- Эвглениды
-
Эвглениды 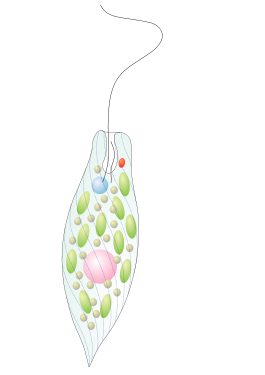
Эвглена зелёнаяНаучная классификация Международное научное название Euglenida
Систематика
на Викивидах
Изображения
на ВикискладеITIS 43804 EOL 4842 Эвглениды (лат. Euglenida) — группа протистов. Объединяет около 1000 видов, среди них имеется много бесцветных форм.
Общая характеристика
Форма тела
У эвгленовых форма тела варьирует от веретеновидной, овальной до плосколистовидной и игловидной. Передний конец тела более или менее закруглён, задний может быть вытянутым и заканчиваться заострённым отростком. Клетки могут быть спирально скручены. Длина клеток от 5 до 500 мкм и более.
Жгутиковый аппарат
Эвглениды имеют 1, 2, 3, 4 и 7 видимых жгутиков, за исключением небольшой группы безжгутиковых форм, а также прикрепленных организмов. Жгутики отходят от колбообразного впячивания на переднем конце клетки — глотки (ампулы). Глотка состоит из продолжительного канала и внутреннего расширения — резервуара. Чаще всего у эвгленовых два неравных по длине жгутика: один направлен вперёд (передний, спинной, дорсальный), другой — назад (задний, брюшной, вентральный). У некоторых родов, например Euglena, короткий жгутик не выходит за пределы глотки. На поперечном разрезе жгутика виден параксиальный тяж, состоящий из белка. Из-за наличия этого тяжа жгутики эвгленовых толще, поэтому они заметнее, чем жгутики других эукариотных водорослей. Параксиальный тяж хорошо развит у длинного жгутика, но может быть слабо выраженным или отсутствовать у короткого. У некоторых видов он отсутствует и в длинном жгутике. Жгутики эвгленовых и кинетопластид покрыты рядом тонких волосков, которые отличаются по своему строению от тубулярных (трубчатых) волосков — мастигонем — охрофитов. Волоски на жгутиках эвгленовых расположены группами по 3-4 в один ряд, который тянется по спирали по всей длине жгутика. Помимо этого на жгутиках могут быть короткие волоски в виде покрытия (войлока). Полагают, что функция волосков у эвглен — увеличение гидродинамической устойчивости жгутиков.
Жгутики эвгленовых имеют типичную для эукариот аксонему. Особенность жгутикового аппарата эвгленовых — просто устроенная переходная зона. В длинной переходной зоне часто отсутствует поперечная пластинка. Только у Entosiphon в ней имеется мощная псевдозвёздчатая структура. Базальные тела («переднее» и «заднее») располагаются параллельно друг другу и имеют типичное для эукариот строение (9 триплетов микротрубочек).
У некоторых эвгленовых изучено расположение микротрубочковых корешков жгутиков. Имеют место три асимметрично расположенных таких корешков. Два из них — «промежуточный» и «передний» — связаны с базальным телом переднего жгутика, третий корешок — с базальным телом заднего жгутика. Корешки направлены к верхнему концу клетки вдоль глотки и, как полагают, играют важную роль в придании определённой формы клетке, а также принимают участие в фагоцитозе. Микротрубочки заднего и промежуточного корешков связаны с лентами микротрубочек, поддерживающих канал и пелликулу. Обычно имеется исчерченное соединение между двумя базальными телами жгутиков, функция которого — координирование движения жгутиков. У ряда эвгленовых присутствуют фибриллярные корешки. Так, поперечно исчерченный корешок ризопласт имеется у Peranema, а простые фибриллярные корешки у Distigma.
Светочувствительный аппарат
Светочувствительная система эвгленовых состоит из двух структур. Первый компонент — это парафлагелларное тело (парабазальное вздутие), представляет собой вздутие при основании одного видимого жгутика и содержит синие светочувствительные флавины. Это вздутие выполняет функцию фоторецептора. Мутантные клетки эвгленовых без жгутиков не обладают фототаксисом, даже при наличии глазка. Второй компонент системы — глазок (стигма), расположенный в цитоплазме около резервуара напротив парафлагелларного тела. Глазок состоит из 50-60 глобул, образующих один слой и окружённых мембраной. Глобулы содержат каротиноиды астаксантин и/или эхиненон, которые придают глазку оранжево-красную окраску. При движении эвгленовых глазок затенят парабазальное вздутие. При положительном фототаксисе клетка ориентирована таким образом, чтобы парабазальное вздутие затенялось минимально.
Клеточная оболочка
У эвгленовых оболочка клетки называется пелликулой (по терминологии зоологов — кутикула). Она состоит из цитоплазматической мембраны, расположенных под ней белковых полос, системы микротрубочек и трубчатых цистерн эндоплазматической сети. По химическому составу пелликула на 70—80 % состоит из белков, а также содержит липиды и углеводы. Пелликулярные белковые полосы (состоят преимущественно из артикулинов), у большинства эвгленовых идущие по спирали, реже продольно, располагаются в цитоплазме клетки непосредственно под цитоплазматической мембраной, своими краями находя друг на друга. В местах соединения одна полоса по краю формирует выпуклость, а соседняя вогнутость. Кроме того, в таком «замке» расположены четыре микротрубочки, по две с каждой стороны, образуя как бы рельс, в который вставляется край соседней полосы, обеспечивая скользящее движение одной полосы относительно другой. Пелликулярные полосы исчезают в месте перехода канала в резервуар глотки, но остаются микротрубочки, которые вместе с микротрубочками, отходящими от переднего корешка, выстилают резервуар, образуя его цитоскелет.
Если пелликулярные полосы эластичны и подвижны одна относительно другой, то такие клетки способны к изменению своей формы — метаболии и могут осуществлять движение, используя или не используя жгутик. К такому эвгленоидному движению способны, например, представители рода Astasia, некоторые Euglena и др. Эвгленовые водоросли, имеющие толстую пелликулу и, как правило, продольные пелликулярные полосы, не способны к эвгленоидному движению. Такая жёсткая пелликула характерна для родов Lepocinclis, Phacus и некоторых Euglena.
С каждой пелликулярной полосой связана цистерна эндоплазматической сети, которая, по-видимому, функционирует как резервуар кальция.
Ниже пелликулы располагаются выделительные органеллы — слизистые тельца и мукоцисты. Большинство таксонов эвгленовых содержат слизистые тельца, имеющие вид мешков, заполненных аморфными или фибриллярными водорастворимыми мукополисахаридами. Они находятся под местами соединения пелликулярных полос и выделяют слизь наружу через свои узкие каналы, расположенные между пелликулярными полосами и открывающимися порами. Слизистые тельца принимают участие в формировании ножки у Colacium, образования домиков у Strombomonas и Trachelomonas, образовании цист, а также в эвгленоидном движении. У таких таксонов, как Peranema и Entosiphon, имеются мукоцисты — крупные цилиндричесике структуры, содержащие решётчатую конструкцию из углеводов, которые сохраняют свою удлинённую форму при выделении через пору.
Эвгленовые водоросли могут формировать цисты и пальмеллоидные стадии, связанные с образованием слоистого слизистого чехла вокруг клетки, препятствующего её движению. Слизистый секрет для таких пальмеллоидных стадий освобождается аппаратом Гольджи первоначально в глотку, из которой уже распределяется по поверхности всей клетки.
У некоторых эвгленовых, таких, как Ascoglena, Trachelomonas и Strombomonas образуются домики, в которых монады помещаются свободно. Домики имеют на переднем конце отверстие, через которое выходит жгутик, а при размножении выползает одна из дочерних особей. Домики выделяются протопластом в виде слизистого слоя, в котором откладываются соли марганца и железа. Цвет домика может варьировать от бесцветного до тёмно-коричневого даже в пределах одной популяции одного вида. Это зависит от накопления окислов железа и марганца. Поэтому цвет домика зависит от экологических условий: содержания этих элементов в воде, концентрации кислорода (при высокой концентрации кислорода железо и марганец находятся в виде слабо растворимых соединений, при низкой концентрации — эти минералы растворимы).
Ядро
Клетки эвгленовых имеют одно крупное ядро. Хромосомы при окрашивании хорошо видны даже в интерфазе, так как находятся, как и у динофитов, в конденсированном состоянии. Митоз закрытый, веретено деления внутреннее, центриоли отсутствуют. Хромосомные микротрубочки присоединяются к кинетохорам. Оформленные центры организации микротрубочек внутри ядра отсутствуют. Концы микротрубочек обычно не связаны с ядерной мембраной. Перед митозом ядро мигрирует вверх клетки к резервуару. Часто, но не всегда, пара базальных тел (которые удваиваются перед ядерным делением) формирует полюса веретена. Веретено развивается под прямым углом к продольной оси клетки. Ядрышко не исчезает во время митоза, в метафазе становится сильно удлинённым. Хромосомы формируют неправильную метафазную пластинку. Во время телофазы ядро приобретает форму гантели, становясь сильно удлинённым в анафазе. Оболочки дочерних ядер образуются путём перешнуровки оболочки материнского ядра.
Цитокинез происходит вслед за делением ядра путём впячивания мембраны по продольной оси клетки.
Хлоропласт
Форма хлоропласта у разных видов эвгленовых широко варьирует: звёздчатая, пластинчатая, лентовидная, дисковидная, с пиреноидом или без. Обычно в клетке содержится несколько хлоропластов. Эвглены, растущие при высокой температуре, УФ-облучении или на среде со стрептомицином и другими антибиотиками, теряют свои пластиды. Такие клетки не способны синтезировать пластиды de novo и остаются бесцветными.
В хлоропластах эвгленовых тилакоиды собраны в ламеллы по три. Иногда их может быть больше, но граны, как у растений, не образуются. Хлоропласт покрыт тремя мембранами. Толщина такой оболочки составляет 35—45 нм. Полагают, что хлоропласт эвгленовых — результат вторичного эндосимбиоза бесцветного эукариотного эвгленового предка с зелёной эукариотной водорослью. В этом случае две его внутренние мембраны интерпретируются как мембраны хлоропласта зелёной водоросли, а третья, наружная, — как мембрана пищеварительной вакуоли клетки-хозяина.
Запасной продукт
У большинства эвгленовых запасным продуктом является парамилон (β-1-3-связанный глюкан); некоторые запасают лейкозин и липиды. Помимо эвгленовых парамилон обнаружен у примнезиофитов из рода Pavlova. При гидролизе парамилон даёт глюкозу; от йода он не синеет; образуется в цитоплазме. Встречается в цитоплазме даже у неокрашенных представителей. Парамилоновые зёрна бывают различной формы: шаровидные, эллипсоидные, кольцевидные, палочковидные. У одних видов зёрна парамилона мелкие, многочисленные, у других — крупные (парамилии), от одного до нескольких на клетку. Положение парамилий в клетке строго локализовано. Считают, что помимо энергетической функции парамилон имеет определённое отношение к перемещению клеток, так как у клеток, заполненных парамилоном, затрудняется метаболия и замедляется движение.
Сократительная (пульсирующая) вакуоль
Расположена на переднем конце клетки и связана с осморегуляторной функцией. Заполненная сократительная вакуоль имеет сферическую функцию и окружена рядом мелких добавочных вакуолей. Она изливает своё содержимое в резервуар.
Митохондрии
В клетках эвгленовых митохондрии могут сливаться, образуя сеть. Для них характерно наличие дисковидных крист. У эвгленид, обитающих в анаэробных условиях, митохондрии отсутствуют.
Биосинтез лизина
У эвгленовых, в отличие от всех других фотосинтезирующих организмов, лизин синтезируется через α-аминоадипиновую кислоту, как у настоящих грибов и животных.
Жизненный цикл
Эвгленовые размножаются делением клетки пополам. Борозда деления проходит от переднего конца клетки к заднему. При стрессе у эвгленовых могут формироваться толстостенные цисты. Их образование связано с потерей жгутиков, увеличением числа парамилоновых гранул, вздутием и округлением клетки, увеличением числа слизистых телец и образованием слоистой слизистой стенки, состоящей из полисахаридов. Образование цист стимулируется низким уровнем питательных веществ и низким соотношением азот/фосфор в среде обитания.
Хотя для Scytomonas сообщают о половом цикле с хологамным половым процессом, у подавляющего большинства эвгленовых половое размножение не известно. Отсутствие полового процесса у них, как полагают, может быть связано с их ранним отделением от общей линии эукариот ещё до появления полового размножения. По другой точке зрения, эвгленовые могли произойти от предков с половым размножением, но в дальнейшем его потеряли.
Экология и значение
Эвгленовые водоросли обитают в основном в пресных водах, предпочитая водоёмы с замедленным стоком и богатым содержанием органических веществ. Их можно обнаружить в прибрежье озёр и рек, в мелких водоёмах, включая лужи, на рисовых полях, на сырой почве. В почвах бесцветные представители встречаются на глубине 8-25 см. Окрашенные эвгленовые могут вызывать цветение воды, образуя на её поверхности плёнку зелёного или красного цвета.
В значительной степени эвгленовые водоросли реагируют на степень минерализации воды: чем она выше, тем беднее их качественный и количественный состав. Некоторые выдерживают высокую солёность воды. В прибрежных водах Норвегии они могут формировать крупные популяции после весеннего цветения, вызванного диатомовыми водорослями, а у северного побережья Ирландии в летние месяцы концентрация клеток Eutreptiella достигает 1 млн на литр.
Эвгленовые могут развиваться при широкой амплитуде колебаний pH: 4,5-8,8, однако, E. mutabilis, например, может обитать при pH 3 и даже ниже. Эвгленовые способны развиваться в условиях значительного колебания температур (от 0 до 30 °C), но максимум их развития наблюдается летом и осенью. Среди них встречаются холодолюбивые виды, которые могут вмерзать в лёд, не теряя жизнеспособности.
Среди эвгленовых встречаются фотоавтотрофы, гетеротрофы (фаготрофы и сапротрофы) и миксотрофы. Только треть родов способны к фотосинтезу, а остальные — фаготрофы и осмотрофы. Даже фотосинтезирующие эвгленовые способны к гетеротрофному росту. Большинство гетеротрофных форм — сапротрофы, поглощающие растворённые в воде питательные вещества. Такие осмотрофы, как, например, Astasia, Hyalophacus произошли от фотосинтезирующих предков, потеряв пластиды. Distigma и Rhabdomonas отделились от общего дерева эвгленофит до момента отделения фототрофов от фаготрофных Peranema. Среди фаготрофов встречаются бактериотрофы (например, Petalomonas, Ploeotila) и эукариотрофы (например, Dinema, Peranema). Фаготрофы, например, такие, как Peranema и Entosiphon, имеют специальный аппарат для захвата и переваривания пищевых частиц.
Большинство эвгленовых — свободно живущие представители, в то же время виды рода Colacium — эпибионты на беспозвоночных животных или эпифиты на нитчатых водорослях. Некоторые эвгленовые паразитируют в кишечнике коловраток, нематод, олигохет, плоских червей, амфибий, на жабрах рыб.
Эвгленовые — существенный компонент биоценозов различных типов водоёмов. Они активно участвуют в круговороте железа, органических кислот. Ими питаются личинки некоторых беспозвоночных.
Миксотрофия обеспечивает их участие в процессах самоочищения природных вод и очистке сточных вод. В санитарной гидробиологии некоторые виды используют как биологические индикаторы для проведения биологического анализа состояния органического загрязнения пресных вод.
Благодаря наличию различных способов питания, эвгленовых используют в качестве модельных объектов для разработки общебиологических проблем, вопросов биохимии, физиологии и цитологии. В лабораториях их часто используют как хорошие объекты для изучения фотосинтеза, тонкого строения хлоропластов, фототаксиса, движения жгутиков и т. д. В медицинской практике особенно популярна E. gracilis, используемая, в частности, для выяснения действия антибиотиков. Она применяется также для выяснения действия гербицидов, ростовых веществ, для количественного определения в среде витамина B12, к недостатку которого E. gracilis очень чувствительна.
Филогения
Эвгленовые — хорошо очерченная группа. В настоящее время доминирует точка зрения о первичности в эволюции эвгленовых гетеротрофных форм. Полагают, что фотосинтезирующие эвгленовые произошли от двужгутиковых фототрофных форм, каждый жгутик которых имел параксиальный тяж, базальные тела жгутиков были соединены исчерченным тяжом и от них отходили три асимметрично расположенных микротрубочковых корешка. Эта точка зрения подтверждается данными молекулярной биологии, обнаружением у ряда фототрофных эвгленовых скрытого цитостома (клеточного рта), особенностями строения жгутикового аппарата. Молекулярная филогения показывает, что от фототрофных эвгленовых за счёт потери пластид произошли некоторые осмотрофные представители и что фототрофы с двумя видимыми жгутиками предшествовали фототрофам с одним видимым жгутиком.
Наличие двух мембран у хлоропластов эвгленовых указывает на то, что хлоропласты эвгленовых были приобретены в результате вторичного симбиоза с зелёной водорослью.
Неудачными оказались попытки сблизить эвгленовых с какими-либо другими группами водорослей. От других групп они резко отличались строением жгутикового аппарата, организацией хлоропласта и пигментных систем, дисковидными кристами митохондрий, клеточными покровами, продуктами ассимиляции и другими признаками. Изолированное положение эвгленовых среди других водорослей не вызывает в настоящее время никаких сомнений. В то же время лучшими кандидатами на роль родственников эвгленовых оказались кинетопластиды. С этой группой их сближают строение крист митохондрий, структура жгутиков, похожее деление ядра и ряд других признаков (например, митохондрии представителей рода Petalomonas, расположенные в области базальных тел жгутиков, в свободном от крист пространстве несут включения, близкие к кинетопластам). К числу отличий следует отнести разный способ синтеза лизина, отсутствие кинетопласта. Данные по последовательности 16S рРНК подтверждают наличие филогенетической близости эвгленовых и кинетопластид при их значительной удалённости от других эукариот. Полагают, что эвгленовые и кинетопластиды произошли от одного предка, к которому у кинетопластид наиболее близок род Bodo, а у эвгленовых — Petalomonas.
Разнообразие и систематика
Благодаря работам русского ученого Л. С. Ценковского, обнаружившего у эвгленовых пальмеллевидное состояние, в XIX веке они были отнесены к растениям.
В большинстве ботанических классификаций эвгленовых относят к единственному классу — Euglenophyceae, который включает около 50 родов и 800—1000 видов.
T.Cavalier-Smith (1993) придерживается другой точки зрения и выделяет три класса: Petalomonadea (нефотосинтезирующие фаготрофные бактериотрофы, с постоянной формой тела и отсутствием специальных аппаратов для захватывания пищи); Aphagea (нефаготрофные эвгленовые, как фотосинтезирующие, так и лишённые пластид); Peranemea (фаготрофные представители со специальным аппаратом для захватывания и поглощения пищи, метаболия отсутствует или в разной степени имеется). Различные системы отличаются и количеством, и объёмом порядков (от трёх до семи).
На Западе общепринятой остаётся классификация Гордона Лидейла (1967), хотя современные авторы (напр., R.E. Triemer) указывают на ее несостоятельность. По Лидейлу (1967), отдел Euglenophyta содержит только один класс Euglenophyceae и шесть порядков.
Современное положение в системе
Основные роды
- Euglena Ehrenberg, 1830 — Эвглена
- Astasia Dujardin, 1841 — Астазия
- Colacium Ehrenberg, 1838 — Колациум
- Phacus Dujardin, 1841 — Факус
- Monomorphina Mereschkowsky, 1877 — Мономорфина
- Lepocinclis Perty, 1852 — Лепоцинклис
- Cryptoglena Ehrenberg, 1835 — Криптоглена
- Trachelomonas Ehrenberg, 1833 — Трахеломонас
- Strombomonas Deflandre, 1930 — Стромбомонас
Примечания
Категории:- Протозоа по алфавиту
- Классификация протистов
- Эвгленовые

Wikimedia Foundation. 2010.