический тип — наиболее распространённый. Организм крепится к субстрату при помощи ножки, существующей на протяжении всей жизни животного. Для представителей этого экологического типа характерны выпуклые створки. Подразделяется на три экологических подтипа[29]:
- Основной подтип якорного типа. Ножка крупная, иногда разветвлённая, удерживает раковину над субстратом без помощи опорных образований.
- Опорный подтип. На взрослых стадиях ножка не способна удерживать раковину над субстратом и дополнительной опорой служит макушка.
- Усложнённый подтип. Функцию дополнительной опоры выполняет высокая арея — участок раковины вдоль задних краев створок.
- Свободнолежащий экологический тип. Организм свободно лежит на субстрате, не прикрепляясь к нему. Отверстие в раковине для выхода ножки зарастает у взрослых особей[29].
- Первый подтип включает формы с сильно выпуклыми створками и массивной макушечной частью.
- Второй подтип включает формы, лежащие на дне на широкой уплощенной или вогнутой брюшной створке.
- Третий (понтонный) подтип включает формы, которые удерживаются на поверхности мягкого грунта с помощью отростков раковины, имеющих форму игл или пластин нарастания.
- Прирастающий экологический тип. Организм прикрепляется к субстрату путём цементации поверхностью брюшной створки или её частью. Прикрепление ножкой отсутствует. Характерны асимметричные брюшные и уплощённые спинные створки[29].
Питание
Пищей представителям Rhynchonelliformea служат в основном агрегаты бактериальных клеток и молекулы органических веществ. Большинство из них не может питаться фитопланктоном, в частности диатомовыми водорослями, которыми питаются прочие плеченогие[30].
Питание осуществляется путём фильтрации. Створки раковины в процессе питания приоткрыты. Сомкнутые друг с другом щупальца лофофора делят мантийную полость на две части: в одну из этих частей вода поступает из внешней среды, затем проходит сквозь лофофор и профильтрованная выходит из второй части мантийной полости[31]. Входящий и исходящий ток воды создаются колебательными движениями ресничек на щупальцах лофофора[32].
Распространение и среда обитания
Представители подтипа широко распространены в морях с нормальной солёностью (не менее 26-30‰). Большинство современных видов приурочено к глубинам менее 300 метров, но отдельные виды встречаются и на глубине 6 километров.
В ископаемом состоянии известны начиная с атдабанского века раннего кембрия, весьма многочисленны и разнообразны в палеозойских отложениях, однако к мезозою это разнообразие заметно сократилось и до настоящего времени дожили представители лишь трёх отрядов.
Классификация
А. Уильямс и его соавторы, выделившие подтип Rhynchonelliformea, включили в его состав группы, которые ранее объединялись под названием замковых брахиопод (Articulata). Замковые брахиоподы были разделены на три самостоятельных класса: Rhynchonellata, Strophomenata и Chileata.
Кроме того, в состав Rhynchonelliformea в качестве самостоятельного класса Obolellata[3] были включены некоторые формы, которые прежде включались в состав беззамковых брахиопод (Inarticulata)[33].
Ещё один класс Kutorginata, также включённый в состав данного подтипа[3], ранее рассматривался в качестве отряда Kutorginida среди беззамковых[34] или как группа брахиопод неясного систематического положения[35].
Таким образом, в составе подтипа выделяют пять классов, из которых четыре являются полностью вымершими:
- † Класс Chileata
- † Класс Obolellata
- † Класс Kutorginata
- _ Класс Rhynchonellata
- † Класс Strophomenata
Примечания
- ↑ Williams, 1965, p. H2
- ↑ 1 2 Williams et al., 1996
- ↑ 1 2 3 4 Williams et al., 2000
- ↑ Лихарев, 1960, с. 116
- ↑ 1 2 3 Лихарев, 1960, с. 121
- ↑ 1 2 3 Williams, Brunton & McKinnon, 1997, p. 364
- ↑ Holmer, 2000, p. 198
- ↑ Williams, Brunton & McKinnon, 1997, p. 364—365
- ↑ Popov & Williams, 2000, p. 208—209
- ↑ Смирнова, 1990, с. 16
- ↑ Popov & Williams, 2000, p. 210
- ↑ 1 2 Williams et al., 1997, pp. 125—126
- ↑ Williams et al., 1997, pp. 176—178
- ↑ 1 2 3 Williams et al., 1997, p. 153
- ↑ Williams et al., 1997, p. 156
- ↑ Смирнова, 1990, с. 9
- ↑ 1 2 3 4 5 6 7 Williams et al., 1997, p. 163
- ↑ 1 2 Freeman, 2003, p. 272
- ↑ 1 2 3 Pennington, Stricker, 2001, p. 443
- ↑ 1 2 3 Williams et al., 1997, p. 165
- ↑ 1 2 Williams et al., 1997, p. 173
- ↑ Williams et al., 1997, p. 174
- ↑ Williams et al., 1997, p. 176
- ↑ Williams et al., 1997, p. 179—181
- ↑ 1 2 3 Williams et al., 1997, p. 186
- ↑ Williams et al., 1997, p. 182
- ↑ Williams et al., 1997, p. 183
- ↑ Смирнова, 1990, с. 11
- ↑ 1 2 3 4 Смирнова, 1990, с. 30—33
- ↑ Смирнова, 1990, с. 34
- ↑ Абрикосов, 1987, с. 417
- ↑ Абрикосов, 1987, с. 416
- ↑ Rowell, 1965b, p. 291H
- ↑ Смирнова, 1990, с. 42
- ↑ Rowell, 1965a, p. 296H
Литература
- Абрикосов Г. Г. Класс Плеченогие (Brachiopoda) // Жизнь животных. — Москва: Просвещение, 1987. — Т. 1. — С. 415—419. — 300 000 экз.
- Лихарев Б. К. Общая часть // Основы палеонтологии / Главный редактор Ю. А. Орлов. Ответственный редактор тома Т. Г. Сарычева. — Москва: Издательство Академии наук СССР, 1960. — Т. Мшанки, брахиоподы. Приложение — форониды. — С. 115—162. — 344 с. — 3200 экз.
- Смирнова Т. Н. Брахиоподы. — Москва: Издательство Московского университета, 1990. — 72 с. — 500 экз. — ISBN 5-211-02389-7
- Gary Freeman. Regional specification during embryogenesis in Rhynchonelliform brachiopods (англ.) // Developmental Biology : Журнал. — 2003. — Vol. 261. — № 1. — С. 268–287. — DOI:10.1016/S0012-1606(03)00307-5
- Lars E. Holmer Dictyonellida // Treatise on Invertebrate Paleontology. Part H, Brachiopoda / Roger L. Kaesler, Editor. — Revised. — Kansas: The Geological Society of America and the University of Kansas, 2000. — Vol. 2: Linguliformea, Craniiformea, and Rhynchonelliformea (part). — P. 196—200. — ISBN 0–8137–3108–9
- J. T. Pennington & S. A. Stricker. Phylum Brachiopoda // Atlas of Marine Invertebrate Larval Forms / C. M. Young, Editor. — New York: Academic Press, 2001. — P. 441—461. — ISBN 0127731415
- Leonid E. Popov & Alwyn Williams Kutorginata // Treatise on Invertebrate Paleontology. Part H, Brachiopoda / Roger L. Kaesler, Editor. — Revised. — Kansas: The Geological Society of America and the University of Kansas, 2000. — Vol. 2: Linguliformea, Craniiformea, and Rhynchonelliformea (part). — P. 208—215. — ISBN 0–8137–3108–9
- A. J. Rowell. Class Uncertain // Treatise on Invertebrate Paleontology. Part H, Brachiopoda / Raymond C. Moore, Editor. — The Geological Society of America and the University of Kansas, 1965. — Vol. 1. — P. H296—H297. — ISBN 0-8137-3008-2
- A. J. Rowell. Inarticulata // Treatise on Invertebrate Paleontology. Part H, Brachiopoda / Raymond C. Moore, Editor. — The Geological Society of America and the University of Kansas, 1965. — Vol. 1. — P. H260—H296. — ISBN 0-8137-3008-2
- Alwyn Williams. Introduction // Treatise on Invertebrate Paleontology. Part H, Brachiopoda / Raymond C. Moore, Editor. — The Geological Society of America and the University of Kansas, 1965. — Vol. 1. — P. H1—H5. — ISBN 0-8137-3008-2
- Alwyn Williams, C. Howard C. Brunton & David I. McKinnon. Morphology // Treatise on Invertebrate Paleontology. Part H, Brachiopoda / Roger L. Kaesler, Editor. — Revised. — Kansas: The Geological Society of America and the University of Kansas, 1997. — Vol. 1: Introduction. — P. 321—422. — xx + 539 p. — ISBN 0-8137-3108-9
- Alwyn Williams, S. J. Carlson, C. H. C. Brunton, L. E. Holmer & L. E. Popov. A supra-ordinal classification of the Brachiopoda (англ.) // Philosophical Transactions of the Royal Society of London (series B). — 1996. — Vol. 351. — P. 1171—1193. — DOI:10.1098/rstb.1996.0101
- Alwyn Williams, M. A. James, C. C. Emig, Sarah Mackay & M. C. Rhodes. Anatomy // Treatise on Invertebrate Paleontology. Part H, Brachiopoda / Roger L. Kaesler, Editor. — Revised. — Kansas: The Geological Society of America and the University of Kansas, 1997. — Vol. 1: Introduction. — P. 7—188. — xx + 539 p. — ISBN 0-8137-3108-9
- Alwyn Williams, Sandra J. Carlson & C. Howard C. Brunton. Brachiopod Classification // Treatise on Invertebrate Paleontology. Part H, Brachiopoda / Roger L. Kaesler, Editor. — Revised. — Kansas: The Geological Society of America and the University of Kansas, 2000. — Vol. 2: Linguliformea, Craniiformea, and Rhynchonelliformea (part). — P. 1—21. — ISBN 0–8137–3108–9
Тип Плеченогие Домен Эукариоты · Царство Животные · Подцарство Эуметазои · Двусторонне-симметричные · Первичноротые · Надтип Спиральные Подтип Linguliformea Класс Lingulata Отряды: Лингулиды · † Акротретиды · † Сифонотретиды Класс † Paterinata Отряд: † Патериниды Подтип Craniiformea Класс Craniata Отряды: Кранииды · † Craniopsida · † Trimerellida Подтип Rhynchonelliformea Класс † Chileata Отряды: † Chileida · † Диктионеллиды Класс † Obolellata Отряды: † Оболеллиды · † Наукатиды Класс † Kutorginata Отряд: † Куторгиниды Класс Rhynchonellata Отряды: † Протортиды · † Ортиды · † Пентамериды · Ринхонеллиды · † Атрипиды · † Атиридиды · † Спирифериды · † Спирифериниды · Тецидеиды · Теребратулиды
Надсемейство: † Cadomelloidea
Семейства: † Cardiarinidae · † Mongolellidae · † Tropidoleptidae
Род: † DentatrypaКласс † Strophomenata Отряды: † Строфомениды · † Продуктиды · † Ортотетиды · † Биллингселлиды
Семейство: (?) † LiosomeniidaeУстаревшие таксоны: Беззамковые · Замковые · Calciata Группы неясного систематического положения, сходные с плеченогими:
Семейства † Mickwitziidae · † TianzhushanellidaeКатегории:- Животные по алфавиту
- Rhynchonelliformea




Wikimedia Foundation. 2010.