- Физиологические особенности культур микроскопических водорослей как фотосинтезирующих систем
- Физиологические особенности культур микроскопических водорослей как фотосинтезирующих систем
-
Культуры микроскопических водорослей являются целостными фотосинтезирующими системами. Для получения высокой фотосинтетической продуктивности необходимо знание физиологических особенностей этих систем, а не только физиологии отдельной клетки.Как бы ни различались современные методы культивирования водорослей, все они основаны на снабжении клеток достаточным количеством света, углекислоты и других питательпых веществ.Исследование функциональной зависимости продуктивности культур микроводорослей от напряженности и сочетания значимых факторов — необходимая предпосылка для разработки физиологически обоснованных режимов их культивирования и принципов интенсификации их продуктивности.
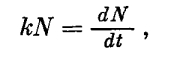 Как следует из приведенного уравнения, при выращивании в условиях экспоненциального роста, но при низкой плотности суспензии можно получить высокий коэффициент размножения культуры, однако это еще не значит, что будет получен высокий суммарный урожай. Такая культура хороша для изучения потенциальных свойств клетки, но недостаточна для использования при массовом культивировании. В реальных условиях культивирования суммарная фотосинтетическая продуктивность водорослей тем выше, чем при более высокой плотности суспензии сохраняется высокая скорость размножения. И далеко не всегда результаты, полученные при исследовании разбавленных суспензий (что может быть приравнено к изучению отдельной клетки), могут быть перенесены на культуры высокой плотности. Например, впервые полученная в 1953 г. термофильная форма хлореллы (Сорокин и Майерс) прославилась еще и тем, что для нее в определенных условиях света и температуры в суспензии с низкой плотностью клеток было найдено тысячекратное увеличение числа клеток в сутки (10 удвоений). Следовательно, если создать этой культуре оптимальные условия экспоненциального роста при плотности хотя бы 50 млн. клеток в 1 мл, то 1 л такой культуры мог бы обеспечить урожай в 500 г/л в сутки и это была бы высокоинтенсивная культура водорослей, размножающаяся с максимальной скоростью и дающая высокие урожаи за максимально короткий срок. Однако организация такого рода культивирования практически трудноосуществима, так как уже при четвертом удвоении плотность культуры поднимается до 800 млн. в 1 мл и, естественно, при этом резко меняются все условия культивирования. Для правильного освещения суспензии этот литр культуры требовалось бы распределить очень тонким слоем по большой поверхности, в результате чего конструкция установки стала бы чрезвычайно сложной и громоздкой. Таким образом, для правильной организации культивирования водорослей с высокой фотосинтетической продуктивностью необходимо изучение действия хотя бы основных факторов (свет, температура, концентрация С02 и т. д.) на фотосинтезирующие системы высокой плотности.В отличие от популяций гетеротрофных организмов, где каждому из факторов, определяющих продуктивность культуры, обеспечен одинаковый доступ ко всем индивидам в любой рассматриваемый момент времени, в популяции автотрофных клеток исключение составляет световой фактор, который в силу оптических свойств культуры как сильно поглощающей свет среды достаточно быстро становится ограничивающим. Кратковременность экспоненциальной фазы роста в интенсивно растущих культурах и выход на линейную стадию роста объясняется быстрым формированием полностью поглощающего свет слоя, численность клеток в котором при заданной интенсивности света остается величиной постоянной, несмотря на увеличение плотности по мере роста всей популяции. Формирование такого слоя определяет достаточно резкий переход от процесса возрастания интенсивности фотосинтеза по мере роста культуры к его стационарному состоянию (и постоянству прироста числа клеток и биомассы в единицу времени), несмотря на продолжающееся суммарное увеличение числа клеток. При этом плотность культуры, при которой происходит указанный переход, различна в реакторах с разной толщиной слоя суспензии, однако поверхностная концентрация клеток в момент этого перехода остается величиной постоянной.
Как следует из приведенного уравнения, при выращивании в условиях экспоненциального роста, но при низкой плотности суспензии можно получить высокий коэффициент размножения культуры, однако это еще не значит, что будет получен высокий суммарный урожай. Такая культура хороша для изучения потенциальных свойств клетки, но недостаточна для использования при массовом культивировании. В реальных условиях культивирования суммарная фотосинтетическая продуктивность водорослей тем выше, чем при более высокой плотности суспензии сохраняется высокая скорость размножения. И далеко не всегда результаты, полученные при исследовании разбавленных суспензий (что может быть приравнено к изучению отдельной клетки), могут быть перенесены на культуры высокой плотности. Например, впервые полученная в 1953 г. термофильная форма хлореллы (Сорокин и Майерс) прославилась еще и тем, что для нее в определенных условиях света и температуры в суспензии с низкой плотностью клеток было найдено тысячекратное увеличение числа клеток в сутки (10 удвоений). Следовательно, если создать этой культуре оптимальные условия экспоненциального роста при плотности хотя бы 50 млн. клеток в 1 мл, то 1 л такой культуры мог бы обеспечить урожай в 500 г/л в сутки и это была бы высокоинтенсивная культура водорослей, размножающаяся с максимальной скоростью и дающая высокие урожаи за максимально короткий срок. Однако организация такого рода культивирования практически трудноосуществима, так как уже при четвертом удвоении плотность культуры поднимается до 800 млн. в 1 мл и, естественно, при этом резко меняются все условия культивирования. Для правильного освещения суспензии этот литр культуры требовалось бы распределить очень тонким слоем по большой поверхности, в результате чего конструкция установки стала бы чрезвычайно сложной и громоздкой. Таким образом, для правильной организации культивирования водорослей с высокой фотосинтетической продуктивностью необходимо изучение действия хотя бы основных факторов (свет, температура, концентрация С02 и т. д.) на фотосинтезирующие системы высокой плотности.В отличие от популяций гетеротрофных организмов, где каждому из факторов, определяющих продуктивность культуры, обеспечен одинаковый доступ ко всем индивидам в любой рассматриваемый момент времени, в популяции автотрофных клеток исключение составляет световой фактор, который в силу оптических свойств культуры как сильно поглощающей свет среды достаточно быстро становится ограничивающим. Кратковременность экспоненциальной фазы роста в интенсивно растущих культурах и выход на линейную стадию роста объясняется быстрым формированием полностью поглощающего свет слоя, численность клеток в котором при заданной интенсивности света остается величиной постоянной, несмотря на увеличение плотности по мере роста всей популяции. Формирование такого слоя определяет достаточно резкий переход от процесса возрастания интенсивности фотосинтеза по мере роста культуры к его стационарному состоянию (и постоянству прироста числа клеток и биомассы в единицу времени), несмотря на продолжающееся суммарное увеличение числа клеток. При этом плотность культуры, при которой происходит указанный переход, различна в реакторах с разной толщиной слоя суспензии, однако поверхностная концентрация клеток в момент этого перехода остается величиной постоянной.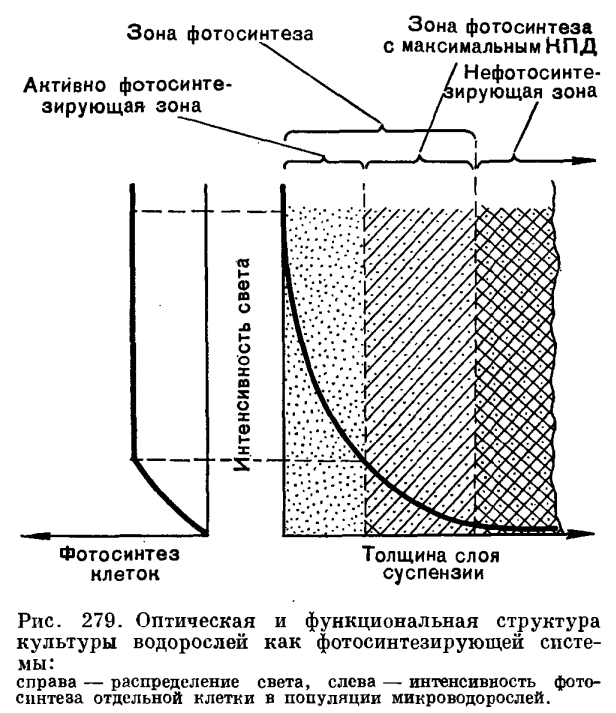 В плотной, вышедшей на линейную фазу роста культуре суммарная фотосинтетическая продуктивность суспензии определяется в каждый элементарный отрезок времени работой только тех клеток, которые находятся в зоне облучения (рис. 279). При этом часть клеток в зоне облучения осуществляет фотосинтез на линейном участке, а часть — на плато световой кривой фотосинтеза отдельной клетки. Остальные клетки (вне «зоны фотосинтеза») в принципе являются балластными и не дают вклад в общий фотосинтез популяции, поскольку длительность световых реакций фотосинтеза порядка 10/-3 сек существенно короче периода миграции клеток между просвечиваемыми и непросвечиваемыми зонами суспензии, что достигается за счет перемешивания культуры.Описанные закономерности положены в основу ряда разработаннцх в настоящее время математических моделей фотосинтетической продуктивности микроводорослей и используются при конструкторской разработке технологии, реакторов и управляющей аппаратуры для высокоинтенсивного культивирования этих организмов.
В плотной, вышедшей на линейную фазу роста культуре суммарная фотосинтетическая продуктивность суспензии определяется в каждый элементарный отрезок времени работой только тех клеток, которые находятся в зоне облучения (рис. 279). При этом часть клеток в зоне облучения осуществляет фотосинтез на линейном участке, а часть — на плато световой кривой фотосинтеза отдельной клетки. Остальные клетки (вне «зоны фотосинтеза») в принципе являются балластными и не дают вклад в общий фотосинтез популяции, поскольку длительность световых реакций фотосинтеза порядка 10/-3 сек существенно короче периода миграции клеток между просвечиваемыми и непросвечиваемыми зонами суспензии, что достигается за счет перемешивания культуры.Описанные закономерности положены в основу ряда разработаннцх в настоящее время математических моделей фотосинтетической продуктивности микроводорослей и используются при конструкторской разработке технологии, реакторов и управляющей аппаратуры для высокоинтенсивного культивирования этих организмов.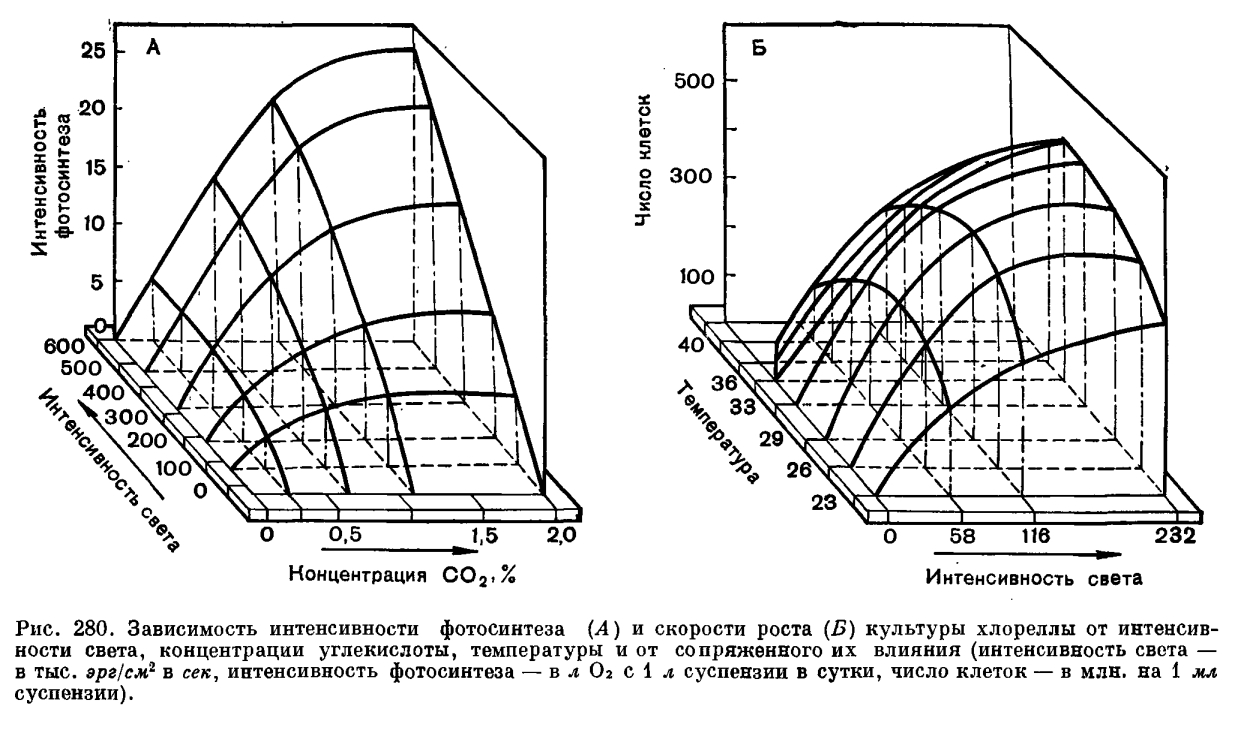 Углекислота может вводиться в автотрофно выращиваемые культуры водорослей различными методами. Наиболее распространенный способ — это подача углекислоты в виде газовоздушной смеси, осуществляющей, помимо снабжения водорослей источником углерода, функцию перемешивания культуры. Найденные применительно к условиям высокоинтенсивного культивирования хлореллы насыщающие и полунасыщающие концентрации С02 (рис. 280) оказались более высокими, чем описаны в литературе для разбавленных суспензий, но не превышающими значений 1,6 — 1,7% С02 в газовой фазе (35 — 40x10/-5 моля растворенной С02 на 1 л) при насыщающих иптенсивностях света и плотности суспензии 600 млн. клеток в 1 мл. На рисунке 280, А в виде трехмерного графика представлены семейства углекислотных и световых кривых фотосинтеза хлореллы и зависимость фотосинтетической продуктивности культуры от взаимосвязи этих характеристик. Так, при наибольшей, насыщающей интенсивности света продуктивность культуры в 4 раза выше при концентрации 1% С02, чем при 0,25% С02, и обратно: при одной и той же концентрации С02 (например, 1% С02) продуктивность тем выше, чем выше интенсивность света.Водоросли различаются по световым и температурным оптимумам для роста и фотосинтеза.В пределах рода Chlorella есть светолюбивые и теневыносливые культуры с меньшим или большим содержанием хлорофилла, с большей или меньшей активностью единицы хлорофилла, есть термофильные, мезофильные и психрофильные (хорошо растущие при температуре +5... +10 °С) штаммы. Каждый штамм в той или иной мере обладает адаптационными способностями. Но у каждой культуры свой генетически закрепленный диапазон требований к освещенности и температуре.На основании известных экспериментальных данных можно охарактеризовать в целом группу мезофильных культур как более тенелюбивую, а термофильных культур как более светолюбивую. Так, мезофильные формы обладают более низким и более узким плато светового насыщения для роста по сравнению с термофильными культурами.Применение культур высоких плотностей имеет особенно большой смысл при высоких освещенностях, позволяющих максимально нриблизиться к потенциально возможным скоростям роста культуры. На рисунке 280, Б приведены световые и температурные кривые зависимости роста термофильной культуры. Трехмерный график хорошо показывает тесную функциональную взаимообусловленность характера температурных и световых кривых роста и необходимость тщательного подбора оптимальных сочетаний этих параметров для высокопродуктивного культивирования. Для каждой интенсивности света существует своя температурная кривая роста и продуктивности культуры, так же как для каждой температуры имеется своя световая кривая. По мере повышения освещенности температурный оптимум смещается в сторону более высоких значений температуры, так же как и при повышении температуры для получения максимальной продуктивности необходимо соответствующее изменение освещенности. Важно, что в культурах высоких плотностей, как и в разбавленных суспензиях, четко обнаруживается эффект фотоингибирования роста (угнетение ростовых процессов под действием света) культуры при температуре, не сочетающейся с примененной интенсивностью света.Если учесть, что температурные условия являются диффузно (или равномерно) действующим фактором и не зависят от плотности культуры, а облученность отдельных клеток в популяции, наоборот, сильно зависит от плотности и оптических свойств суспензии, то становится ясно: чтобы найти оптимальные сочетания облученности и температуры для каждой культуры, необходимо снять световые и температурные характеристики не только для клетки, но и для популяции.Как показано на примерах влияния света, температуры, концентрации С02, для получения высокой продуктивности культуры существенное значение имеет не столько зависимость роста и фотосинтеза водорослей от каждого из этих факторов в отдельности, сколько их сопряженное действие. Обнаруженные взаимосвязи могут быть показаны в виде схемы:
Углекислота может вводиться в автотрофно выращиваемые культуры водорослей различными методами. Наиболее распространенный способ — это подача углекислоты в виде газовоздушной смеси, осуществляющей, помимо снабжения водорослей источником углерода, функцию перемешивания культуры. Найденные применительно к условиям высокоинтенсивного культивирования хлореллы насыщающие и полунасыщающие концентрации С02 (рис. 280) оказались более высокими, чем описаны в литературе для разбавленных суспензий, но не превышающими значений 1,6 — 1,7% С02 в газовой фазе (35 — 40x10/-5 моля растворенной С02 на 1 л) при насыщающих иптенсивностях света и плотности суспензии 600 млн. клеток в 1 мл. На рисунке 280, А в виде трехмерного графика представлены семейства углекислотных и световых кривых фотосинтеза хлореллы и зависимость фотосинтетической продуктивности культуры от взаимосвязи этих характеристик. Так, при наибольшей, насыщающей интенсивности света продуктивность культуры в 4 раза выше при концентрации 1% С02, чем при 0,25% С02, и обратно: при одной и той же концентрации С02 (например, 1% С02) продуктивность тем выше, чем выше интенсивность света.Водоросли различаются по световым и температурным оптимумам для роста и фотосинтеза.В пределах рода Chlorella есть светолюбивые и теневыносливые культуры с меньшим или большим содержанием хлорофилла, с большей или меньшей активностью единицы хлорофилла, есть термофильные, мезофильные и психрофильные (хорошо растущие при температуре +5... +10 °С) штаммы. Каждый штамм в той или иной мере обладает адаптационными способностями. Но у каждой культуры свой генетически закрепленный диапазон требований к освещенности и температуре.На основании известных экспериментальных данных можно охарактеризовать в целом группу мезофильных культур как более тенелюбивую, а термофильных культур как более светолюбивую. Так, мезофильные формы обладают более низким и более узким плато светового насыщения для роста по сравнению с термофильными культурами.Применение культур высоких плотностей имеет особенно большой смысл при высоких освещенностях, позволяющих максимально нриблизиться к потенциально возможным скоростям роста культуры. На рисунке 280, Б приведены световые и температурные кривые зависимости роста термофильной культуры. Трехмерный график хорошо показывает тесную функциональную взаимообусловленность характера температурных и световых кривых роста и необходимость тщательного подбора оптимальных сочетаний этих параметров для высокопродуктивного культивирования. Для каждой интенсивности света существует своя температурная кривая роста и продуктивности культуры, так же как для каждой температуры имеется своя световая кривая. По мере повышения освещенности температурный оптимум смещается в сторону более высоких значений температуры, так же как и при повышении температуры для получения максимальной продуктивности необходимо соответствующее изменение освещенности. Важно, что в культурах высоких плотностей, как и в разбавленных суспензиях, четко обнаруживается эффект фотоингибирования роста (угнетение ростовых процессов под действием света) культуры при температуре, не сочетающейся с примененной интенсивностью света.Если учесть, что температурные условия являются диффузно (или равномерно) действующим фактором и не зависят от плотности культуры, а облученность отдельных клеток в популяции, наоборот, сильно зависит от плотности и оптических свойств суспензии, то становится ясно: чтобы найти оптимальные сочетания облученности и температуры для каждой культуры, необходимо снять световые и температурные характеристики не только для клетки, но и для популяции.Как показано на примерах влияния света, температуры, концентрации С02, для получения высокой продуктивности культуры существенное значение имеет не столько зависимость роста и фотосинтеза водорослей от каждого из этих факторов в отдельности, сколько их сопряженное действие. Обнаруженные взаимосвязи могут быть показаны в виде схемы: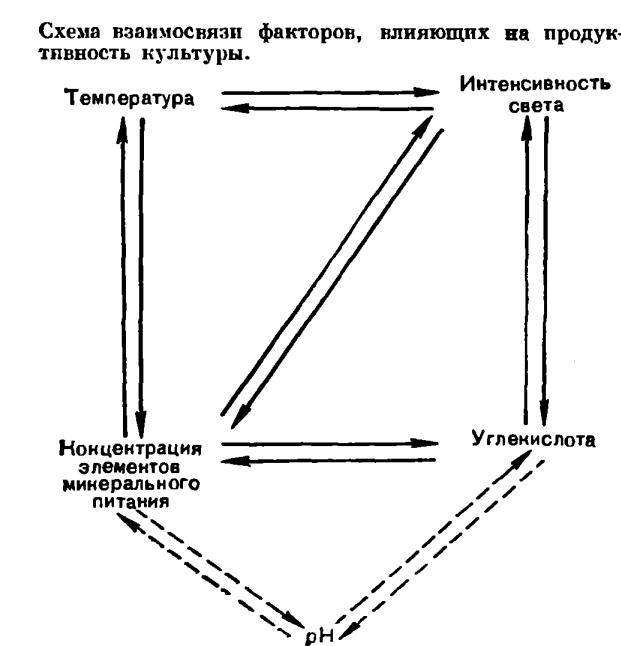 Поскольку в культурах высоких плотностей для обеспечения интенсивного роста применяют высокие освещенности, высокие температуры и высокие концентрации солей, т. е. создается напряженность всех факторов среды, «отставание» какого-либо одного фактора, нарушая сбалансированное взаимодействие всех факторов, может повлечь за собой не только снижение продуктивности до уровня действия этого минимального фактора, но во многих случаях и гибель культуры.При создании систем культивирования водорослей высокой фотосинтетической производительности одним из первых встает вопрос о применении культур, генотипические свойства которых отвечали бы заданным условиям. Например, водоросли в интенсивно растущих плотных популяциях должны переносить кратковременную переоблученность, т. е. культура должна иметь высокое и продолженное плато светового насыщения для роста и фотосинтеза. Этим свойством в большей степени обладают термофильные, штаммы.При массовом культивировании под открытым небом более эффективными в определенные периоды могут быть мезофильные культуры. Японские исследователи провели выращивание смешанной культуры водорослей, состоящей из мезофильного и термофильного штаммов. В жаркие месяцы преимущественно развивалась термофильная форма, в остальное время — мезофильная.Отбором в природе и селекционно-генетическими методами получено значительное число мезофильных и термофильных высокопродуктивных форм хлореллы. Сравнительное изучение различных штаммов водорослей показывает широкое их разнообразие в отношении к температуре, освещенности, способности роста в плотных популяциях. Это открывает возможность направленного подбора форм водорослей применительно к конкретным условиям и целям их культивирования.Отбор и поддержание в коллекции перспективных форм является важной задачей.
Поскольку в культурах высоких плотностей для обеспечения интенсивного роста применяют высокие освещенности, высокие температуры и высокие концентрации солей, т. е. создается напряженность всех факторов среды, «отставание» какого-либо одного фактора, нарушая сбалансированное взаимодействие всех факторов, может повлечь за собой не только снижение продуктивности до уровня действия этого минимального фактора, но во многих случаях и гибель культуры.При создании систем культивирования водорослей высокой фотосинтетической производительности одним из первых встает вопрос о применении культур, генотипические свойства которых отвечали бы заданным условиям. Например, водоросли в интенсивно растущих плотных популяциях должны переносить кратковременную переоблученность, т. е. культура должна иметь высокое и продолженное плато светового насыщения для роста и фотосинтеза. Этим свойством в большей степени обладают термофильные, штаммы.При массовом культивировании под открытым небом более эффективными в определенные периоды могут быть мезофильные культуры. Японские исследователи провели выращивание смешанной культуры водорослей, состоящей из мезофильного и термофильного штаммов. В жаркие месяцы преимущественно развивалась термофильная форма, в остальное время — мезофильная.Отбором в природе и селекционно-генетическими методами получено значительное число мезофильных и термофильных высокопродуктивных форм хлореллы. Сравнительное изучение различных штаммов водорослей показывает широкое их разнообразие в отношении к температуре, освещенности, способности роста в плотных популяциях. Это открывает возможность направленного подбора форм водорослей применительно к конкретным условиям и целям их культивирования.Отбор и поддержание в коллекции перспективных форм является важной задачей. Развитие исследований по массовой культуре водорослей дало толчок к совершенствованию методов лабораторного их выращивания, благодаря чему возник метод лабораторного интенсивного культивирования микроводорослей. На рисунке 281 представлен один из способов современного лабораторного культивирования одноклеточных водорослей, обеспечивающий интенсивный рост культур, относящихся к различным систематическим группам (зеленые — Chlorella, Scenedesmus, Ankistrodesmus, Dunaliella, Chlamydomonas; сине-зеленые — Anacystis, Spirulina, Anabaena, Synechocystis, Aphanizomenon, Nostoc, Tolypothrix и др.).
Развитие исследований по массовой культуре водорослей дало толчок к совершенствованию методов лабораторного их выращивания, благодаря чему возник метод лабораторного интенсивного культивирования микроводорослей. На рисунке 281 представлен один из способов современного лабораторного культивирования одноклеточных водорослей, обеспечивающий интенсивный рост культур, относящихся к различным систематическим группам (зеленые — Chlorella, Scenedesmus, Ankistrodesmus, Dunaliella, Chlamydomonas; сине-зеленые — Anacystis, Spirulina, Anabaena, Synechocystis, Aphanizomenon, Nostoc, Tolypothrix и др.).
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.
.