- Криптофитовые водоросли
-
Криптофитовые водоросли 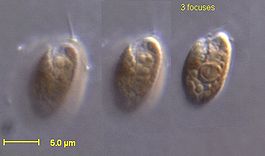
Rhodomonas salinaНаучная классификация Международное научное название Cryptophyceae Pascher, 1913
Систематика
на Викивидах
Изображения
на ВикискладеNCBI 3027 EOL 3354 Криптофитовые водоросли (криптомонады, криптофиты, лат. Cryptophyta) — тип хромистов, включающий класс Cryptophyceae и два порядка — Cryptomonadales и Goniomonadales.
Криптофиты — небольшая, но экологически и эволюционно очень важная группа организмов, обитающих в морских и континентальных водах. Почти все они — одноклеточные подвижные жгутиконосцы, некоторые представители способны формировать пальмеллоидную стадию. Только один род Bjornbergiella (выделен из почв Гавайских островов) способен к образованию простых нитчатых талломов (положение этого рода до конца не выяснено, и в ряде систем его не относят к криптофитам), оспаривается и существование колониальных форм.
Содержание
Клетки
У криптофит клетки обычно уплощённые, имеют выпуклую спинную и прямую или вогнутую брюшную стороны. Форма клеток разнообразная: яйцевидная, эллипсоидная, грушевидная, бобовидная, веретеновидная и др. Передний конец клетки более или менее скошен, от него отходит продольная борозда, которая не доходит до заднего конца. Наличие борозды и её длина — признаки, которые используются для систематики на уровне рода. Глотка мешковидная, по её краю располагаются трихоцисты. По периферии клетки также могут располагаться трихоцисты, но более мелких размеров. Трихоцисты (эжектосомы) окружены мембраной и внутри содержат 2 ленты, скрученные рулоном (цилиндры). Более крупный цилиндр состоит из намотанного по спирали лентовидного мембранного материала. Вверху он образует V-образное углубление, в котором находится второй цилиндр. Эти две части, по-видимому, связаны между собой с помощью волокон. При химическом и физическом раздражении туго скрученные ленты распрямляются, разрывая мембрану эжектосомы и перипласт. Сначала выбрасывается маленький цилиндр и, покидая клетку, он вытягивает за собой ленту большого цилиндра. Образуются эжектосомы в пузырьках аппарата Гольджи. Вначале цилиндры содержат несколько оборотов, но по мере роста увеличивают число оборотов.
Криптомонады очень чувствительны к изменению окружающих условий, таких, как изменение рН, осмотических условий или температуры. Они в массе отстреливают содержимое эжектосом, что приводит к быстрому разрушению клетки. С этим явлением связана значительная трудность по переводу криптомонад на лабораторные среды.
На переднем конце клетки расположены сократительные вакуоли, изливающие своё содержимое в глотку или борозду.
В клетках криптомонад имеются два тельца Mona, функции которых неизвестны. Так как тельца Mona содержат много мембран и фибрилл, они, возможно, вовлечены в процесс деструкции клеточных органелл, особенно отработанных трихоцист.
Клеточные покровы
У криптофит клеточные покровы представлены перипластом, который состоит из плазмалеммы и дополнительного белкового материала, расположенного над плазмалеммой. Внутриклеточный компонент перипласта может иметь форму чехла, как у Chilomonas, или состоять из пластинок (шестиугольных, как у Cryptomonas, или прямоугольных, как у Chroomonas). Белковые пластинки прикрепляются к плазмалемме с помощью округлых или многоугольных внутримембранных частиц. Наружный компонент перипласта может состоять из пластинок, чешуек, слизи или их комбинации. Перипласт у криптомонад отсутствует в области глотки.
Жгутиковый аппарат
На переднем конце клеток расположены два жгутика, длина которых сопоставима с длиной клетки. При движении они или оба направлены вперёд, или один — вперёд, а другой — назад. На жгутиках имеются особые криптофитовые мастигонемы, которые могут располагаться в два супротивных ряда на длинном жгутике (длина мастигонем до 2 мкм), в один ряд на коротком (длина мастигонем 1 мкм) либо на каждом жгутике. Мастигонемы криптомонад двухчастные, состоят из трубчатой части и тонкого терминального волоска. У некоторых видов короткий жгут свободен от мастигонем. На длинном жгутике при основании может находиться дополнительный пучок, состоящий приблизительно из 45 волосков. На жгутиках могут располагаться небольшие органические чешуйки в виде семиугольных розеток (140—170 нм в диаметре).
Переходная зона жгутиков содержит две или более пластинчатые структуры (септы) ниже точки, где заканчиваются две центральные микротрубочки. Корешковая система представлена ризостилем (состоит из 6—10 микротрубочек, соединённых сократительными фибриллами), который идёт в глубь клетки, трёх микротрубочковых корешков и фибриллярного корешка, прилегающего к одному из микротрубочковых. В разных родах имеются различия в строении корешкового аппарата жгутиков.
Глазок (стигма) у криптомонад связан с хлоропластом, но не связан со жгутиковым аппаратом, поэтому может располагаться и в центре клетки. Он состоит из большого числа пигментных глобул, расположенных параллельными рядами. Для некоторых криптомонад показан положительный фототаксис. Стигма встречается не у всех представителей.
Хлоропласты
В клетках криптомонад расположены один или два хлоропласта, реже больше, окрашенные в разные цвета — от сине-зелёного и оливково-зелёного до жёлто-бурого, коричневого и тёмно-красного. Иногда хлоропласты отсутствуют.
В хлоропластах присутствует ряд пигментов. Помимо хлорофилла а имеется хлорофилл с2, который передаёт энергию света от фикобилинов к хлорофиллу а. У криптомонад отсутствует аллофикоцианин, выполняющий эту функцию у красных и синезелёных водорослей. Фикобилипротеиновые пигменты криптомонад криптофикоциан и криптофикоэритрин отличаются организацией полипептидных компонентов и по спектральным характеристикам от фикобилипротеиновых пигментоы синезелёных и красных водорослей. Они локализованы не в фикобилисомах, а во внутритилакоидных пространствах, из-за этого тилакоиды криптомонад толще, чем у других водорослей. Каждая фотосинтезирующая криптомонада имеет только один тип фикобилипротеина — или фикоэритрин, или фикоцианин, но никогда вместе.
Криптомонады образуют несколько уникальных ксантофиллов, таких, как аллоксантин, который может быть использован для их обнаружения в смеси планктонных водорослей. У них встречаются α- и β-каротины, крококсантин, зеаксантин и монадоксантин.
В хлоропластах могут быть пиреноиды, в которые могут заходить ламеллы (каждая из 1—2 тилакоидов).
Оболочка хлоропласта состоит из четырёх мембран, наружная на своей поверхности несёт рибосомы. Между двумя парами мембран в оболочке хлоропласта криптомонад находится перипластидное пространство, в котором располагаются 80S рибосомы с эукариотным типом рРНК, зёрна крахмала и сильно редуцированное ядро, известное как нуклеоморфа. Нуклеоморфа окружена двухмембранной оболочкой с порами и содержит ядрышко. Размер ДНК нуклеоморфы составляет только 660 тыс. пар нуклеотидов. Она имеет всего три хромосомы (240, 225 и 195 тыс. пар нкулеотидов), которые кодируют процессы, необходимые только для функционирования нуклеоморфы. Нуклеоморфа делится во время деления ядра вслед за репликацией базального тела, но раньше, чем хлоропласт. Деление амитотическое, и хотя в её геноме обнаружены гены α-, β- и γ-тубулинов, веретено деления не образуется из микротрубочек, но имеются фибриллярные структуры, появляющиеся только во время деления. Дочерние нуклеоморфы мигрируют в противоположные концы пластиды, таким образом после деления пластиды каждая дочерняя пластида получает свою нуклеоморфу.
Наличие нуклеоморфы, перипластидного пространства и строение оболочек хлоропласта свидетельствуют о вторичном симбиотическом происхождении хлоропластов криптофитовых, когда бесцветная фаготрофная флагеллата поглотила («съела») эукариотную красную водоросль, которая затем трансформировалась в пластиду. Тогда две внутренние мембраны хлоропласта криптофитовых интерпретируют как мембраны хлоропласта красной водоросли, третью мембрану — как плазмалемму эндосимбионта (красной водоросли), наружную, четвёртую, как мембрану пищеварительной вакуоли клетки-хозяина. Симбиотическое происхождение хлоропласта криптомонад от красных водорослей подтверждается филогенетическим анализом, основанным на сравнении нуклеотидных последовательностей гена 16S рРНК нуклеоморфы и ядер красных водорослей.
Запасные продукты
Основное запасаемое вещество у криптофит — крахмал, откладывается между второй и третьей мембранами оболочки хлоропласта. Криптомонады — единственная группа водорослей с таким местом откладывания крахмала. Как и у красных водорослей, он содержит больше амилопектина (разветвлённые молекулы с α-1,4- и α-1,6-связями), чем амилозы (линейные цепочки с α-1,4-связями). С йодом такой крахмал даёт красное окрашивание. В цитоплазме могут присутствовать капли липидов.
Особенности питания
Среди криптомонад встречаются автотрофы, гетеротрофы (сапротрофы и фаготрофы) и миксотрофы. Большинство из них требуют витамин В12 и тиамин, некоторые нуждаются в биотине. Криптомонады могут использовать аммоний и органические источники азота, но морские представители менее способны к превращениям нитратов в нитриты по сравнению с другими водорослями. Органические вещества стимулируют рост криптомонад. Для различных бесцветных представителей и для окрашенного Cryptomonas ovata показана фаготрофность. Окрашенный в сине-зелёный цвет Chroomonas pochmanni обладает миксотрофным питанием. Для захвата и удержания бактериальных клеток образуются специализированные вакуоли. Бактерии втягиваются в вакуоль через маленькую пору в глотке, где отсутствуют перипластные пластинки. Перевариваются они в небольших вакуолях.
Ядро
Единственное ядро расположено в задней части клетки, или, реже, посередине. Митоз начинается после деления хлоропласта и имеет ряд особенностей: он открытый, ядрышко не исчезает, центриоли, по-видимому, отсутствуют. Область полюса веретена уплощена и ограничена цистернами эндоплазматической сети.
Число хромосом у криптомонад очень большое — от 40 до 210. В метафазе хромосомы объединяются в массивную пластинку. В ней образуются каналы, свободные от хроматина, через которые проходят микротрубочки веретена.
Размножение
Основной способ размножения криптомонад — вегетативный, за счёт деления клетки пополам с помощью борозды деления. При этом впячивание плазмалеммы идёт начиная с заднего конца клетки. Чаще всего делящаяся клетка сохраняет подвижность. Максимальная скорость роста для многих криптомонад — одно деление в день при температуре около 20 °С. Дефицит азота и избыточное освещение стимулируют образование покоящихся стадий. Покоящиеся цисты окружены толстым экстрацеллюлярным матриксом.
Для Ch. acuta известен хологамный половой процесс, при котором вегетативные клетки функционируют как гаметы. Слияние клеток происходит на брюшной стороне, причём задний конец одной клетки прикрепляется к середине другой. Когда клетки полностью сольются, формируется четырёхжгутиковая зигота. Затем она делится; первое деление, предположительно, мейотическое.
У криптомонады Proteomonas sulcata в определённой степени гетероморфный гаплодиплонтный жизненный цикл, в котором и диплоидная, и гаплоидная стадии жгутиковые. Эти стадии отличаются друг от друга по размерам клеток, структуре перипласта, конфигурации жгутикового аппарата. До сих пор в этом жизненном цикле для этого вида не известны оплодотворение и мейоз.
Экология и значение
Криптофитовые — типичные представители планктона, изредка они встречаются в иле солёных озёр и среди детрита в пресных водоёмах. Они занимают видное положение в олиготрофных, умеренных и высокоширотных, пресных и морских водах.
Большинство криптомонад встречается в широком температурном диапазоне, но некоторые виды Rhodomonas относятся к стенотермным. Среди стенотермных представителей криофильные (холодолюбивые) виды Cryptomonas способны вегетировать на поверхности льда и снега. Криптофиты играют важную роль в холодных водах, особенно зимой и ранней весной. Например, они могут доминировать в весеннем цветении фитопланктона в Северном море, где, возможно, являются важным звеном в пищевых цепях. Локализованные цветения, вызванные развитием криптомонад, встречаются в антарктических водах. Цветения воды коррелируют с притоком воды от таяния ледников. В покрытых льдами озёрах Антарктиды виды криптомонад могут доминировать в альгофлоре, давая более 70 % биомассы фитопланктона. Очень редко криптомонады встречаются в океанических водах при температуре 22 °С и выше. Они отсутствуют в горячих источниках и гиперсолёных водоёмах.
Пресноводные представители предпочитают искусственные и естественные водоёмы со стоячей водой — отстойники, различные пруды (биологические, технические, рыбоводные), реже встречаются в водохранилищах и озёрах. Они образуют крупные популяции в озёрах на глубине 15—23 м, в месте соединения слоёв воды, богатых и бедных кислородом, где уровень света значительно ниже, чем у поверхности. Бесцветные представители распространены в водах, загрязнённых органическими веществами, их много в сточной воде, поэтому они могут служить показателем загрязнения воды органическими веществами.
Среди криптофит встречаются типичные сфагнофиллы, живущие в болотах с низким значением рН, в то время как ряд видов может развиваться в широком диапазоне рН (например, Chilomonas встречается при рН 4,1—8,4).
Криптофиты играют значительную роль в водоёмах в круговороте кислорода, углерода, азота и фосфора, в синтезе органического вещества из минеральных веществ. Ими питаются различные представители зоопланктона, которыми в свою очередь питаются рыбы. Сообщалось об интенсивном поедании криптомонад кладоцерами в эвтрофных прудах в зимнее время года. В то же время отмечались случаи цветения воды в прудах, вызванных Cyanomonos, которые приводили к массовой гибели сомов.
Криптомонады обитают в качестве эндосимбионтов в миксотрофных инфузориях Myrionecta rubra (прежние названия — Mesodinium rubrum или Cyclotytrichum meunieri). Эта инфузория может образовывать нетоксичное красное цветение воды на побережье Перу, Калифорнии, Камчатки и др. (в основном в зоне апвеллинга). У неё отсутствует клеточный рот (у других представителей этого рода инфузорий, не имеющих эндосимбионтов, на переднем конце клетки имеется клеточный рот), и в своём питании она зависит от эндосимбионта, который на свету способен ассимилировать углерод и фосфор. Каждая инфузория может содержать несколько криптомонад, которые отделены от цитоплазмы хозяина двойной мембраной. Наружная мембрана рассматривается как мембрана пищеварительной вакуоли хозяина, а внутренняя — как плазмалемма симбионта. Симбионты содержат ядро, митохондрии, пластиды с нуклеоморфой и некоторые другие структуры. Хлоропласты имеют все характерные особенности хлоропластов криптофитов. В то же время у эндосимбионта отсутствует перипласт, жгутиковый аппарат, трихоцисты.
Некоторые динофиты (например, Dinophysis) приобрели хлоропласты в результате симбиоза с криптомонадами. В одних случаях у них может сохраняться ядро криптомонады, в других — только пластиды.
Эволюция и филогения
В эволюции криптофит исходными считаются формы, близкие к роду Goniomonas. Эти бесцветные представители способны к фагоцитозу, у них отсутствует нуклеоморфа и пластиды. Полагают, что пластиды криптомонад появились в результате вторичного симбиоза с эукариотом из Rhodophyta, что подтверждается филогенетическим анализом (сравнение пластидного генома криптомонад и красных водорослей, анализ генов нуклеоморфы, кодирующих α- и β-тубулины). Эволюция внутри криптофит выглядит следующим образом. Наличие борозды без глотки у Goniomonas — наиболее примитивная особенность. Глотка формировалась за счёт смыкания краёв борозды, начиная с заднего конца. Полное исчезновение борозды, но наличие глотки представлено у Chroomonas. В процессе эволюции криптомонад происходил повторный переход к гетеротрофности, что привело к образованию рода Chilomonas, у которого редуцирована борозда, развита глотка и имеются лейкопласты. Эта схема подтверждается эволюцией жгутикового аппарата и анализом нуклеотидных последовательностей гена 18S рРНК.
Положение криптомонад среди водорослей и их объём до сих пор дискутируются. А. Пашер полагал, что они произошли от хризомонад, и от криптомонад выводил перидинеи. Он объединил криптомонады и перидинеи в отдел Pirrophyta. Большинство альгологов не соглашаются с таким объдинением и исключают криптофитовых из пиррофитовых, рассматривая их как самостоятельную группу на основании того, что криптофиты отличаются от динофит строением ядра, жгутикового аппарата, наличием нуклеоморфы, составом пигментов, биохимическими особенностями. Все эти данные свидетельствуют против их близкого филогенетического родства.
Разнообразие
К настоящему времени описано более 200 видов криптофитовых (приблизительно 100 пресноводных и 100 морских). В последние годы в связи с использованием электронного микроскопа значительные изменения претерпела их систематика. Были переописаны и описаны новые таксоны, однако недостаточная конкретность и неоднозначность использованных признаков затрудняет не только определение родовой, но и видовой принадлежности этих организмов.
Систематика
- порядок Cryptomonadales
- род Campylomonas
- род Chilomonas
- род Chroomonas
- род Cryptomonas
- род Falcomonas
- род Geminigera
- род Guillardia
- род Hemiselmis
- род Plagioselmis
- род Proteomonas
- род Rhodomonas
- род Storeatula
- род Teleaulax
- порядок Goniomonadales
- род Goniomonas
Примечания


Для улучшения этой статьи желательно?: - Найти и оформить в виде сносок ссылки на авторитетные источники, подтверждающие написанное.
- Исправить статью согласно стилистическим правилам Википедии.
- Викифицировать статью.
Категории:- Хромисты по алфавиту
- Хромисты
- Типы хромистов
- порядок Cryptomonadales

Wikimedia Foundation. 2010.