- ПОПУЛЯЦИОННАЯ ГЕНЕТИКА
раздел генетики, изучающий генофонд популяций и его изменение в пространстве и во времени. Разберемся подробнее в этом определении. Особи не живут поодиночке, а образуют более или менее устойчивые группировки, сообща осваивая среду обитания. Такие группировки, если они самовоспроизводятся в поколениях, а не поддерживаются только за счет пришлых особей, называют популяциями. Например, стадо семги, нерестящейся в одной реке, образует популяцию, потому что потомки каждой рыбы из года в год, как правило, возвращаются в ту же реку, на те же нерестилища. У сельскохозяйственных животных популяцией принято считать породу: все особи в ней единого происхождения, т.е. имеют общих предков, содержатся в сходных условиях и поддерживаются единой селекционной и племенной работой. У аборигенных народов популяция - это члены связанных родством стойбищ. При наличии миграций границы популяций размыты и потому неопределимы. Например, все население Европы - потомки кроманьонцев, заселивших наш континент десятки тысяч лет назад. Изоляция между древними племенами, усиливавшаяся с развитием у каждого из них собственного языка и культуры, вела к различиям между ними. Но обособленность их относительна. Постоянные войны и захваты территории, а в последнее время - гигантская миграция вели и ведут к определенному генетическому сближению народов. Приведенные примеры показывают, что под словом "популяция" следует понимать группировку особей, связанных территориальной, исторической и репродуктивной общностью. Особи каждой популяции отличаются друг от друга, и каждая из них в чем-то уникальна. Многие из этих различий наследственные, или генетические, - они определяются генами и передаются от родителей к детям. Совокупность генов у особей данной популяции называют ее генофондом. Для того чтобы решать проблемы экологии, демографии, эволюции и селекции, важно знать особенности генофонда, а именно: сколь велико генетическое разнообразие в каждой популяции, каковы генетические различия между географически разделенными популяциями одного вида и между различными видами, как генофонд изменяется под действием окружающей среды, как он преобразуется в ходе эволюции, как распространяются наследственные заболевания, насколько эффективно используется генофонд культурных растений и домашних животных. Изучением этих вопросов и занимается популяционная генетика.
ОСНОВНЫЕ ПОНЯТИЯ ПОПУЛЯЦИОННОЙ ГЕНЕТИКИ
Частоты генотипов и аллелей. Важнейшим понятием популяционной генетики является частота генотипа - доля особей в популяции, имеющих данный генотип. Рассмотрим аутосомный ген, имеющий k аллелей, A1, A2, ..., Ak. Пусть популяция состоит из N особей, часть которых имеет аллели Ai Aj. Обозначим число этих особей Nij. Тогда частота этого генотипа (Pij) определяется как Pij = Nij/N. Пусть, например, ген имеет три аллеля: A1, A2 и A3 - и пусть популяция состоит из 10000 особей, среди которых имеются 500, 1000 и 2000 гомозигот A1A1, A2A2 и A3A3, а гетерозигот A1A2, A1A3 и A2A3 - 1000, 2500 и 3000 соответственно. Тогда частота гомозигот A1A1 равна P11 = 500/10000 = 0,05, или 5%. Таким образом мы получаем следующие наблюдаемые частоты гомо- и гетерозигот:
P11 = 0,05, P22 = 0,10, P33 = 0,20, P12 = 0,10, P13 = 0,25, P23 = 0,30.
Еще одним важным понятием популяционной генетики является частота аллеля - его доля среди имеющих аллелей. Обозначим частоту аллеля Ai как pi. Поскольку у гетерозиготной особи аллели разные, частота аллеля равна сумме частоты гомозиготных и половине частот гетерозиготных по этому аллелю особей. Это выражается следующей формулой: pi = Pii + 0,5*еjPij. В приведенном примере частота первого аллеля равна p1 = P11 + 0,5*(P12 + P13) = 0,225. Соответственно, p2 = 0,300, p3 = 0,475.
Соотношения Харди - Вайнберга. При исследовании генетической динамики популяций, в качестве теоретической, "нулевой" точки отсчета принимают популяцию со случайным скрещиванием, имеющую бесконечную численность и изолированную от притока мигрантов; полагают также, что темпы мутирования генов пренебрежимо малы и отбор отсутствует. Математически доказывается, что в такой популяции частоты аллелей аутосомного гена одинаковы для самок и самцов и не меняются из поколения в поколение, а частоты гомо- и гетерозигот выражаются через частоты аллелей следующим образом: Pii = pi2, Pij = 2pi pj. Это называется соотношениями, или законом, Харди - Вайнберга - по имени английского математика Г.Харди и немецкого медика и статистика В. Вайнберга, одновременно и независимо открывших их: первый - теоретически, второй - из данных по наследованию признаков у человека. Реальные популяции могут значительно отличаться от идеальной, описываемой уравнениями Харди - Вайнберга. Поэтому наблюдаемые частоты генотипов отклоняются от теоретических величин, вычисляемых по соотношениям Харди - Вайнберга. Так, в рассмотренном выше примере теоретические частоты генотипов отличаются от наблюдаемых и составляют P11 = 0,0506, P22 = 0,0900, P33 = 0,2256, P12 = 0,1350, P13 = 0,2138, P23 = 0,2850. Подобные отклонения можно частично объяснить т.н. ошибкой выборки; ведь в действительности в эксперименте изучают не всю популяцию, а лишь отдельных особей, т.е. выборку. Но главная причина отклонения частот генотипов - несомненно, те процессы, что протекают в популяциях и влияют на их генетическую структуру. Опишем их последовательно. ПОПУЛЯЦИОННО-ГЕНЕТИЧЕСКИЕ ПРОЦЕССЫ
Дрейф генов. Под дрейфом генов понимают случайные изменения генных частот, вызванные конечной численностью популяции. Чтобы понять, как возникает генный дрейф, рассмотрим вначале популяцию минимально возможной численности N = 2: один самец и одна самка. Пусть в исходном поколении самка имеет генотип A1A2, а самец - A3A4. Таким образом, в начальном (нулевом) поколении частоты аллелей A1, A2, A3 и A4 равны 0,25 каждая. Особи следующего поколения могут равновероятно иметь один из следующих генотипов: A1A3, A1A4, A2A3 и A2A4. Допустим, что самка будет иметь генотип A1A3, а самец - A2A3. Тогда в первом поколении аллель A4 теряется, аллели A1 и A2 сохраняют те же частоты, что и в исходном поколении - 0,25 и 0,25, а аллель A3 увеличивает частоту до 0,5. Во втором поколении самка и самец тоже могут иметь любые комбинации родительских аллелей, например A1A2 и A1A2. В этом случае окажется, что аллель A3, несмотря на большую частоту, исчез из популяции, а аллели A1 и A2 увеличили свою частоту (p1 = 0,5, p2 = 0,5). Колебания их частот в конце концов приведут к тому, что в популяции останется либо аллель A1, либо аллель A2; иными словами и самец и самка будут гомозиготны по одному и тому же аллелю: A1 или A2. Ситуация могла сложиться и так, что в популяции остался бы аллель A3 или A4, но в рассмотренном случае этого не произошло. Описанный нами процесс дрейфа генов имеет место в любой популяции конечной численности, с той лишь разницей, что события развиваются с гораздо меньшей скоростью, чем при численности в две особи. Генный дрейф имеет два важных последствия. Во-первых, каждая популяция теряет генетическую изменчивость со скоростью, обратно пропорциональной ее численности. Со временем какие-то аллели становятся редкими, а затем и вовсе исчезают. В конце концов, в популяции остается один-единственный аллель из имевшихся, какой именно - это дело случая. Во-вторых, если популяция разделяется на две или большее число новых независимых популяций, то дрейф генов ведет к нарастанию различий между ними: в одних популяциях остаются одни аллели, а в других - другие. Процессы, которые противодействуют потере изменчивости и генетическому расхождению популяций, - это мутации и миграции.
Мутации. При образовании гамет происходят случайные события - мутации, когда родительский аллель, скажем A1, превращается в другой аллель (A2, A3 или любой иной), имевшийся или не имевшийся ранее в популяции. Например, если бы в нуклеотидной последовательности "...TЦT ТГГ...", кодирующей участок полипептидной цепи "...серин-триптофан...", третий нуклеотид, Т, в результате мутации передался ребенку как Ц, то в соответствующем участке аминокислотной цепи белка, синтезирующегося в организме ребенка, вместо серина был бы расположен аланин, поскольку его кодирует триплет TЦЦ (см. НАСЛЕДСТВЕННОСТЬ). Регулярно возникающие мутации и образовали в длинном ряду поколений всех обитающих на Земле видов то гигантское генетическое разнообразие, которое мы сейчас наблюдаем. Вероятность, с которой происходит мутация, называется частотой, или темпом, мутирования. Темпы мутирования разных генов варьируют от 10-4 до 10-7 на поколение. На первый взгляд, эти величины кажутся незначительными. Однако следует учесть, что, во-первых, геном содержит много генов, а, во-вторых, что популяция может иметь значительную численность. Поэтому часть гамет всегда несет мутантные аллели, и практически в каждом поколении появляется одна или больше особей с мутациями. Их судьба зависит от того, насколько сильно эти мутации влияют на приспособленность и плодовитость. Мутационный процесс ведет к увеличению генетической изменчивости популяций, противодействуя эффекту дрейфа генов.
Миграции. Популяции одного вида не изолированы друг от друга: всегда есть обмен особями - миграции. Мигрирующие особи, оставляя потомство, передают следующим поколениям аллели, которых в этой популяции могло вовсе не быть или они были редки; так формируется поток генов из одной популяции в другую. Миграции, как и мутации, ведут к увеличению генетического разнообразия. Кроме того, поток генов, связывающий популяции, приводит к их генетическому сходству.
Системы скрещивания. В популяционной генетике скрещивание называют случайным, если генотипы особей не влияют на образование брачных пар. Например, по группам крови скрещивание может рассматриваться как случайное. Однако окраска, размеры, поведение могут сильно влиять на выбор полового партнера. Если предпочтение оказывается особям сходного фенотипа (т.е. со сходными индивидуальными характеристиками), то такое положительное ассортативное скрещивание ведет к увеличению в популяции доли особей с родительским генотипом. Если при подборе брачной пары предпочтение имеют особи противоположного фенотипа (отрицательное ассортативное скрещивание), то в генотипе потомства будут представлены новые сочетания аллелей; соответственно в популяции появятся особи либо промежуточного фенотипа, либо фенотипа, резко отличающегося от фенотипа родителей. Во многих регионах мира высока частота близкородственных браков (например, между двоюродными и троюродными родственниками). Образование брачных пар на основе родства называют инбридингом. Инбридинг увеличивает долю гомозиготных особей в популяции, поскольку в этом случае высока вероятность того, что родители имеют сходные аллели. С повышением числа гомозигот возрастает и количество больных рецессивными наследственными болезнями. Но инбридинг способствует также большей концентрации определенных генов, что может обеспечить лучшую адаптацию данной популяции.
Отбор. Различия в плодовитости, выживаемости, половой активности и т.п. приводят к тому, что одни особи оставляют больше половозрелых потомков, чем другие - с иным набором генов. Различный вклад особей с разными генотипами в воспроизводство популяции называют отбором. Изменения нуклеотидов могут влиять, а могут и не влиять на продукт гена - полипептидную цепь и образуемый ею белок. Например, аминокислота серин кодируется шестью разными триплетами - ТЦА, ТЦГ, ТЦТ, TЦЦ, АГТ и АГЦ. Поэтому мутация может превратить один из этих триплетов в другой, но при этом не изменить самой аминокислоты. Напротив, аминокислота триптофан кодируется только одним триплетом - ТГГ, и потому любая мутация заменит триптофан на другую аминокислоту, например на аргинин (ЦГГ) или серин (ТЦГ), или даже приведет к обрыву синтезируемой полипептидной цепи, если в результате мутации появится т.н. стоп-кодон (ТГА или ТАГ). Различия между вариантами (или формами) белка могут быть незаметны для организма, но могут и существенно влиять на его жизнедеятельность. Например, известно, что когда в 6-й позиции бета-цепи гемоглобина человека вместо глутаминовой кислоты стоит другая аминокислота, а именно валин, это приводит к тяжелой патологии - серповидноклеточной анемии. Изменения в других участках молекулы гемоглобина приводят к иным формам патологии, называемым гемоглобинопатиями. Следует иметь в виду, что варианты белков не всегда можно трактовать как лучшие или худшие. Например, дефектный гемоглобин в серповидных эритроцитах не разрушается паразитом - малярийным плазмодием, поэтому больные серповидноклеточной анемией не заболевают малярией даже там, где велика смертность от нее у лиц с нормальным гемоглобином. Наличие той или иной группы крови системы AB0 не сказывается на жизнедеятельности человека, но может обеспечить иммунную защиту организма от определенных заболеваний, а в редких случаях обусловливает несовместимость плода и матери. Для ряда белков как животных, так и растений описан такой феномен: один вариант молекулы устойчив к высокой температуре, а другой хорошо работает в условиях холода. Такие примеры наглядно показывают, как индивидуальные различия в ДНК приводят к различиям в наследственной приспособленности особей к разным условиям среды. Еще большие различия в приспособленности наблюдаются по генам, определяющим размеры, физиологические признаки и поведение особей; таких генов может быть много. Отбор, как правило, затрагивает их все и может вести к формированию ассоциаций аллелей разных генов.
Генетические параметры популяции. При описании популяций или их сравнении между собой используют целый ряд генетических характеристик.
Полиморфизм. Популяция называется полиморфной по данному локусу, если в ней встречается два или большее число аллелей. Если локус представлен единственным аллелем, говорят о мономорфизме. Исследуя много локусов, можно определить среди них долю полиморфных, т.е. оценить степень полиморфизма, которая является показателем генетического разнообразия популяции.
Гетерозиготность. Важной генетической характеристикой популяции является гетерозиготность - частота гетерозиготных особей в популяции. Она отражает также генетическое разнообразие.
Коэффициент инбридинга. С помощью этого коэффициента оценивают распространенность близкородственных скрещиваний в популяции.
Ассоциация генов. Частоты аллелей разных генов могут зависеть друг от друга, что характеризуется коэффициентами ассоциации.
Генетические расстояния. Разные популяции отличаются друг от друга по частоте аллелей. Для количественной оценки этих различий предложены показатели, называемые генетическими расстояниями. Различные популяционно-генетические процессы по-разному влияют на эти параметры: инбридинг приводит к уменьшению доли гетерозиготных особей; мутации и миграции увеличивают, а дрейф уменьшает генетическое разнообразие популяций; отбор изменяет частоты генов и генотипов; генный дрейф увеличивает, а миграции уменьшают генетические расстояния и т.д. Зная эти закономерности, можно количественно исследовать генетическую структуру популяций и прогнозировать ее возможные изменения. Этому способствует солидная теоретическая база популяционной генетики - популяционно-генетические процессы математически формализованы и описаны уравнениями динамики. Для проверки различных гипотез о генетических процессах в популяциях разработаны статистические модели и критерии. Прилагая эти подходы и методы к исследованию популяций человека, животных, растений и микроорганизмов, можно решить многие проблемы эволюции, экологии, медицины, селекции и др. Рассмотрим несколько примеров, демонстрирующих связь популяционной генетики с другими науками.
ПОПУЛЯЦИОННАЯ ГЕНЕТИКА И ЭВОЛЮЦИЯ
Нередко думают, что основная заслуга Чарлза Дарвина в том, что он открыл явление биологической эволюции. Однако это совсем не так. Еще до издания его книги Происхождение видов (1859) биологи сходились во мнении, что старые виды порождают новые. Разногласия имелись лишь в понимании того, как именно это могло происходить. Наиболее популярной была гипотеза Жана Батиста Ламарка, согласно которой в течение жизни каждый организм изменяется в направлении, соответствующем среде, в которой он живет, и эти полезные изменения ("благоприобретенные" признаки) передаются потомкам. При всей своей привлекательности эта гипотеза не прошла проверку генетическими экспериментами. Напротив, эволюционная теория, разработанная Дарвином, утверждала, что 1) особи одного и того же вида отличаются друг от друга по многим признакам; 2) эти различия могут обеспечить приспособление к разным условиям среды; 3) эти различия наследственны. В терминах популяционной генетики данные положения можно сформулировать так: больший вклад в следующие поколения дают те особи, которые имеют наиболее подходящие для данной среды генотипы. Изменись среда, и начнется отбор генов, более соответствующих новым условиям. Таким образом, из теории Дарвина следует, что эволюционируют генофонды. Эволюцию можно определить как необратимое изменение генофондов популяций во времени. Совершается она путем накопления мутационных изменений ДНК, возникновения новых генов, хромосомных преобразований и др. Важную роль при этом играет то, что гены обладают способностью удваиваться (дуплицироваться), а их копии - встраиваться в хромосомы. В качестве примера вновь обратимся к гемоглобину. Известно, что гены альфа- и бета-цепи произошли путем дупликации некоего предкового гена, который, в свою очередь, произошел от предка гена, кодирующего белок миоглобин - переносчик кислорода в мышцах. Эволюционно это привело к возникновению гемоглобина - молекулы с тетрамерной структурой, состоящей из четырех полипептидных цепей: двух альфа- и двух бета-. После того как природа "нашла" тетрамерную структуру гемоглобина (у позвоночных), остальные типы структур для транспорта кислорода оказались практически неконкурентоспособными. Затем уже в течение десятков миллионов лет возникали и отбирались лучшие варианты гемоглобина (свои - в каждой эволюционной ветви животных), но в рамках тетрамерной структуры. Сегодняшний отбор по этому признаку у человека стал консервативным: он "охраняет" единственный прошедший миллионы поколений вариант гемоглобина, и любая замена в любой из цепей этой молекулы приводит к болезни. Однако многие виды позвоночных имеют два или более равноценных вариантов гемоглобина - отбор "поощрял" их одинаково. И у человека есть белки, по которым эволюция "оставила" несколько вариантов. Популяционная генетика позволяет оценить время, когда произошли те или иные события в эволюционной истории. Вновь вернемся к примеру с гемоглобином. Пусть, например, желательно оценить время, когда произошло разделение предковых генов альфа- и бета-цепей и, следовательно, возникла такая система дыхания. Мы анализируем структуру этих полипептидных цепей у человека или какого-либо животного и, сравнивая их, определяем, насколько отличаются друг от друга соответствующие нуклеотидные последовательности. Поскольку в начале своей эволюционной истории обе предковые цепи были идентичными, то, зная скорость замены одного нуклеотида на другой и число различий в сравниваемых цепях, можно узнать время от момента их дупликации. Таким образом, здесь белки выступают в качестве своеобразных "молекулярных часов". Другой пример. Сравнивая гемоглобин или другие белки у человека и приматов, можно оценить, сколько миллионов лет назад существовал наш общий с ними предок. В настоящее время в качестве молекулярных часов используют "безмолвные", не кодирующие белки участки ДНК, менее подверженные внешним воздействиям. Популяционная генетика позволяет заглянуть в глубь веков и проливает свет на такие события в эволюционной истории человечества, которые невозможно было бы выяснить по современным археологическим находкам. Так, совсем недавно, сравнивая генофонды людей из различных частей света, большинство ученых сошлись на том, что общий предок всех рас современного человека возник примерно 150 тысяч лет назад в Африке, откуда он и расселился по всем континентам через Переднюю Азию. Более того, сопоставляя ДНК людей в разных регионах Земли, можно оценить время, когда популяции человека стали расти в численности. Исследования показывают, что это произошло нескольких десятков тысяч лет назад. Таким образом, в изучении истории человечества популяционно-генетические данные начинают играть столь же важную роль, как и данные археологии, демографии и лингвистики.
ПОПУЛЯЦИОННАЯ ГЕНЕТИКА И ЭКОЛОГИЯ
Обитающие в каждом регионе виды животных, растений и микроорганизмов образуют целостную систему, известную как экосистема. Каждый вид представлен в ней своей, уникальной популяцией. Оценить экологическое благополучие данной территории или акватории позволяют данные, характеризующие генофонд ее экосистемы, т.е. генофонд слагающих ее популяций. Именно он обеспечивает существование экосистемы в данных условиях. Поэтому за изменениями в экологической обстановке региона можно проследить, изучая генофонды популяций обитающих там видов. Осваивая новые территории, прокладывая нефте- и газопроводы, следует заботиться о сохранении и восстановлении природных популяций. Популяционная генетика уже предложила свои меры, например выделение природных генетических резерватов. Они должны быть достаточно обширными, чтобы содержать основной генофонд растений и животных данного региона. Теоретический аппарат популяционный генетики позволяет определить ту минимальную численность, которая необходима для поддержания генетического состава популяции, чтобы в ней не было т.н. инбридинговой депрессии, чтобы она содержала основные генотипы, присущие данной популяции, и могла воспроизводить эти генотипы. При этом каждый регион должен иметь свои собственные природные генетические резерваты. Нельзя восстанавливать загубленные сосняки Севера Западной Сибири, завозя семена сосны из Алтая, Европы или Дальнего Востока: через десятки лет может оказаться, что "чужаки" генетически плохо приспособлены к местным условиям. Вот почему экологически грамотное промышленное освоение территории должно обязательно включать популяционные исследования региональных экосистем, позволяющие выявить их генетическое своеобразие. Сказанное относится не только к растениям, но и к животным. Генофонд той или иной популяции рыб эволюционно приспособлен именно к тем условиям, в которых он обитал в течение многих поколений. Поэтому интродукция рыб из одного природного водоема в другой порой приводит к непредсказуемым последствиям. Например, попытки развести сахалинскую горбушу в Каспии оказались безуспешными, ее генофонд оказался не в состоянии "освоить" новое местообитание. Та же горбуша, интродуцированная в Белое море, покинула его и ушла в Норвегию, образовав там временные стада "русского лосося". Не надо думать, что основными объектами заботы о природе должны быть только экономически ценные виды растений и животных, такие, как древесные породы, пушные звери или промысловые рыбы. Травянистые растения и мхи, мелкие млекопитающие и насекомые - их популяции и их генофонды наравне со всеми другими обеспечивают нормальную жизнь территории. То же относится к микроорганизмам - тысячи их видов населяют почву. Изучение почвенных микробов - задача не только микробиологов, но и популяционных генетиков. Изменение генофонда популяций при грубых вмешательствах в природу выявляется не сразу. Могут пройти десятилетия, прежде чем станут очевидными последствия в виде исчезновения одних популяций, а за ними - других, связанных с первыми.
ПОПУЛЯЦИОННАЯ ГЕНЕТИКА И МЕДИЦИНА
Один из насущнейших вопросов человечества - как лечить наследственные болезни. Однако до недавнего времени сама постановка такого вопроса казалась фантастической. Речь могла идти только о профилактике наследственных заболеваний в форме медико-генетического консультирования. Опытный врач-генетик, изучая историю болезни пациента и исследуя, сколь часто наследственное заболевание проявлялось среди его близких и дальних родственников, давал заключение о том, может ли у пациента появиться ребенок с такой патологией; и если может, то какова вероятность данного события (например, 1/2, 1/10, или 1/100). Основываясь на этой информации, супруги сами решали, иметь им ребенка или не иметь. Бурное развитие молекулярной биологии существенно приблизило нас к заветной цели - лечению наследственных болезней. Для этого прежде всего необходимо найти среди множества генов человека тот, который ответствен за болезнь. Популяционная генетика помогает решить эту сложную задачу. Известны генетические метки - т.н. ДНК-маркеры, которые позволяют отметить в длинной нити ДНК, скажем, каждую тысячную или десятитысячную "бусинку". Исследуя больного, его родственников и здоровых лиц из популяции, можно установить, какой из маркеров сцеплен с геном болезни. С помощью специальных математических методов популяционные генетики выявляют тот участок ДНК, в котором расположен интересующий нас ген. После этого в работу включаются молекулярные биологи, которые детально анализируют этот отрезок ДНК и находят в нем дефектный ген. Таким способом картированы гены большинства наследственных болезней. Теперь врачи получили возможность в первые месяцы беременности прямо судить о здоровье будущего ребенка, а родители - решать вопрос, сохранять или не сохранять беременность, если заранее известно, что ребенок родится больным. Более того, уже предпринимаются попытки исправлять допущенные природой ошибки, устранять "поломки" в генах. С помощью ДНК-маркеров можно не только искать гены болезней. Используя их, проводят своеобразную паспортизацию индивидов. Такая ДНК-идентификация - распространенный вид судебно-медицинской экспертизы, позволяющий определить отцовство, опознать перепутанных в роддоме детей, выявить личность участников преступления, жертв катастроф и военных действий.
ПОПУЛЯЦИОННАЯ ГЕНЕТИКА И СЕЛЕКЦИЯ
Согласно теории Дарвина, отбор в природе направлен только на непосредственную пользу - выжить и размножиться. Например, у рыси окраска шерсти палево-дымчатая, а у льва - песчано-желтая. Окраска, как маскировочная одежда, служит тому, чтобы особь сливалась с местностью. Это позволяет хищникам незаметно подкрадываться к жертве или выжидать. Поэтому хотя цветовые вариации постоянно появляются в природе, дикие кошки с такой "меткой" не выживают. Лишь человек с его вкусовыми пристрастиями создает все условия для жизни домашних кошек самых разнообразных окрасок. Переходя к оседлому образу жизни, люди уходили от охоты на животных и собирательства растений к их воспроизводству, резко уменьшая свою зависимость от катаклизмов природы. Тысячелетиями размножая особей с нужными признаками и ведя тем самым отбор соответствующих генов из генофондов популяций, люди постепенно создали все те сорта домашних растений и породы животных, что нас окружают. Это был тот же отбор, что проводила миллионами лет природа, но только теперь в роли природы выступил человек, направляемый разумом. С началом развития популяционный генетики, т.е. с середины 20 в., селекция пошла по научному пути, а именно по пути прогнозирования ответа на отбор и выбора оптимальных вариантов селекционной работы. Например, в скотоводстве племенная ценность каждого животного вычисляется сразу по многим признакам продуктивности, определяемым не только у данного животного, но и у его родственников (матерей, сестер, потомков и др.). Все это сводится в некий общий индекс, учитывающий как генетическую обусловленность признаков продуктивности, так и их экономическую значимость. Это особенно важно при оценке производителей, у которых собственную продуктивность определить невозможно (например, у быков в молочном скотоводстве или у петухов яичных пород). С внедрением искусственного осеменения появилась необходимость в разносторонней популяционной оценке племенной ценности производителей при их использовании в разных стадах с разным уровнем кормления, содержания и продуктивности. В селекции растений популяционный подход помогает количественно оценить генетическую способность линий и сортов давать перспективные гибриды и прогнозировать их приспособленность и продуктивность в разных по климату и почвам регионах. Таким образом, из чисто академической отрасли знаний, какой она была до недавнего времени, популяционная генетика превращается в науку, решающую многие теоретические и прикладные задачи.
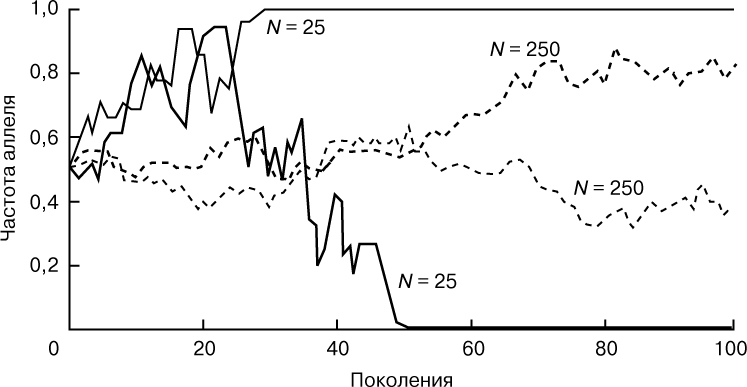
ИЗМЕНЕНИЕ ЧАСТОТ АЛЛЕЛЕЙ ПРИ ДРЕЙФЕ. Представлены результаты моделирования процесса дрейфа генов в двух популяциях численности N = 25 и двух популяциях численности N = 250, при частоте аллеля равной 0,5 в исходном поколении. Под действием дрейфа частота данного аллеля хаотически меняется из поколения в поколение, причем "скачки" частот более выражены в популяциях меньшей численности. За 50 поколений дрейф привел к фиксации аллеля в одной популяции численности N = 25, и к полной его элиминации - в другой. В популяциях большей численности этот аллель еще находится на промежуточных частотах, но популяции уже заметно отличаются друг от друга начиная с 60-го поколения.
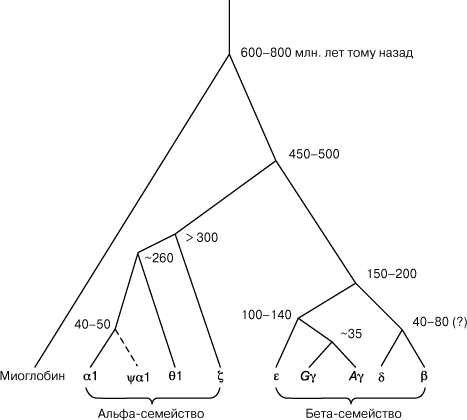
ЭВОЛЮЦИЯ ГЛОБИНОВЫХ ГЕНОВ ЧЕЛОВЕКА. На диаграмме показана часть глобиновых генов человека с оценкой времени их дивергенции (расхождения) друг от друга. Видно, что гены, кодирующие альфа- и бета-цепи гемоглобина, дивергировали друг от друга примерно полмиллиарда лет тому назад, а их предок и миоглобин отделились друг от друга на несколько сот миллионов лет раньше. Затем произошли дальнейшие дупликации и их дивергенция в пределах этих семейств генов. При этом возникли как гены, несущие определенные функции, так и гены, которые не имеют какой-либо функции, - т.н. псевдогены (помечено греческой буквой y).
ЛИТЕРАТУРА
Тимофеев-Ресовский Н.В., Яблоков А.В., Глотов Н.В. Очерк учения о популяции. М., 1973 Айала Ф., Кайгер Дж. Современная генетика, тт. 1-3, М., 1988 Фогель Ф., Мотульски А. Генетика человека, тт. 1-3. М., 1990
Энциклопедия Кольера. — Открытое общество. 2000.