- Семейство рестиевые (Restionaceae)
- Семейство рестиевые (Restionaceae)
-
В некоторых районах Южной Африки и Австралии место осоковых и злаков в растительных группировках занимают травянистые растения, по строению соцветий очень похожие на осоковые, но имеющие оригинальный внешний облик, который является следствием полной или почти полной редукции листовых пластинок и большой жесткости прутьевидных стеблей, взявших на себя функцию фотосинтеза и транспирации. Это представители семейства рестиевых, к которому принадлежит около 300 видов из 30 родов. Рестиевые особенно характерны для внетропических стран южного полушария и потому мало известны жителям северного полушария. Лишь один вид этого семейства — лептокарпус разъединенный (Leptocarpus disjunctus) встречается к северу от экватора — во Вьетнаме и Малайзии; другой вид этого же, преимущественно австралийского рода — лептокарпус чилийский (L. chilensis) является единственным представителем рестиевых в Америке (Чили и Южная Аргентина).
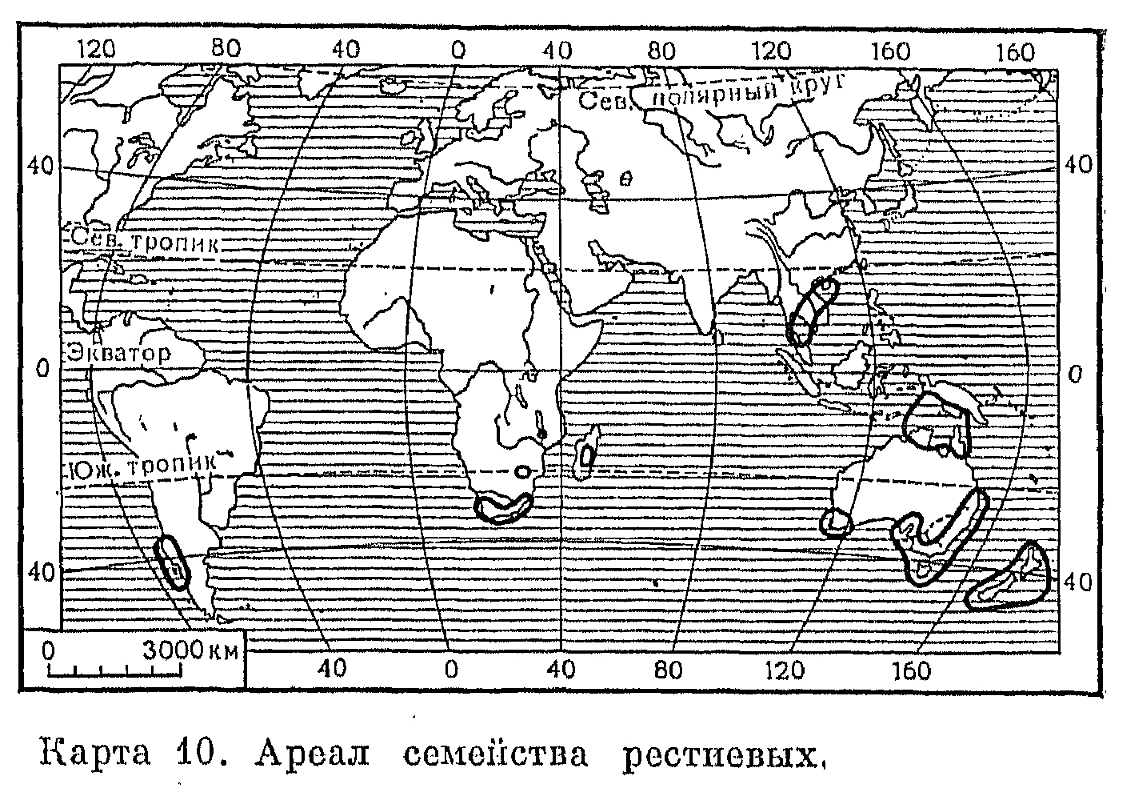 В пределах общего ареала семейства выделяют две области, наиболее богатые родами и видами рестиевых. Одна из них — внетропическая Австралия, включая Тасманию, где известны 13 эндемичных родов этого семейства. Лишь немногие виды рестиевых заходят на остров Чатам и в Новую Зеландию, во флоре которой имеется и один эндемичный род рестиевых — спорадантус (Sporadanthus). Второй основной областью распространения семейства является южная оконечность Африки, выделяемая, несмотря на небольшие размеры территории, в особое Капское флористическое царство. Здесь представлены 12 эндемичных родов рестиевых, и только немногие виды этого семейства заходят отсюда в тропическую Африку (до Малави) и на юг Мадагаскара. Наиболее богатый видами (около 125) род рестио (Restio) встречается в обеих основных областях распространения семейства, однако в последнее время выявлено много существенных анатомических, палинологических и биохимических различий между южноафриканскими и австралийскими видами, и они, по-видимому, будут отнесены к разным родам.Рестиевые — многолетние травянистые растения с длинными ползучими корневищами или короткокорневищные и тогда образующие густые дерновины. Корневища обычно несут кожистые чешуевидные листья, у основания которых по 1 — 2 отходят придаточные корни. Лишь у западноавстралийской дильсии (Dielsia) корневища лишены чешуи, но покрыты густым буроватым войлоком, подобно корневищам некоторых папоротников. Надземные побеги большинства рестиевых представлены только у основания разветвленными жесткими цилиндрическими стеблями, несущими соцветия и обычно достигающими в высоту 25 — 100 см. Лишь у немногих видов (например, у капской элегии мутовчатой — Elegia verticillaris) они достигают в высоту 2 м и в толщину 4 (8) см. Виды очень обособленного австралийского рода анартрия (Anarthria) имеют сильно сплюснутые с боков безлистные стрелкообразные стебли, а у капского рода антохортус (Anthochortus) они острочетырехгранные. Удлиненные междоузлия стеблей могут быть как полыми, так и заполненными сердцевиной. У видов некоторых родов, кроме неразветвленных репродуктивных побегов, имеются еще более или менее разветвленные вегетативные побеги (например, у рестио остроконечного — Restio cuspidatus, рис. 188). У других видов стебли могут ветвиться во всех узлах, и в этом случае из пазух влагалищеобразных листьев могут выходить или пучки фертильных и стерильных ветвей, или только стерильные ветви. У элегии мутовчатой последние располагаются в узлах ложными мутовками и весь побег напоминает по облику побег гигантского хвоща или харовой водоросли.
В пределах общего ареала семейства выделяют две области, наиболее богатые родами и видами рестиевых. Одна из них — внетропическая Австралия, включая Тасманию, где известны 13 эндемичных родов этого семейства. Лишь немногие виды рестиевых заходят на остров Чатам и в Новую Зеландию, во флоре которой имеется и один эндемичный род рестиевых — спорадантус (Sporadanthus). Второй основной областью распространения семейства является южная оконечность Африки, выделяемая, несмотря на небольшие размеры территории, в особое Капское флористическое царство. Здесь представлены 12 эндемичных родов рестиевых, и только немногие виды этого семейства заходят отсюда в тропическую Африку (до Малави) и на юг Мадагаскара. Наиболее богатый видами (около 125) род рестио (Restio) встречается в обеих основных областях распространения семейства, однако в последнее время выявлено много существенных анатомических, палинологических и биохимических различий между южноафриканскими и австралийскими видами, и они, по-видимому, будут отнесены к разным родам.Рестиевые — многолетние травянистые растения с длинными ползучими корневищами или короткокорневищные и тогда образующие густые дерновины. Корневища обычно несут кожистые чешуевидные листья, у основания которых по 1 — 2 отходят придаточные корни. Лишь у западноавстралийской дильсии (Dielsia) корневища лишены чешуи, но покрыты густым буроватым войлоком, подобно корневищам некоторых папоротников. Надземные побеги большинства рестиевых представлены только у основания разветвленными жесткими цилиндрическими стеблями, несущими соцветия и обычно достигающими в высоту 25 — 100 см. Лишь у немногих видов (например, у капской элегии мутовчатой — Elegia verticillaris) они достигают в высоту 2 м и в толщину 4 (8) см. Виды очень обособленного австралийского рода анартрия (Anarthria) имеют сильно сплюснутые с боков безлистные стрелкообразные стебли, а у капского рода антохортус (Anthochortus) они острочетырехгранные. Удлиненные междоузлия стеблей могут быть как полыми, так и заполненными сердцевиной. У видов некоторых родов, кроме неразветвленных репродуктивных побегов, имеются еще более или менее разветвленные вегетативные побеги (например, у рестио остроконечного — Restio cuspidatus, рис. 188). У других видов стебли могут ветвиться во всех узлах, и в этом случае из пазух влагалищеобразных листьев могут выходить или пучки фертильных и стерильных ветвей, или только стерильные ветви. У элегии мутовчатой последние располагаются в узлах ложными мутовками и весь побег напоминает по облику побег гигантского хвоща или харовой водоросли. Хорошо развитые мечевидные прикорневые листья, напоминающие листья ирисов, но более жесткие, имеются лишь у видов анартрии, например у анартрии шероховатой (Anarthria scabra, рис. 188). У других рестиевых листья обычно видоизменены в кожистые или перепончатые, долго сохраняющиеся на растении или опадающие влагалища, края которых обычно перекрываются, но, в отличие от влагалищ осоковых, не срастаются друг с другом. У некоторых родов (например, у австралийского онихосепалума — Onychosepalum) все узлы и расположенные на них влагалища тесно сближены у основания надземных побегов, но обычно на побегах имеется несколько расставленных узлов и влагалищ. Более или менее редуцированные листовые пластинки, обычно шиловидной формы и часто колючие на верхушке, есть у многих рестиевых, особенно на вегетативных ветвях. У рестио остроконечного (рис. 188) и некоторых других видов этого рода на границе влагалища с пластинкой расположен перепончатый вырост — язычок, сходный с язычком злаков и некоторых водных однодольных. По строению устьиц, которые здесь так называемого граминоидного (злакового) типа, рестиевые более сходны со злаками, чем с осоковыми. Расположение влагалищеобразных листьев на стеблях у рестиевых всегда двурядное, как у злаков.Цветки рестиевых почти всегда однополые и двудомные, хотя рудимент гинецея в мужских цветках и стаминодии в женских нередко присутствуют. Мужские и женские цветки в одних и тех же соцветиях имеются только у монотипного капского рода филокомус (Phylocomus), а всегда обоеполые цветки — у австралийского вида лепиродия обоеполая (Lepyrodia hermaphrodita). Подобно злакам и осоковым, для рестиевых очень характерны элементарные соцветия — колоски, которые обычно, в свою очередь, собраны в колосовидные, кистевидные, метельчатые или зонтиковидные общие соцветия, имеющие у своего основания, а часто и у основания своих ветвей влагалищеобразные верхушечные листья. Внешне колоски представителей этого семейства особенно похожи на колоски некоторых осоковых, например видов камыша (Scirpus). Соцветия с мужскими и соцветия с женскими цветками в пределах одного и того же вида обычно настолько отличаются по внешнему виду, что их нетрудно принять за принадлежащие разным видам и даже родам. Женские соцветия меньше разветвлены, число цветков в женских колосках обычно уменьшено, а у ряда родов (например, харперия — Harperia, гиполена — Hypolaena и др.) они имеют только один цветок. Каждый цветок в колоске располагается в пазухе перепончатой или кожистой кроющей чешуи — прицветника, а у двух родов — анартрии и лепиродии — по бокам цветков имеются еще по 2 прицветничка. Самые нижние в колоске кроющие чешуи обычно не несут в своих пазухах цветков.Околоцветник у рестиевых перепончатый или пленчатый, обычно состоящий из 4 или 6 расположенных 2 кругами бесцветных или слабоокрашенных сегментов более или менее одинакового строения. Сегменты внутреннего круга могут срастаться у своего основания. Кроме того, у некоторых видов один из сегментов, а иногда и все сегменты этого круга могут редуцироваться, и тогда околоцветник состоит из 3 или 5 сегментов. У австралийского рода локсокария (Loxocarya) околоцветник в женских цветках полностью отсутствует. У большинства родов семейства цветки актиноморфные или почти актиноморфные, однако у некоторых родов они становятся зигоморфными. Так, у капского тамнохортуса колосоносного (Thamnochortus spicigerus) боковые сегменты сросшегося при основании околоцветника сильно увеличены и снабжены килем, переходящим в широкий крыловидный вырост (рис. 188). У этого вида околоцветник сохраняется при плодах, играя роль летучки.Мужские цветки рестиевых имеют 3, реже 2 тычинки, расположенные супротивно внутренним сегментам околоцветника. Тычиночные нити обычно свободные, тонкие, с одногнездными, реже двугнездными (у примитивных и в других отношениях австралийских родов анартрии, лигинии — Lyginia и хопкинсии — Hopkinsia) пыльниками, прикрепленными к нити спинной стороной и вскрывающимися интрорзно поперечной щелью. Лишь у лигинии (рис. 188) нити тычинок срастаются в нижней части, образуя колонку. У этого последнего рода, как и у некоторых других родов, имеются надсвязники в виде шипиковидного выроста. Пыльцевые зерна рестиевых имеют однопоровую оболочку и обычно очень сходны с пыльцевыми зернами злаков.Синкарпный или паракарпный гинецей женских цветков рестиевых обычно состоит из 3 или 2 плодолистиков, лишь у родов тамнохортус и лепидоболус (Lepidobolus) он псевдомономерный в результате редукции переднего и одного из двух боковых гнезд первично-3-гнездной завязи. В двучленных женских цветках (например, у многочисленных видов рестио) два гнезда завязи располагаются супротивно двум наружным сегментам околоцветника. Ветви столбика обычно соответствуют числу гнезд завязи и в разной степени срастаются друг с другом. Волосистая рыльцевая поверхность находится на их внутренней стороне. Однако у некоторых родов гинецей имеет только один неразветвленный столбик, что связано или с редукцией двух гнезд завязи (у австралийского рода тамнохортус), или с полным срастанием всех ветвей столбика (например, у капской харперии). У гиподискуса (Hypodiscus) и вильденовии (Willdenowia) завязь, а позднее и плод располагаются на короткой дисковидной ножке, а у дильсии основание двухраздельного столбика пирамидообразно утолщено, как это бывает у некоторых родов осоковых. В каждом развитом гнезде гинецея имеется только один висячий ортотропный семязачаток.У большинства рестиевых плоды 2 — 3-семянные коробочки, раскрывающиеся продольной щелью по выступающим в виде ребер средним жилкам плодолистиков, т. е. локулицидно. Лишь немногие роды, например элегия и лепидоболус, имеют нераскрывающиеся орехообразные плоды с одним семенем. Семена всех рестиевых сходны по строению; они содержат обильный мучнистый эндосперм, у верхушки которого располагается маленький зародыш.Большинство рестиевых обитает на бедных питательными веществами, обычно кислых, песчаных или торфянистых почвах. Это растения преимущественно влажных открытых местообитаний, однако более или менее пересыхающих во время сухого периода года. Среди капских видов много обитателей междюнных западин на морских побережьях, а некоторые австралийские виды могут расти на солончаках и даже в условиях настоящей пустыни при почти полном отсутствии осадков. Однако ксероморфный облик свойствен всем рестиевым, даже растущим на непересыхающих болотах, что, по-видимому, объясняется историческими причинами (предки рестиевых могли расти в еще более влажных условиях) или физиологической сухостью местообитаний.Рестиевые являются ветроопыляемыми (анемофильными) растениями, причем возможность самоопыления у большинства видов исключается за счет двудомности — расположеиия мужских и женских цветков на разных особях. Специальных приспособлений для распространения семян у большинства рестиевых не отмечается. Выпадающие из коробочек семена могут переноситься ветром на некоторое расстояние от материнского растения, а затем разносятся или водными потоками, или на ногах животных вместе с комочками почвы. Лишь мелкие односемянные плоды рода тамнохортус, окутанные околоцветником с ширококрылатыми боковыми сегментами околоцветника, могут разноситься ветром на большие расстояния.Система рестиевых пока еще недостаточно разработана, хотя их обычно делят на две группы по числу гнезд в пыльниках: двугнезднопыльниковые (Diplantherae) — более примитивная группа и одногнезднопыльниковые (Haplantherae), к которым принадлежит большинство родов семейства. В последнее время рестиевые, исключая выделяемый в особое монотипное семейство род экдейоколея (Ecdeiocolea), делят на 2 подсемейства: собственно рестиевые (Restionoideae) и анартриевые (Anarthrioideae), к которому принадлежит только оригинальный австралийский род анартрия (рис. 188), обладающий рядом примитивных признаков (хорошо развитые листья, присутствие прицветничков и др.).Хозяйственное значение рестиевых незначительно. Их жесткие, безлистные или почти безлистные побеги не поедает или плохо поедает скот. Подобно тростнику, их можно использовать как покрытие для легких построек (особенно капский хондропеталум кровельный — Chondropetalum tectorum). В последнее время некоторые южноафриканские виды из родов элегия, хондропеталум, рестио и тамиохортус вводят в культуру в качестве декоративных растений, интересных своим своеобразным обликом.
Хорошо развитые мечевидные прикорневые листья, напоминающие листья ирисов, но более жесткие, имеются лишь у видов анартрии, например у анартрии шероховатой (Anarthria scabra, рис. 188). У других рестиевых листья обычно видоизменены в кожистые или перепончатые, долго сохраняющиеся на растении или опадающие влагалища, края которых обычно перекрываются, но, в отличие от влагалищ осоковых, не срастаются друг с другом. У некоторых родов (например, у австралийского онихосепалума — Onychosepalum) все узлы и расположенные на них влагалища тесно сближены у основания надземных побегов, но обычно на побегах имеется несколько расставленных узлов и влагалищ. Более или менее редуцированные листовые пластинки, обычно шиловидной формы и часто колючие на верхушке, есть у многих рестиевых, особенно на вегетативных ветвях. У рестио остроконечного (рис. 188) и некоторых других видов этого рода на границе влагалища с пластинкой расположен перепончатый вырост — язычок, сходный с язычком злаков и некоторых водных однодольных. По строению устьиц, которые здесь так называемого граминоидного (злакового) типа, рестиевые более сходны со злаками, чем с осоковыми. Расположение влагалищеобразных листьев на стеблях у рестиевых всегда двурядное, как у злаков.Цветки рестиевых почти всегда однополые и двудомные, хотя рудимент гинецея в мужских цветках и стаминодии в женских нередко присутствуют. Мужские и женские цветки в одних и тех же соцветиях имеются только у монотипного капского рода филокомус (Phylocomus), а всегда обоеполые цветки — у австралийского вида лепиродия обоеполая (Lepyrodia hermaphrodita). Подобно злакам и осоковым, для рестиевых очень характерны элементарные соцветия — колоски, которые обычно, в свою очередь, собраны в колосовидные, кистевидные, метельчатые или зонтиковидные общие соцветия, имеющие у своего основания, а часто и у основания своих ветвей влагалищеобразные верхушечные листья. Внешне колоски представителей этого семейства особенно похожи на колоски некоторых осоковых, например видов камыша (Scirpus). Соцветия с мужскими и соцветия с женскими цветками в пределах одного и того же вида обычно настолько отличаются по внешнему виду, что их нетрудно принять за принадлежащие разным видам и даже родам. Женские соцветия меньше разветвлены, число цветков в женских колосках обычно уменьшено, а у ряда родов (например, харперия — Harperia, гиполена — Hypolaena и др.) они имеют только один цветок. Каждый цветок в колоске располагается в пазухе перепончатой или кожистой кроющей чешуи — прицветника, а у двух родов — анартрии и лепиродии — по бокам цветков имеются еще по 2 прицветничка. Самые нижние в колоске кроющие чешуи обычно не несут в своих пазухах цветков.Околоцветник у рестиевых перепончатый или пленчатый, обычно состоящий из 4 или 6 расположенных 2 кругами бесцветных или слабоокрашенных сегментов более или менее одинакового строения. Сегменты внутреннего круга могут срастаться у своего основания. Кроме того, у некоторых видов один из сегментов, а иногда и все сегменты этого круга могут редуцироваться, и тогда околоцветник состоит из 3 или 5 сегментов. У австралийского рода локсокария (Loxocarya) околоцветник в женских цветках полностью отсутствует. У большинства родов семейства цветки актиноморфные или почти актиноморфные, однако у некоторых родов они становятся зигоморфными. Так, у капского тамнохортуса колосоносного (Thamnochortus spicigerus) боковые сегменты сросшегося при основании околоцветника сильно увеличены и снабжены килем, переходящим в широкий крыловидный вырост (рис. 188). У этого вида околоцветник сохраняется при плодах, играя роль летучки.Мужские цветки рестиевых имеют 3, реже 2 тычинки, расположенные супротивно внутренним сегментам околоцветника. Тычиночные нити обычно свободные, тонкие, с одногнездными, реже двугнездными (у примитивных и в других отношениях австралийских родов анартрии, лигинии — Lyginia и хопкинсии — Hopkinsia) пыльниками, прикрепленными к нити спинной стороной и вскрывающимися интрорзно поперечной щелью. Лишь у лигинии (рис. 188) нити тычинок срастаются в нижней части, образуя колонку. У этого последнего рода, как и у некоторых других родов, имеются надсвязники в виде шипиковидного выроста. Пыльцевые зерна рестиевых имеют однопоровую оболочку и обычно очень сходны с пыльцевыми зернами злаков.Синкарпный или паракарпный гинецей женских цветков рестиевых обычно состоит из 3 или 2 плодолистиков, лишь у родов тамнохортус и лепидоболус (Lepidobolus) он псевдомономерный в результате редукции переднего и одного из двух боковых гнезд первично-3-гнездной завязи. В двучленных женских цветках (например, у многочисленных видов рестио) два гнезда завязи располагаются супротивно двум наружным сегментам околоцветника. Ветви столбика обычно соответствуют числу гнезд завязи и в разной степени срастаются друг с другом. Волосистая рыльцевая поверхность находится на их внутренней стороне. Однако у некоторых родов гинецей имеет только один неразветвленный столбик, что связано или с редукцией двух гнезд завязи (у австралийского рода тамнохортус), или с полным срастанием всех ветвей столбика (например, у капской харперии). У гиподискуса (Hypodiscus) и вильденовии (Willdenowia) завязь, а позднее и плод располагаются на короткой дисковидной ножке, а у дильсии основание двухраздельного столбика пирамидообразно утолщено, как это бывает у некоторых родов осоковых. В каждом развитом гнезде гинецея имеется только один висячий ортотропный семязачаток.У большинства рестиевых плоды 2 — 3-семянные коробочки, раскрывающиеся продольной щелью по выступающим в виде ребер средним жилкам плодолистиков, т. е. локулицидно. Лишь немногие роды, например элегия и лепидоболус, имеют нераскрывающиеся орехообразные плоды с одним семенем. Семена всех рестиевых сходны по строению; они содержат обильный мучнистый эндосперм, у верхушки которого располагается маленький зародыш.Большинство рестиевых обитает на бедных питательными веществами, обычно кислых, песчаных или торфянистых почвах. Это растения преимущественно влажных открытых местообитаний, однако более или менее пересыхающих во время сухого периода года. Среди капских видов много обитателей междюнных западин на морских побережьях, а некоторые австралийские виды могут расти на солончаках и даже в условиях настоящей пустыни при почти полном отсутствии осадков. Однако ксероморфный облик свойствен всем рестиевым, даже растущим на непересыхающих болотах, что, по-видимому, объясняется историческими причинами (предки рестиевых могли расти в еще более влажных условиях) или физиологической сухостью местообитаний.Рестиевые являются ветроопыляемыми (анемофильными) растениями, причем возможность самоопыления у большинства видов исключается за счет двудомности — расположеиия мужских и женских цветков на разных особях. Специальных приспособлений для распространения семян у большинства рестиевых не отмечается. Выпадающие из коробочек семена могут переноситься ветром на некоторое расстояние от материнского растения, а затем разносятся или водными потоками, или на ногах животных вместе с комочками почвы. Лишь мелкие односемянные плоды рода тамнохортус, окутанные околоцветником с ширококрылатыми боковыми сегментами околоцветника, могут разноситься ветром на большие расстояния.Система рестиевых пока еще недостаточно разработана, хотя их обычно делят на две группы по числу гнезд в пыльниках: двугнезднопыльниковые (Diplantherae) — более примитивная группа и одногнезднопыльниковые (Haplantherae), к которым принадлежит большинство родов семейства. В последнее время рестиевые, исключая выделяемый в особое монотипное семейство род экдейоколея (Ecdeiocolea), делят на 2 подсемейства: собственно рестиевые (Restionoideae) и анартриевые (Anarthrioideae), к которому принадлежит только оригинальный австралийский род анартрия (рис. 188), обладающий рядом примитивных признаков (хорошо развитые листья, присутствие прицветничков и др.).Хозяйственное значение рестиевых незначительно. Их жесткие, безлистные или почти безлистные побеги не поедает или плохо поедает скот. Подобно тростнику, их можно использовать как покрытие для легких построек (особенно капский хондропеталум кровельный — Chondropetalum tectorum). В последнее время некоторые южноафриканские виды из родов элегия, хондропеталум, рестио и тамиохортус вводят в культуру в качестве декоративных растений, интересных своим своеобразным обликом.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.
.