- Род хвощ (Equisetum)
- Род хвощ (Equisetum)
-
Латинское название Equisetum впервые употребил для одного хвоща древнеримский естествоиспытатель Плиний Старший, имея в виду, очевидно, сходство ветвистых побегов хвоща с хвостом лошади (от лат. equius — лошадь и saeta, seta — щетина, жесткие волосы). Сходство взрослого растения с пучком волос, с хвостом отражено и в наиболее широко распространенном русском названии (хвощ) и в названиях на многих славянских языках. В некоторых районах нашей страны хвощи с мутовчато расположенными ветвями называли также «елка» или «сосенка», а по наличию у некоторых видов бурых спороносных побегов, похожих на пест,— пестиками или толкачиками. В природе хвощи обычно встречаются в виде клонов (группы растений, возникшие путем вегетативного размножения от одного экземпляра), которые нередко занимают участки площадью в несколько десятков и даже сотен квадратных метров.Размеры стебля хвощей сильно варьируют не только у разных видов, но даже в пределах одного достаточно обширного клона в зависимости от условий окружающей среды. Среди хвощей встречаются и небольшие, карликовые растения со стеблем высотой 5—15 см и диаметром 0,5—1 мм (хвощ камышковый — Equisetum scirpoides) и растения со стеблем длиной в несколько метров. Так, у хвоща многощетинкового (Е. myriochaetum) стебель достигает длины 9 м, но из-за малой толщины (0,5—2 см) такой длинный стебель не может самостоятельно пребывать в прямостоячем положении и вынужден опираться на соседние растения. Стебли большинства распространенных в умеренной зоне видов хвоща редко превышают высоты 1 м, у тропических видов стебли обычно более длинные.По характеру надземных побегов, а точнее, по их консистенции и функциональной морфологии виды хвоща можно разбить на две группы. У одних видов все надземные побеги однотипного строения. Они очень жесткие, обычно вечнозеленые (т. е. живут более одного сезона и в умеренной зоне перезимовывают) и развивают верхушечные стробилы (табл. 17). У видов второй группы побеги двух типов — одни спороносные, буроватые или зеленые, а другие вегетативные, зеленые. Побеги обоих типов появляются весной над поверхностью почвы, они нежнее по своей консистенции, чем побеги видов первой группы, и к зиме отмирают.
 Развитие вегетативных и спороносных побегов у видов второй группы идет по-разному. Например, у хвоща полевого (Е. arvense) спороносные побеги появляются первыми (ранней весной). Они буровато-розовые, неветвистые, обычно толще быстро зеленеющих вегетативных побегов и после спороношения большей частью отмирают. У хвощей лесного (Е. sylvaticum) и лугового (Е. pratense) спороносные побеги с хорошо развитыми стробилами появляются ранней весной одновременно с вегетативными. И те и другие вначале бледно-розовые, а затем зеленеют. После спороношения (и отсыхания стробила) спороносные побеги функционируют подобно вегетативным (табл. 17). Наконец, у таких видов, как хвощ болотный (Е. palustre), хвощ приречный (Е. fluviatile) и хвощ боготский (Е. bogotense), спороносные побеги в момент их появления над поверхностью земли с трудом можно отличить от вегетативных (такая же зеленая окраска и сходные размеры). Даже к моменту своего полного созревания спороносные побеги отличаются от вегетативных лишь наличием стробила (табл. 17).На конце растущего вегетативного побега хвоща под защитой молодых листовых влагалищ расположена верхушечная клетка, имеющая вид трехгранной пирамиды. Верхушечная клетка последовательно отделяет сегменты по трем граням, и в результате делений этих сегментов на стебле возникают кольцевидные зачатки влагалищ. Два — четыре слоя клеток в нижней части кольцевидного зачатка очень долго сохраняют свою способность к делению, образуя зону вставочной, или интеркалярной, меристемы, прикрытую снаружи листовым влагалищем ниже расположенного узла (если потянуть за верхушку растущего стебля хвоща, то разрыв происходит как раз по зоне вставочной меристемы). В результате удлинения производных интеркалярной меристемы происходит рост стебля хвоща в длину. Клетки вставочной меристемы, расположенные над каким-либо узлом, в конце концов теряют свою способность к делению и дифференцируются в постоянные ткани (именно поэтому зрелые, прекратившие рост стебли хвоща разрываются с большим трудом, чем молодые).На поверхности стебля хвоща в области междоузлия видны гребни и ложбинки, обычно чередующиеся с гребнями и ложбинками выше и ниже расположенных междоузлий. Число гребней обычно связано с диаметром стебля. На поперечном срезе междоузлия зрелого фотосинтезирующего стебля видны эпидерма, кора, проводящие пучки и центральная полость (рис. 78).
Развитие вегетативных и спороносных побегов у видов второй группы идет по-разному. Например, у хвоща полевого (Е. arvense) спороносные побеги появляются первыми (ранней весной). Они буровато-розовые, неветвистые, обычно толще быстро зеленеющих вегетативных побегов и после спороношения большей частью отмирают. У хвощей лесного (Е. sylvaticum) и лугового (Е. pratense) спороносные побеги с хорошо развитыми стробилами появляются ранней весной одновременно с вегетативными. И те и другие вначале бледно-розовые, а затем зеленеют. После спороношения (и отсыхания стробила) спороносные побеги функционируют подобно вегетативным (табл. 17). Наконец, у таких видов, как хвощ болотный (Е. palustre), хвощ приречный (Е. fluviatile) и хвощ боготский (Е. bogotense), спороносные побеги в момент их появления над поверхностью земли с трудом можно отличить от вегетативных (такая же зеленая окраска и сходные размеры). Даже к моменту своего полного созревания спороносные побеги отличаются от вегетативных лишь наличием стробила (табл. 17).На конце растущего вегетативного побега хвоща под защитой молодых листовых влагалищ расположена верхушечная клетка, имеющая вид трехгранной пирамиды. Верхушечная клетка последовательно отделяет сегменты по трем граням, и в результате делений этих сегментов на стебле возникают кольцевидные зачатки влагалищ. Два — четыре слоя клеток в нижней части кольцевидного зачатка очень долго сохраняют свою способность к делению, образуя зону вставочной, или интеркалярной, меристемы, прикрытую снаружи листовым влагалищем ниже расположенного узла (если потянуть за верхушку растущего стебля хвоща, то разрыв происходит как раз по зоне вставочной меристемы). В результате удлинения производных интеркалярной меристемы происходит рост стебля хвоща в длину. Клетки вставочной меристемы, расположенные над каким-либо узлом, в конце концов теряют свою способность к делению и дифференцируются в постоянные ткани (именно поэтому зрелые, прекратившие рост стебли хвоща разрываются с большим трудом, чем молодые).На поверхности стебля хвоща в области междоузлия видны гребни и ложбинки, обычно чередующиеся с гребнями и ложбинками выше и ниже расположенных междоузлий. Число гребней обычно связано с диаметром стебля. На поперечном срезе междоузлия зрелого фотосинтезирующего стебля видны эпидерма, кора, проводящие пучки и центральная полость (рис. 78).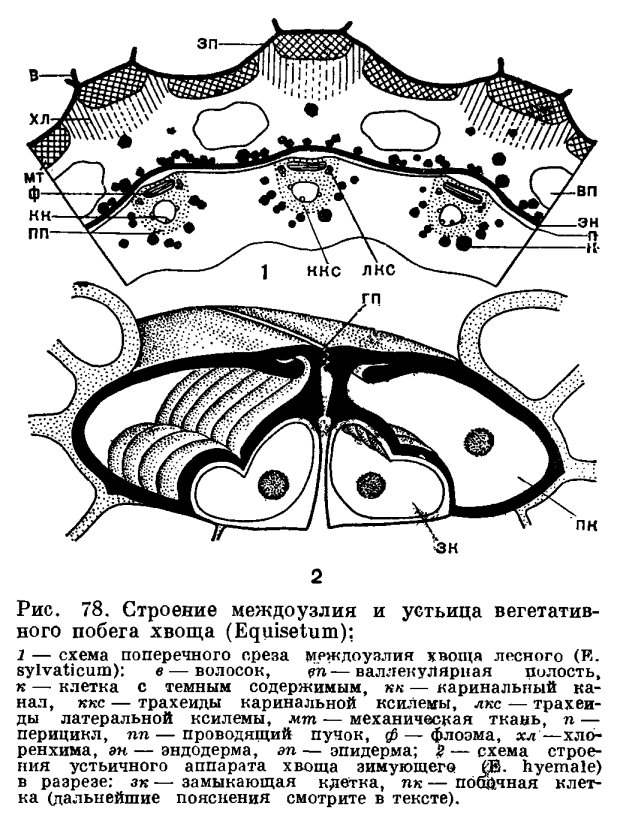 Эцидерма междоузлия состоит из плотно сомкнутых клеток с извилистайми смежными стенками. Внешняя оболочка зтих клеток сильно утолщена и снаружи покрыта шариками, стерженьками, сосочками, гребнями и другими скульптурными образованиями разной величины (рис. 79).
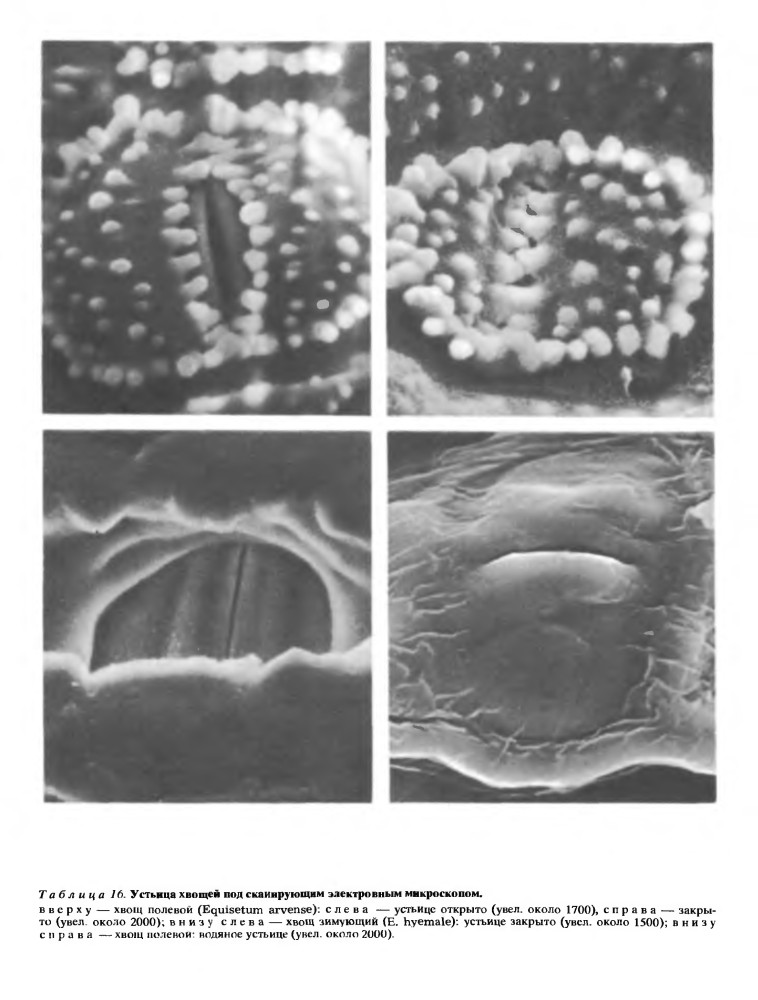
Эцидерма междоузлия состоит из плотно сомкнутых клеток с извилистайми смежными стенками. Внешняя оболочка зтих клеток сильно утолщена и снаружи покрыта шариками, стерженьками, сосочками, гребнями и другими скульптурными образованиями разной величины (рис. 79).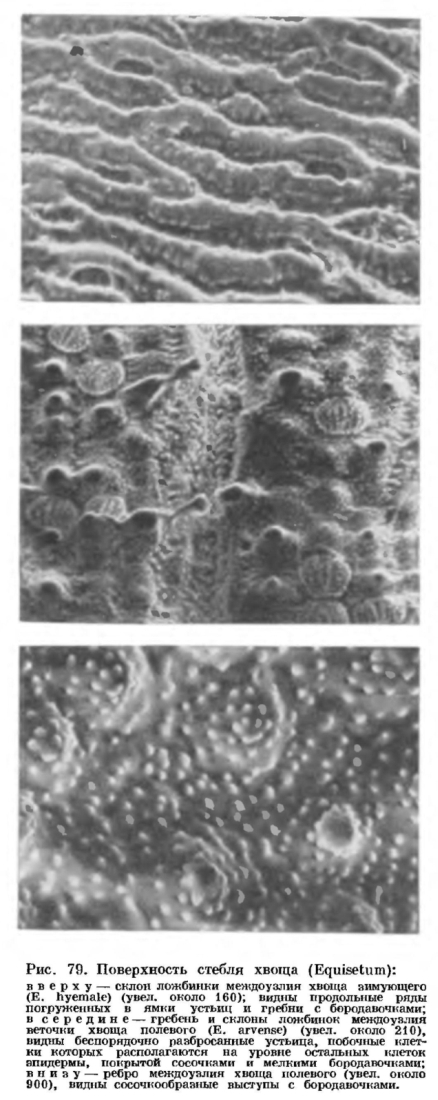 В состав оболочки клеток эпидермы и, в меньшей степени, других тканей побега входит наряду с целлюлозой кремнезем. Частицы кремнезема располагаются в клеточной оболочке между тяжами целлюлозы, а на поверхности клеток эпидермы образуют непрерывный и гомогенный, очень прочный слой с мелкими бугорками, приуроченными к выростам оболочки. Слой из кремнезема, в свою очередь, покрыт снаружи тонкой кутикулой с восковым налетом и играет не только механическую, но, по-видимому, и защитную роль, так как хвощи практически не подвергаются нападению различных растительноядных моллюсков и насекомых.Устьица в междоузлиях приурочены к тем участкам эпидермы, которые выстилают ложбинки. У одних видов хвоща устьица на каждом склоне ложбинки располагаются узкой полосой, в 1—4 четких продольных ряда, и погружены на дно воронковидной камеры, иногда общей для нескольких устьиц; у других видов устьичные полосы на склонах или дне ложбинок более широкие, без четко выраженных продольных рядов устьиц, а побочные клетки устьичного аппарата располагаются на уровне остальных клеток эпидермы (рис. 79, в середине). Число устьиц на поверхности стебля очень велико. Так, например, в одном устьичном ряду междоузлия хвоща зимующего (Е. hyemale) насчитывают около 400 устьиц, на поверхности одного междоузлия может быть уже около 15 тыс. устьиц, а на поверхности стебля высотой всего около полуметра — более 300 тыс. устьиц.Устьичный аппарат хвощей состоит из четырех клеток, расположенных попарно «в два зтажа»,— две побочные клетки лежат сверху и прикрывают две замыкающие клетки (рис. 78). Все четыре клетки возникают из одной материнской. На внутренней поверхности той стенки побочной клетки, которая граничит с замыкающей, есть гребневидные утолщения. Внешняя стенка побочной клетки снаружи гладкая или покрыта шариками из кремнезема, а изнутри несет сводообразное утолщение. Побочные клетки могут плотно замыкать устьичную щель благодаря наличию замкового механизма. У одних видов хвоща этот механизм построен по принципу гребень — паз (рис. 78), а у других—по принципу замка «молния» (кремниевые пупырышки или стерженьки, расположенные в один — три ряда по краю одной побочной клетки, входят в промежутки между пупырышками или стерженьками, расположенными по краю другой клетки, и наоборот, см. рис. 79, табл. 16).
В состав оболочки клеток эпидермы и, в меньшей степени, других тканей побега входит наряду с целлюлозой кремнезем. Частицы кремнезема располагаются в клеточной оболочке между тяжами целлюлозы, а на поверхности клеток эпидермы образуют непрерывный и гомогенный, очень прочный слой с мелкими бугорками, приуроченными к выростам оболочки. Слой из кремнезема, в свою очередь, покрыт снаружи тонкой кутикулой с восковым налетом и играет не только механическую, но, по-видимому, и защитную роль, так как хвощи практически не подвергаются нападению различных растительноядных моллюсков и насекомых.Устьица в междоузлиях приурочены к тем участкам эпидермы, которые выстилают ложбинки. У одних видов хвоща устьица на каждом склоне ложбинки располагаются узкой полосой, в 1—4 четких продольных ряда, и погружены на дно воронковидной камеры, иногда общей для нескольких устьиц; у других видов устьичные полосы на склонах или дне ложбинок более широкие, без четко выраженных продольных рядов устьиц, а побочные клетки устьичного аппарата располагаются на уровне остальных клеток эпидермы (рис. 79, в середине). Число устьиц на поверхности стебля очень велико. Так, например, в одном устьичном ряду междоузлия хвоща зимующего (Е. hyemale) насчитывают около 400 устьиц, на поверхности одного междоузлия может быть уже около 15 тыс. устьиц, а на поверхности стебля высотой всего около полуметра — более 300 тыс. устьиц.Устьичный аппарат хвощей состоит из четырех клеток, расположенных попарно «в два зтажа»,— две побочные клетки лежат сверху и прикрывают две замыкающие клетки (рис. 78). Все четыре клетки возникают из одной материнской. На внутренней поверхности той стенки побочной клетки, которая граничит с замыкающей, есть гребневидные утолщения. Внешняя стенка побочной клетки снаружи гладкая или покрыта шариками из кремнезема, а изнутри несет сводообразное утолщение. Побочные клетки могут плотно замыкать устьичную щель благодаря наличию замкового механизма. У одних видов хвоща этот механизм построен по принципу гребень — паз (рис. 78), а у других—по принципу замка «молния» (кремниевые пупырышки или стерженьки, расположенные в один — три ряда по краю одной побочной клетки, входят в промежутки между пупырышками или стерженьками, расположенными по краю другой клетки, и наоборот, см. рис. 79, табл. 16).
,
, Лежащая под эпидермой первичная кора в своей периферической части состоит из участков механической и фотосинтезирующей тканей. Взаимное расположение, величина и очертания этих участков на поперечном срезе междоузлия различны у разных видов (рис. 80), но всегда под выступами стебля, т. е. в гребнях, располагаются каринальные тяжи механической ткани. Механическая ткань сложена вытянутыми вдоль стебля узкими и длинными (до 0,5—1,5 мм) живыми клетками. Оболочка зрелых клеток содержит большое количество кремнезема, поэтому тяжи механической ткани совместно с эпидермой составляют главную механическую опору надземного стебля хвоща.
Лежащая под эпидермой первичная кора в своей периферической части состоит из участков механической и фотосинтезирующей тканей. Взаимное расположение, величина и очертания этих участков на поперечном срезе междоузлия различны у разных видов (рис. 80), но всегда под выступами стебля, т. е. в гребнях, располагаются каринальные тяжи механической ткани. Механическая ткань сложена вытянутыми вдоль стебля узкими и длинными (до 0,5—1,5 мм) живыми клетками. Оболочка зрелых клеток содержит большое количество кремнезема, поэтому тяжи механической ткани совместно с эпидермой составляют главную механическую опору надземного стебля хвоща.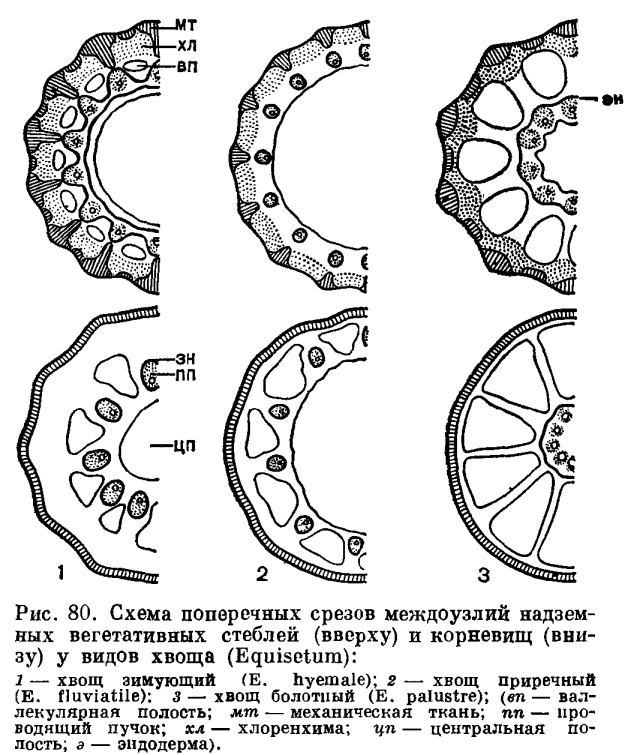 Фотосинтезирующая ткань, или хлоренхима стебля (обычные фотосинтезирующие органы, листья, у хвощей редуцированы и зту функцию выполняет стебель), подстилает в первую очередь те участки эпидермы, в которых находятся устьица, но хлоренхима может также находиться под гребнями или располагаться сплошным кольцом.Внутреннюю часть первичной коры составляет основная паренхима из тонкостенных, рыхло расположенных округлых клеток. В ней под ложбинками располагаются валлекулярные, или ложбиночные, полости, заполненные в начале своего развития водой, а затем воздухом.Зона проводящих пучков в междоузлии отграничена от окружающих тканей одним или двумя кольцами однослойной эндодермы, или каждый пучок окружен эндодермой (рис 80).
Фотосинтезирующая ткань, или хлоренхима стебля (обычные фотосинтезирующие органы, листья, у хвощей редуцированы и зту функцию выполняет стебель), подстилает в первую очередь те участки эпидермы, в которых находятся устьица, но хлоренхима может также находиться под гребнями или располагаться сплошным кольцом.Внутреннюю часть первичной коры составляет основная паренхима из тонкостенных, рыхло расположенных округлых клеток. В ней под ложбинками располагаются валлекулярные, или ложбиночные, полости, заполненные в начале своего развития водой, а затем воздухом.Зона проводящих пучков в междоузлии отграничена от окружающих тканей одним или двумя кольцами однослойной эндодермы, или каждый пучок окружен эндодермой (рис 80).
,Проводящая система хвоща представлена артростелой. На поперечном срезе междоузлия под гребнями видны закрытые (т. е. без камбия) коллатеральные пучки с каринальными каналами (от лат. carina — киль, здесь: гребень стебля), проводящими воду (рис. 78). Канал возникает вследствие растворения некоторых проводящих элементов ксилемы и расхождения окружающих эти элементы клеток паренхимы. Обычно у внутреннего края каринального канала, слегка вдаваясь в его полость, располагаются кольчатые трахеиды каринальной протоксилемы. Их очень немного, и каждая трахеида может быть равной по длине междоузлию. К каналу близ его наружного края примыкают трахеиды каринальной метаксилемы; одревесневшие кольца вторичной оболочки в них нередко соединены перемычками.Кнаружи от канала располагается флоэма, по бокам которой обычно лежат группы латеральной, или боковой, ксилемы. Иногда вследствие сильного удлинения междоузлия некоторые протоксилемные элементы латеральных групп разрушаются, и на их месте образуются полости, или лакуны, нередко почти такого же диаметра, что и каринальный канал. По этим лакунам, как и по каналу, идет восходящий ток воды с растворенными в ней веществами. Наличием трех независимых центров заложения ксилемы (каринального и двух боковых) проводящий пучок хвоща резко отличается от пучков других растений.Флоэма у хвощей состоит из ситовидных элементов и паренхимных клеток. Ситовидные элементы представляют собой узкие и длинные (иногда до 3 мм) клетки с небольшими ситовидными полями на продольных и конечных стенках. У большинства хвощей паренхимные клетки в центре междоузлия при росте стебля расходятся. При этом образуется полость, первоначально заполненная водой, а впоследствии воздухом. Эта полость вместе с ложбиночными полостями играет важную роль в газообмене зрелого растения с окружающей средой.Проводящий пучок междоузлия в средней части стебля, подходя к выше расположенному узлу, делится на три ветви, из которых средняя отклоняется наружу и становится пучком, идущим в лист (т. е. листовым следом) (рис. 81). Боковые ветви, сливаясь с боковыми же ветвями соседних пучков, образуют так называемые синтетические пучки, проходящие далее в следующее междоузлие. На поперечном и продольном срезах вегетативного стебля в области узла (рис. 81) видно, что ложбиночные и центральные полости соседних междоузлий разделены перегородками, а проводящая система после от хождения листовых следов имеет вид короткой трубочки, или сифоностелы. Листовые прорывы в стеле при отхождении листовых следов не образуются.
На поперечном и продольном срезах вегетативного стебля в области узла (рис. 81) видно, что ложбиночные и центральные полости соседних междоузлий разделены перегородками, а проводящая система после от хождения листовых следов имеет вид короткой трубочки, или сифоностелы. Листовые прорывы в стеле при отхождении листовых следов не образуются.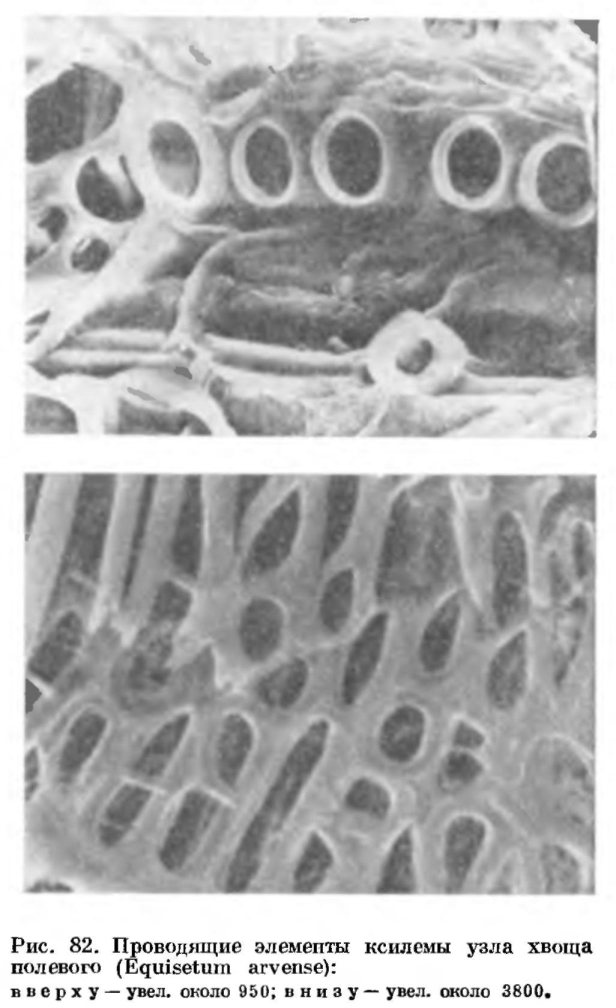 В узлах по сравнению с междоузлиями сильно возрастает объем метаксилемы, которая здесь состоит из очень коротких или почти изодиаметрических сетчатых или пористых трахеид с простыми или окаймленными округлыми порами (рис. 82). В состав ксилемы узла у хвощей входят также своеобразные сосуды, состоящие обычно из двух члеников.В диафрагмах (перегородках) между центральными полостями соседних междоузлий у молодого стебля имеются отверстия, по которым воздух из одной центральной полости стебля может переходить в другую.Листовые влагалища, охватывающие основание каждого междоузлия (рис. 83) и защищающие зоны вставочной меристемы, по своему внутреннему строению близки к периферической части нижележащих междоузлий, а гребни и устьичные ряды этих междоузлий продолжают гребни и устьичные ряды влагалищ и прослеживаются далее в листовых пластинках. Эпидерма примыкающей к стеблю внутренней поверхности влагалища у большинства видов обычно не имеет нормальных (вентиляционных) устьиц.
В узлах по сравнению с междоузлиями сильно возрастает объем метаксилемы, которая здесь состоит из очень коротких или почти изодиаметрических сетчатых или пористых трахеид с простыми или окаймленными округлыми порами (рис. 82). В состав ксилемы узла у хвощей входят также своеобразные сосуды, состоящие обычно из двух члеников.В диафрагмах (перегородках) между центральными полостями соседних междоузлий у молодого стебля имеются отверстия, по которым воздух из одной центральной полости стебля может переходить в другую.Листовые влагалища, охватывающие основание каждого междоузлия (рис. 83) и защищающие зоны вставочной меристемы, по своему внутреннему строению близки к периферической части нижележащих междоузлий, а гребни и устьичные ряды этих междоузлий продолжают гребни и устьичные ряды влагалищ и прослеживаются далее в листовых пластинках. Эпидерма примыкающей к стеблю внутренней поверхности влагалища у большинства видов обычно не имеет нормальных (вентиляционных) устьиц. По верхнему краю влагалища сидят зубцевидные листовые пластинки, которые в почке сращены между собой и образуют колпачок, прикрывающий верхушку стебля с более молодыми зачатками листьев. В дальнейшем, при росте стебля в длину и толщину, листовые пластинки отходят друг от друга, но разрыв происходит иногда не по всем линиям срастания, поэтому число зубцов по краю влагалища может не соответствовать действительному числу листьев.Зубцевидные листовые пластинки обычно короткие и содержат очень мало хлоренхимы. Форма влагалища, цвет, форма и время жизни зубцов различны у разных видов (рис. 83), что и используется при определении хвощей. На внутренней, обращенной к стеблю поверхности пластинки, а иногда и влагалища располагается структура, выделяющая капельно-жидкую воду и называемая гидатодой. С поверхности она выглядит как светлый участок листа, на котором располагаются водяные устьица.
По верхнему краю влагалища сидят зубцевидные листовые пластинки, которые в почке сращены между собой и образуют колпачок, прикрывающий верхушку стебля с более молодыми зачатками листьев. В дальнейшем, при росте стебля в длину и толщину, листовые пластинки отходят друг от друга, но разрыв происходит иногда не по всем линиям срастания, поэтому число зубцов по краю влагалища может не соответствовать действительному числу листьев.Зубцевидные листовые пластинки обычно короткие и содержат очень мало хлоренхимы. Форма влагалища, цвет, форма и время жизни зубцов различны у разных видов (рис. 83), что и используется при определении хвощей. На внутренней, обращенной к стеблю поверхности пластинки, а иногда и влагалища располагается структура, выделяющая капельно-жидкую воду и называемая гидатодой. С поверхности она выглядит как светлый участок листа, на котором располагаются водяные устьица.
,Водяные устьица отличаются от обычных (вентиляционных) как своим расположением, так и строением (табл. 16). Они открыты, располагаются над проводящим пучком, а побочные клетки устьичного аппарата всегда лежат на уровне остальных клеток эпидермы. Замыкающие клетки лишены хлоропластов и имеют равпомерно утолщенные оболочки. Под эпидермой в области гидатоды располагается не фотосинтезирующая ткань, как под обычными устьицами, а особая тонкостенная паренхимная ткань, прикрывающая жилку. Вода, поступающая по трахеидам жилки, через эту ткань попадает к водяным устьицам и выделяется наружу. Выделение капельножидкой воды, или гуттацию, можно наблюдать у хвощей таежной зоны с позднего вечера до раннего утра летом в ясную погоду, когда количество воды, нагнетаемой корнями из теплой, прогретой за день почвы, превышает количество воды, испаряемой побегом. Капли воды в это время хорошо заметны по краю влагалищ, особенно влагалищ последних узлов (табл. 17).Стебель хвощей ветвится моноподиально. Зачатки боковых ветвей (почки) первоначально прикрыты основаниями листовых влагалищ, но при своем дальнейшем развитии прорывают их (рис. 81, 83). Некоторые зачатки остаются спящими, так что на одном побеге можно обнаружить узлы с ветвями и без ветвей. Точно так же в пределах одного клона можно встретить побеги с ветвями и без ветвей. Обычно считают, что хвощи резко отличаются от других высших растений особым типом заложения боковых ветвей, зачатки которых возникают не в пазухах листьев, а чередуются с листьями. Однако ветви хвоща можно считать пазушными, если допустить, что в процессе эволюции большая часть листовой пластинки, в пазухе которой закладывается ветвь, приросла на протяжении целого междоузлия к стеблю (рис. 81). Тогда кончик листа, «родительского» по отношению к какой-либо ветви, следует искать не у того же узла, от которого отходит данная ветвь, а у выше расположенного соседнего узла. Допуская прирастание листа к стеблю, легко объяснить и наличие трех групп ксилемы в пучке междоузлия хвоща.
,У основания боковой почки образуется зачаток корня. Изучение развития почки показывает, что почка и корень происходят от одной материнской клетки, при делении которой возникает более или менее шаровидная масса клеток. В верхней части клеточной массы дифференцируется побег, а в нижней — корень. Зачатки корней у надземных побегов обычно не прорастают, но если побег присыпать землей или погрузить в воду, на его узлах из зачатков корней вырастут корни, хотя некоторые почки останутся спящими.Особое значение в возобновлении надземных побегов и в вегетативном размножении хвощей имеют спящие почки тех узлов стебля, которые находятся под землей близ ее поверхности.У хвощей, которые характеризуются диморфизмом побегов, наблюдаются некоторые особенности в строении стеблей спороносных побегов. Так, стебли спороносных побегов хвоща полевого в начале спороношения напоминают по своему строению незеленые, еще только начинающие показываться из земли стебли вегетативных побегов; они округлы в поперечном сечении, содержат мизерное количество кремнезема, не имеют ложбиночных полостей, хлоренхимы и устьиц (устьица есть лишь на листовых влагалищах), а под эпидермой у них сплошным кольцом располагается ткань, клетки которой можно рассматривать как находящиеся на первых этапах онтогенеза клетки механиче ской ткани, свойственной зрелым вегетативным побегам. По мере созревания спороносных побегов строение их претерпевает незначительные изменения; в них появляются, например, ложбиночные полости. Но обычно в спороносном побеге не возникают хлоренхима и хорошо развитая механическая ткань, столь характерные для зрелого вегетативного стебля.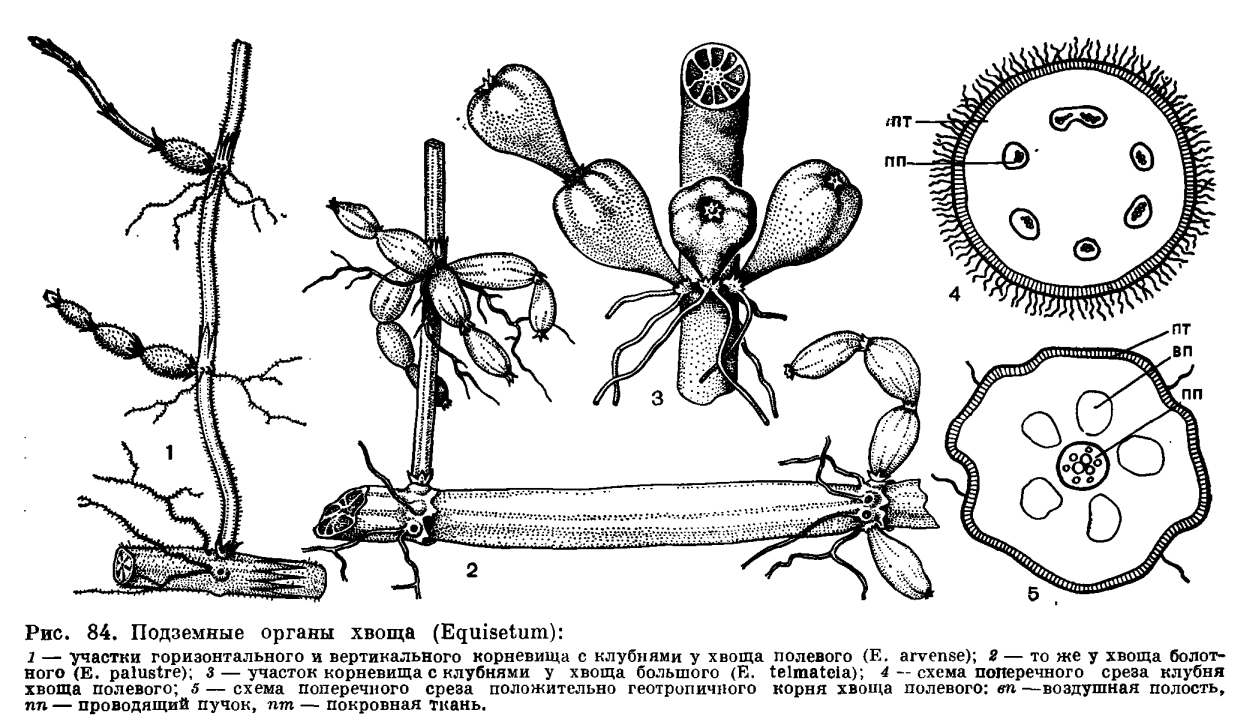 Как уже отмечалось, хвощи являются многолетними корневищными растениями. Их корневища расположены в почве на разной глубине, а размеры корневищ варьируют у разных видов и у растений одного и того же вида. У хвощей есть два типа корневищ — горизонтальные и вертикальные. Горизонтальные корневища обычно более толстые, с более длинными междоузлиями; вертикальные — более тонкие и с более короткими междоузлиями (рис. 84). Например, у хвоща болотного толщина горизонтального корневища равна 5—10 мм при длине междоузлий до 25 см, а толщина вертикального — 1—3 мм при длине междоузлий до 10 см. С помощью ветвящихся горизонтальных корневищ, расположенных часто на глубине до 0,5—2 м, хвощ как бы захватывает новые территории, а с помощью вертикальных — осваивает их. Истинными корневищами следует считать именно горизонтальные — это органы вегетативного размножения. Вертикальные корневища являются, по сути дела, слегка видоизмененными подземными основаниями надземных побегов, отходящих от горизонтальных корневищ.Поверхность междоузлий корневищ матовая или блестящая, по сравнению с поверхностью надземных междоузлий вегетативных побегов более ровная, гребни выражены слабее или вовсе отсутствуют, цвет корневищ варьирует от светло-желтого или розоватого до почти черного. Как и на надземных стеблях, на корневищах имеются листовые влагалища, но они здесь (а часто и поверхность междоузлия) нередко покрыты волосками. Молодые волоски выделяют слизь, которая, очевидно, защищает молодые побеги и почки от иссушения. Со временем оболочки волосков, как и остальных клеток эпидермы, пропитываются жироподобными веществами и становятся непроницаемыми для воды и газов. Тем самым испарение воды с поверхности корневища резко уменьшается.У корневищ нет устьиц, хлоренхимы и тяжей механической ткани такого типа, что находятся в надземных вегетативных стеблях. Под эпидермой междоузлия корневища залегают один — пять слоев более или менее толстостенных, не одревесневших, но пропитанных жироподобными веществами и кремнеземом паренхимных клеток, под которыми, в свою очередь, лежат тонкостенные клетки основной паренхимы, содержащие крахмальные зерна. Количество крахмала в клетках зависит от сезона года и от местоположения данного участка корневища. Например, у хвоща болотного количество крахмала в корневище уменьшается в начале лета (в связи с развитием надземных побегов), затем возрастает в середине лета (когда надземные побеги активно фотосинтезируют) и вновь падает к осени (в связи с образованием зачатков побегов будущего года).Как и на надземных стеблях, на корневищах под защитой листовых влагалищ закладываются зачатки побегов и корней, но, в отличие от надземных стеблей, большинство почек на корневищах обычно остаются спящими, а зачатки корней в основании этих почек прорастают и пробивают влагалища, так что узлы корневища несут не мутовки ветвей, а мутовки корней (рис. 84).Некоторые боковые почки как на горизонтальных, так и на вертикальных корневищах, прорастая, образуют клубень, который представляет собой сильно утолщенное и видоизмененное укороченное междоузлие ветви. Иногда образуется цепочка таких клубней — соседних междоузлий одной ветви (рис. 84). Величина клубней варьирует у разных видов; наиболее крупные клубни (30 х 25 мм), по-видимому, свойственны хвощу большому (Е. telmateia). Клетки паренхимы клубня очень крупные и буквально забиты крахмальными зернами.У хвощей встречаются корни двух типов: корни, обладающие положительным геотропизмом, т. е. растущие вниз, по направлению силы тяжести, и корни агеотропичные, т. е. не реагирующие на силу тяжести. Положительно геотропичные корни толстые, диаметром 1 — 5 мм. Они обычно отходят по одному от узлов корневища и часто достигают длины 0,5—2 м, так что всасывающая зона корней находится в обводненных горизонтах почвы даже у видов, обитающих в сухих местах. Агеотропичные корни тонкие, волосовидные, диаметром менее 1 мм; они располагаются мутовками на узлах корневища, а в длину обычно не превышают 10 см. В корнях хвощей имеются воздухоносные полости и сосуды, членики которых обладают простой перфорационной пластинкой (рис. 85). По сосудам вода быстро подается из глубоких горизонтов почвы к корневищам и надземным побегам.
Как уже отмечалось, хвощи являются многолетними корневищными растениями. Их корневища расположены в почве на разной глубине, а размеры корневищ варьируют у разных видов и у растений одного и того же вида. У хвощей есть два типа корневищ — горизонтальные и вертикальные. Горизонтальные корневища обычно более толстые, с более длинными междоузлиями; вертикальные — более тонкие и с более короткими междоузлиями (рис. 84). Например, у хвоща болотного толщина горизонтального корневища равна 5—10 мм при длине междоузлий до 25 см, а толщина вертикального — 1—3 мм при длине междоузлий до 10 см. С помощью ветвящихся горизонтальных корневищ, расположенных часто на глубине до 0,5—2 м, хвощ как бы захватывает новые территории, а с помощью вертикальных — осваивает их. Истинными корневищами следует считать именно горизонтальные — это органы вегетативного размножения. Вертикальные корневища являются, по сути дела, слегка видоизмененными подземными основаниями надземных побегов, отходящих от горизонтальных корневищ.Поверхность междоузлий корневищ матовая или блестящая, по сравнению с поверхностью надземных междоузлий вегетативных побегов более ровная, гребни выражены слабее или вовсе отсутствуют, цвет корневищ варьирует от светло-желтого или розоватого до почти черного. Как и на надземных стеблях, на корневищах имеются листовые влагалища, но они здесь (а часто и поверхность междоузлия) нередко покрыты волосками. Молодые волоски выделяют слизь, которая, очевидно, защищает молодые побеги и почки от иссушения. Со временем оболочки волосков, как и остальных клеток эпидермы, пропитываются жироподобными веществами и становятся непроницаемыми для воды и газов. Тем самым испарение воды с поверхности корневища резко уменьшается.У корневищ нет устьиц, хлоренхимы и тяжей механической ткани такого типа, что находятся в надземных вегетативных стеблях. Под эпидермой междоузлия корневища залегают один — пять слоев более или менее толстостенных, не одревесневших, но пропитанных жироподобными веществами и кремнеземом паренхимных клеток, под которыми, в свою очередь, лежат тонкостенные клетки основной паренхимы, содержащие крахмальные зерна. Количество крахмала в клетках зависит от сезона года и от местоположения данного участка корневища. Например, у хвоща болотного количество крахмала в корневище уменьшается в начале лета (в связи с развитием надземных побегов), затем возрастает в середине лета (когда надземные побеги активно фотосинтезируют) и вновь падает к осени (в связи с образованием зачатков побегов будущего года).Как и на надземных стеблях, на корневищах под защитой листовых влагалищ закладываются зачатки побегов и корней, но, в отличие от надземных стеблей, большинство почек на корневищах обычно остаются спящими, а зачатки корней в основании этих почек прорастают и пробивают влагалища, так что узлы корневища несут не мутовки ветвей, а мутовки корней (рис. 84).Некоторые боковые почки как на горизонтальных, так и на вертикальных корневищах, прорастая, образуют клубень, который представляет собой сильно утолщенное и видоизмененное укороченное междоузлие ветви. Иногда образуется цепочка таких клубней — соседних междоузлий одной ветви (рис. 84). Величина клубней варьирует у разных видов; наиболее крупные клубни (30 х 25 мм), по-видимому, свойственны хвощу большому (Е. telmateia). Клетки паренхимы клубня очень крупные и буквально забиты крахмальными зернами.У хвощей встречаются корни двух типов: корни, обладающие положительным геотропизмом, т. е. растущие вниз, по направлению силы тяжести, и корни агеотропичные, т. е. не реагирующие на силу тяжести. Положительно геотропичные корни толстые, диаметром 1 — 5 мм. Они обычно отходят по одному от узлов корневища и часто достигают длины 0,5—2 м, так что всасывающая зона корней находится в обводненных горизонтах почвы даже у видов, обитающих в сухих местах. Агеотропичные корни тонкие, волосовидные, диаметром менее 1 мм; они располагаются мутовками на узлах корневища, а в длину обычно не превышают 10 см. В корнях хвощей имеются воздухоносные полости и сосуды, членики которых обладают простой перфорационной пластинкой (рис. 85). По сосудам вода быстро подается из глубоких горизонтов почвы к корневищам и надземным побегам.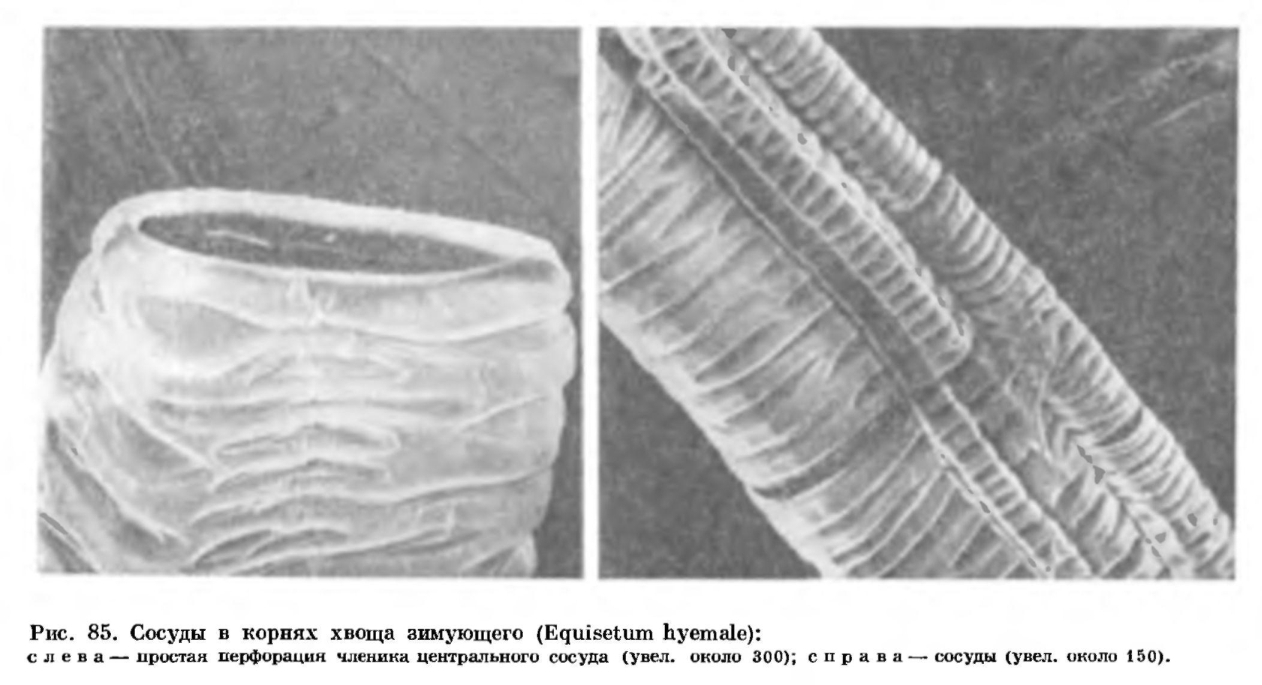 Вегетативное размножение хвощей осуществляется прежде всего с помощью корневищ. Старые участки корневищ отмирают, и первоначально единый материнский клон распадается на несколько производных.Стробилы хвощей возникают обычно на концах стеблей, реже боковых ветвей. Число стробилов на одном побеге варьирует от одного (у большинства видов) до сотни (у хвоща многощетинкового). Стробилы более или менее эллипсоидальные, от 2 до 80 мм в длину, от желтоватых до бурых или почти черных, тупые или острые (рис. 86). В основании стробила находится воротничок, представляющий собой редуцированное листовое влагалище, а на оси стробила мутовками располагаются спорангиофоры в виде шестиугольных (обычно) щитков на ножках.
Вегетативное размножение хвощей осуществляется прежде всего с помощью корневищ. Старые участки корневищ отмирают, и первоначально единый материнский клон распадается на несколько производных.Стробилы хвощей возникают обычно на концах стеблей, реже боковых ветвей. Число стробилов на одном побеге варьирует от одного (у большинства видов) до сотни (у хвоща многощетинкового). Стробилы более или менее эллипсоидальные, от 2 до 80 мм в длину, от желтоватых до бурых или почти черных, тупые или острые (рис. 86). В основании стробила находится воротничок, представляющий собой редуцированное листовое влагалище, а на оси стробила мутовками располагаются спорангиофоры в виде шестиугольных (обычно) щитков на ножках.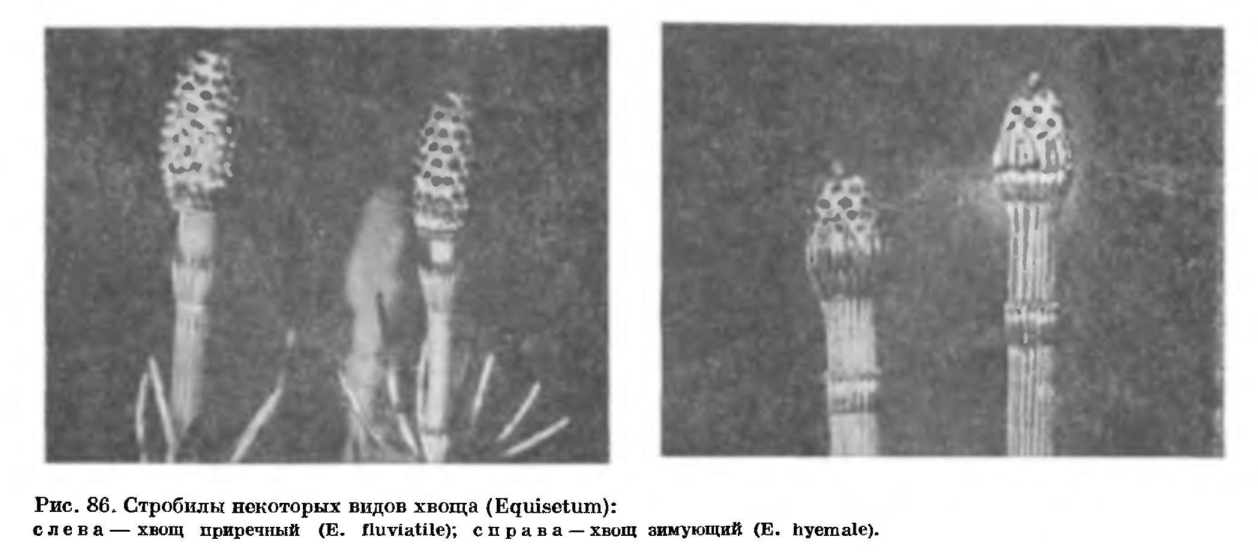 На внутренней стороне щитка располагаются 4—16 вытянутых вдоль ножки спорангиев.Спорангий хвоща развивается из группы поверхностных клеток: одной более крупной (осевой клетки) и нескольких более мелких. Каждая из этих клеток делится перегородкой, проходящей параллельно поверхности спорангиофора. Из внутренних клеток затем образуется спорогенная ткань, а из внешних — спорогенная ткань и многослойная степка спорангия. Клетки, прилегающие к спорогенной ткани, впоследствии дифференцируются в выстилающий слой, или тапетум. Оболочки клеток тапетума и некоторых снорогенных клеток расплываются, и образуется масса цитоплазмы с ядрами, или периплазмодий, который идет на питание развивающихся спор. Большая часть спорогенных клеток, делясь митотически, дает начало материнским клеткам спор, или спороцитам, каждый из которых после мейоза образует тетраду спор. У хвощей, обитающих в умеренной зоне, мейоз в спорангиях протекает летом, или в конце периода роста, а спороношение наступает в то же лето или весной следующего года. У многих хвощей тропической зоны нет периодичности в образовании стробилов и приуроченности мейоза к какомулибо сезону года, так что на одном побеге можно встретить стробилы, находящиеся на разных стадиях развития.Споры хвощей сфероидальные, диаметром 30—80 мкм, причем у каждого вида размер спор представляет собой более или менее постоянную величину. В цитоплазме спор находится крупное ядро, вокруг которого равномерно распределены многочисленные хлоропласты с гранами крахмала. Из внешнего слоя оболочки споры при ее созревании возникают спирально обернутые вокруг тела споры гигроскопические ленты — элатеры (рис. 87).
На внутренней стороне щитка располагаются 4—16 вытянутых вдоль ножки спорангиев.Спорангий хвоща развивается из группы поверхностных клеток: одной более крупной (осевой клетки) и нескольких более мелких. Каждая из этих клеток делится перегородкой, проходящей параллельно поверхности спорангиофора. Из внутренних клеток затем образуется спорогенная ткань, а из внешних — спорогенная ткань и многослойная степка спорангия. Клетки, прилегающие к спорогенной ткани, впоследствии дифференцируются в выстилающий слой, или тапетум. Оболочки клеток тапетума и некоторых снорогенных клеток расплываются, и образуется масса цитоплазмы с ядрами, или периплазмодий, который идет на питание развивающихся спор. Большая часть спорогенных клеток, делясь митотически, дает начало материнским клеткам спор, или спороцитам, каждый из которых после мейоза образует тетраду спор. У хвощей, обитающих в умеренной зоне, мейоз в спорангиях протекает летом, или в конце периода роста, а спороношение наступает в то же лето или весной следующего года. У многих хвощей тропической зоны нет периодичности в образовании стробилов и приуроченности мейоза к какомулибо сезону года, так что на одном побеге можно встретить стробилы, находящиеся на разных стадиях развития.Споры хвощей сфероидальные, диаметром 30—80 мкм, причем у каждого вида размер спор представляет собой более или менее постоянную величину. В цитоплазме спор находится крупное ядро, вокруг которого равномерно распределены многочисленные хлоропласты с гранами крахмала. Из внешнего слоя оболочки споры при ее созревании возникают спирально обернутые вокруг тела споры гигроскопические ленты — элатеры (рис. 87).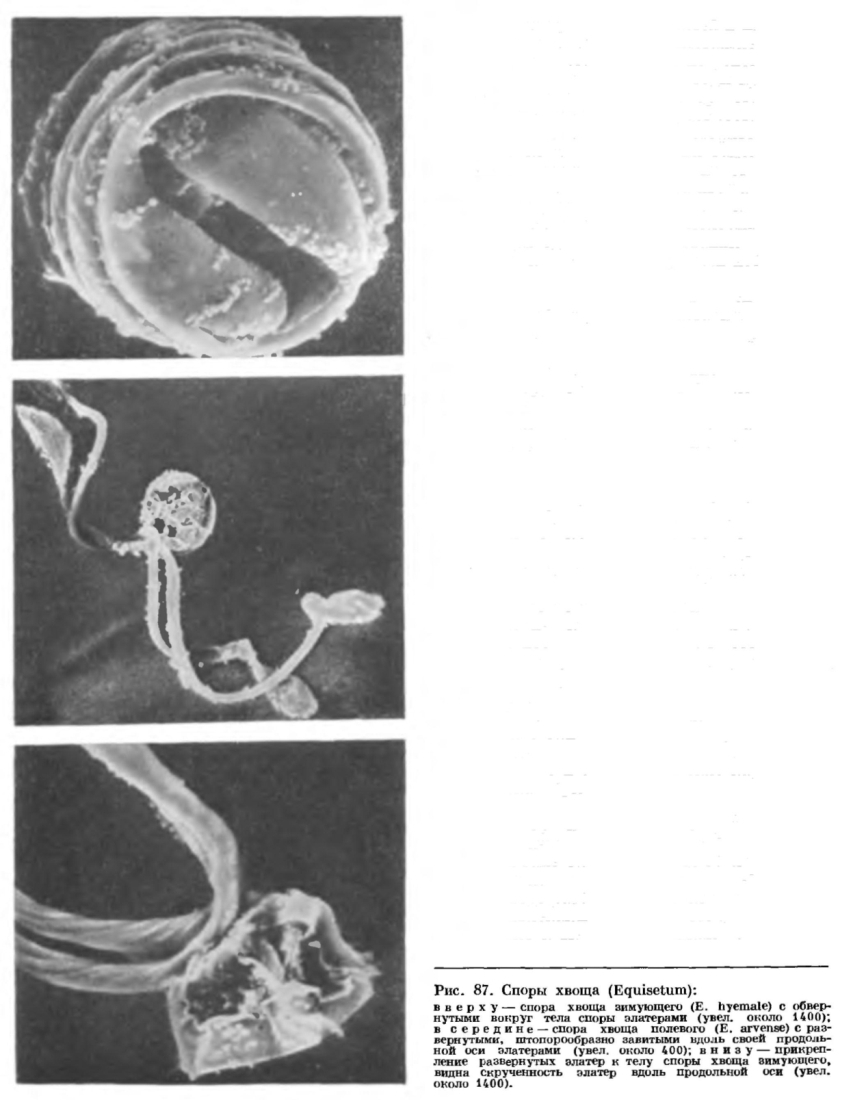 По созревании спор ось стробила слегка вытягивается, щитки спорангиофоров отходят один от другого, а спорангии быстро подсыхают. К этому моменту внутренние слои стенки спорангия уже разрушились, и остался лишь один внешний слой, называемый экзотецием, при подсыхании клеток которого стенка спорангия лопается продольной щелью. Вскрыванию спорангия способствует возрастающее вследствие развертывания и скручивания элатер при подсыхании давление споровой массы на стенку спорангия изнутри. После вскрывания спорангия элатеры еще более развертываются, разрыхляя массу спор и одновременно сцепляя их друг с другом, так что в конечном счете выходящее через щель в стенке содержимое спорангия принимает вид серовато-зеленого рыхлого порошка, комки которого легко сдуваются ветром и переносятся на значительные расстояния. Споры хвоща быстро погибают во внешней среде и прорастают лишь те, которые вскоре после выпадения из спорангия попадают на затененную влажную поверхность почвы или в воду. Попав в благоприятные условия, споры очень быстро прорастают. При этом спора сильно набухает, так называемый срединный, в действительности второй, считая снаружи, слой обо лочки споры лопается и сбрасывается (первый слой — элатеры — в природе нередко сбрасывается еще раньше, при ударе переносимых ветром спор о какое-либо препятствие или в момент резких гигроскопических движений).
По созревании спор ось стробила слегка вытягивается, щитки спорангиофоров отходят один от другого, а спорангии быстро подсыхают. К этому моменту внутренние слои стенки спорангия уже разрушились, и остался лишь один внешний слой, называемый экзотецием, при подсыхании клеток которого стенка спорангия лопается продольной щелью. Вскрыванию спорангия способствует возрастающее вследствие развертывания и скручивания элатер при подсыхании давление споровой массы на стенку спорангия изнутри. После вскрывания спорангия элатеры еще более развертываются, разрыхляя массу спор и одновременно сцепляя их друг с другом, так что в конечном счете выходящее через щель в стенке содержимое спорангия принимает вид серовато-зеленого рыхлого порошка, комки которого легко сдуваются ветром и переносятся на значительные расстояния. Споры хвоща быстро погибают во внешней среде и прорастают лишь те, которые вскоре после выпадения из спорангия попадают на затененную влажную поверхность почвы или в воду. Попав в благоприятные условия, споры очень быстро прорастают. При этом спора сильно набухает, так называемый срединный, в действительности второй, считая снаружи, слой обо лочки споры лопается и сбрасывается (первый слой — элатеры — в природе нередко сбрасывается еще раньше, при ударе переносимых ветром спор о какое-либо препятствие или в момент резких гигроскопических движений).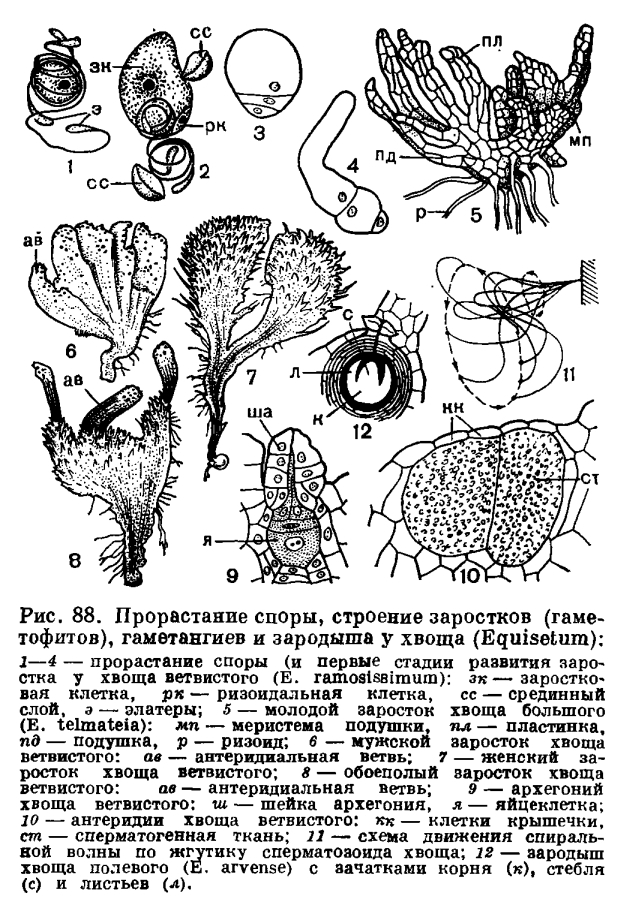 Из споры после первого деления образуются положительно геотропичный ризоид и заростковая клетка (клетка гаметофита), которая многократно делится и дает начало зеленому гаметофиту (рис. 88). При слабом освещении или в загущенных посевах гаметофит имеет вид однорядной зеленой нити — хлоронемы. При достаточном освещении нитевидная стадия в развитии гаметофита обычно не выражена, и он сразу принимает вид однослойной пластинки, нарастающей с помощью верхушечной меристемы. Некоторые клетки пластинчатого гаметофита дают начало вторичным ризоидам. Спустя некоторое время двухмерный пластинчатый гаметофит превращается в трехмерный гаметофит с более или менее массивной распростертой по субстрату базальной частью — подушкой. На верхней стороне подушки развиваются прямостоячие зеленые пластинки длиной до 1—4 мм, а на нижней — располагаются бесцветные, длиной до 1 см ризоиды, прикрепляющие гаметофит к почве и высасывающие из нее воду с минеральными солями. При хороших условиях освещения и питания по краю подушки вскоре дифференцируется меристема, в результате деятельности которой увеличивается как объем подушки, так и число пластинок и ризоидов.При проращивании одиночных спор на стерильной питательной среде удалось выяснить, что у хвощей имеется три типа гаметофитов: мужские, женские и обоеполые, различающиеся между собой по скорости роста, темпам развития и морфологии. Размеры мужских гаметофитов у хвощей варьируют в диаметре от 1 до 10 мм, женских — от 3 до 30 мм. У одного и того же вида мужские гаметофиты при равных условиях культуры мельче женских. Первые антеридии появляются на мужских гаметофитах в зависимости от вида хвоща спустя 20 — 110 дней после посева спор, а первые архегонии на женских гаметофитах — спустя 30 — 130 дней.При формировании мужского гаметофита краевая меристема образует небольшое число пластинок, а затем дает начало антеридиальной ветви — резко изогнутой кверху лопасти, на нижней стороне которой по краю возникают антеридии.У гаметофитов, находимых в природе, число антеридиев обычно невелико. В культуре старые гаметофиты могут иметь несколько ветвей с десятками и сотнями антеридиев. Антеридии могут возникать и на кончиках пластинок, а иногда пластинки прорастают и дают начало более или менее массивному телу с краевой меристемой. Эта краевая меристема или может дать начало новым мужским ветвям с собственной меристемой, или из нее у немногих видов образуется подушка с пластинками и с архегониями, и, таким образом, первоначально мужской гаметофит превращается в обоеполый.При формировании женского гаметофита краевая меристема обычно не прекращает своей деятельности по образованию подушки, пластинок и ризоидов, но, когда подушка достигнет определенной величины, из некоторых клеток меристемы возникают архегонии. Зрелые архегонии располагаются у основания пластинок. У неветвистых гаметофитов спустя некоторое время меристема по всему краю подушки начинает формировать антеридии. Если гаметофиты ветвистые, то краевая меристема только некоторых ветвей перерождается и дает начало антеридиальным ветвям с антеридиями, в основании которых затем возникает антеридиальная меристема. Таким образом, первоначально женский гаметофит превращается в обоеполый, несущий одновременно и архегонии и антеридии. Постепенно все ветви женского гаметофита перерождаются в антеридиальные, и после отмирания архегониев гаметофит становится функционально мужским.В культуре число архегониев на одном гаметофите варьирует от 20 до нескольких сотен; у гаметофитов, находимых в природе, число архегониев нередко много меньше.Следует отметить, что женские гаметофиты всех видов хвоща могут стать обоеполыми, и это резко контрастирует с неспособностью мужских гаметофитов подавляющего большинства представителей этого рода формировать архегонии.Гаметофиты хвощей очень чувствительны к воздействию окружающих условий. В опыте со строго контролируемыми и постоянными факторами внешней среды нередко можно заметить различия в морфологии гаметофитов разных видов и подродов. У гаметофитов, выросших в природе, видовые различия подметить трудно или вовсе не удается, да и сами дикорастущие гаметофиты иногда очень резко отличаются от культивируемых по величине подушки, числу и длине лопастей и пластинок, скорости развития, числу гаметангиев и т. д. В природе гаметофиты могут не успеть до наступления неблагоприятной погоды приступить к формированию гаметангиев другого пола, и популяция, таким образом, будет состоять только из мужских и женских гаметофитов.Обычно количество мужских гаметофитов в популяции очень сильно варьирует (от 3 до 100%) в зависимости и от вида хвоща, и от разнообразных условий среды. В целом условия, благоприятствующие росту молодых гаметофитов, приводят к формированию большего числа женских гаметофитов, а при неблагоприятных условиях роста образуется больше мужских. Из долго хранившихся спор, обладающих малой энергией прорастания, возникает больше мужских гаметофитов, чем при тех же условиях из свежесобранных спор.Зрелые антеридии разных видов хвоща отличаются друг от друга формой полости, в которой лежат сперматоциты, степенью своего возвышения над поверхностью гаметофита, числом, формой и расположением клеток крышечки.Хвощи обладают крупными и очень сложно устроенными сперматозоидами. Локомоторный (двигательный) аппарат сперматозоида состоит из спирально скрученного опорного тела, или блефаропласта, с многочисленными (около сотни) жгутиками, направленными у живого сперматозоида назад и в сторону. Каждый жгутик волнообразно изгибается и описывает в пространстве коническую поверхность (рис. 88). Продолжительность плавания сперматозоидов, направление и характер их движения (длина и амплитуда волны) зависят от состава и температуры воды.Зрелый архегоний состоит из погруженного в ткань гаметофита брюшка и выступающей над его поверхностью трех-, четырехъярусной шейки. В брюшке находится яйцеклетка, над которой лежат брюшная канальцевая клетка и две шейковые канальцевые клетки (рис. 88).Оплодотворение у хвощей совершается только при наличии капельно-жидкой воды на поверхности гаметофита (во время обильной росы, дождя). Слизь, выступающая из шейки архегония, в воде разбухает и выделяет в нее вещества, привлекающие сперматозоиды, которые подплывают к шейке и проникают в ее канал, но только один из сперматозоидов сливается с яйцеклеткой.В результате оплодотворения образуется зигота, которая сразу же, прорастая, дает начало предзародышу спорофита, позднее развивающемуся в зародыш. Зародыш хвощей всасывает пищу из клеток гаметофита основанием первого листового влагалища.Возникающий из зародыша проросток обычно невелик. Его высота лишь изредка превышает 10 см. Стебель проростка несет 10—15 листовых влагалищ с тремя листовыми зубцами каждое. У нижнего узла первичного побега образуется почка, которая дает начало следующему, уже более мощному побегу со своим собственным корнем и листьями, собранными обычно в мутовку по четыре. У основания второго побега возникает затем более мощный третий побег и т. д. Первые междоузлия третьего и последующего побегов обычно изгибаются вниз и углубляются в почву, образуя нисходящее корневище, которое затем переходит в горизонтальное. В конечном счете образуется система подземных разветвленных корневищ и надземных побегов.Все до сих пор изученные в цитологическом отпошении виды хвоща имеют одинаковое хромосомное число — n=108. Столь высокое и стабильное хромосомное число в пределах рода установилось, по всей вероятности, очень давно.Хвощи характеризуются очень большой морфологической пластичностью. Например, у недавно поселившегося на хорошо освещенном участке с нарушенным растительным покровом хвоща полевого стебли обычно низкие, распростертые, но позже, когда разовьется окружающая растительность, от более зрелых корневищ того же клона могут отходить мощные прямостоячие стебли с ветвистыми веточками. При приближении к северным границам ареала этого же вида спороносные побеги, буроватые у растений умеренной зоны, сменяются зеленеющими, а потом и ветвистыми зелеными спороносными побегами тундровых растений. Очень изменчивы бывают побеги и других видов. Эти возникающие под влиянием многообразных факторов внешней среды и нередко в пределах одного клона модификации являлись и являются наряду с многочисленными гибридами и наследуемыми уродствами одной из причин разногласий в оценке объема рода.Большинство современных систематиков считает, что род хвощ (Equisetum) невелик по объему и включает около 20 крайне полиморфных видов, объединяемых в два подрода — эквизетум (Equisetum) и гиппохете (Hippochaete). Некоторые исследователи считают целесообразным выделять эти подроды в самостоятельные роды. К подроду эквизетум из упомянутых уже в тексте видов относятся хвощи боготский, болотный, большой, лесной, луговой, полевой, приречный; к подроду гиппохете — хвощи ветвистый, зимующий, камышковый, многощетинковый.Хвощи довольно широко распространены на Земле. Однако южное полушарие намного беднее ими, чем северное. Совсем нет аборигенных видов хвощей в Австралии, Новой Зеландии и в тропической Африке. В тропиках преимущественно обитают виды из подрода гиппохете, а из подрода эквизетум только два — хвощи боготский (в Цептральной и Южной Америке) и раскидистый — Е. diffusum (на полуострове Индостан). Хвощи подрода эквизетум — жители главным образом умеренной и арктической зон северного полушария.Хвощи обладают признаками как гигрофитов (слабое развитие водопроводящей системы и хорошо развитая воздухоносная ткань), так и ксерофитов (погруженные под поверхность эпидермы замыкающие клетки устьиц, закрытые устьица на старых участках стебля, сильно развитая механическая ткань, фотосинтезирующие стебли, редукция листьев). У некоторых видов хвоща (например, у приречного) на первый план выступают признаки гигрофитов; у других (например, у хвоща ветвистого) — ксерофитов; третьи (например, хвощ луговой) можно отнести к мезофитам — растениям, обитающим в местах со средней степенью увлажнения почвы; четвертые правильнее характеризовать как гигромезофиты (например, хвощ болотный) или ксеромезофиты (например, хвощ полевой). Некоторые хвощи (например, хвощ камышковый) могут жить на холодной и сырой почве и являются психрофитами. Нередко один вид хвоща обладает широкой экологической амплитудой; например, хвощ большой в одних местах ведет себя как психромезофит, в других — как гигрофит.Хвощи можно встретить в разных растительных зонах и сообществах, но в любом случае близ воды или в местах с достаточным содержанием влаги в почве или с относительно неглубоким залеганием грунтовых вод. В крайних условиях существования хвощи, по-видимому, размножаются исключительно вегетативно.Входя в состав пионерных растительных группировок и захватывая территории с нарушенным естественным растительным покровом, хвощи нередко образуют чистые или почти чистые заросли в тех местах, где другие растения не могут жить, например, из-за обилия воды или, напротив, из-за ее недостатка в тех слоях почвы, где расположена корневая система этих растений. Раз поселившись на какой-нибудь территории, хвощи благодаря наличию глубоко залегающих корневищ, масса которых превышает массу надземных частей растения в несколько раз, успешно противостоят таким неблагоприятным воздействиям внешней среды, как засухи, лесные пожары и т. п., и успешно конкурируют с другими растениями, долго удерживая захваченную территорию.Поселяясь на местах с нарушенным растительным покровом, хвощи представляют собой широко распространенные и трудно искореняемые сорняки пастбищ и полей, особенно вновь осваиваемых полей с кислыми почвами, недавно вышедшими из-под леса или из-под луга.Среди видов подрода эквизетум встречаются ядовитые для домашнего скота растения, но сведения об их ядовитости довольно противоречивы. В качестве опасных видов для лошадей указывают обычно болотный, полевой и приречный хвощи; попадаясь в большом количестве в сене, они могут явиться причиной болезни эквизетоза, которая была известна в России под названием «шатуна» или «пьяной болезни». Отравления крупного рогатого скота в Европе и Северной Америке обычно связывают с хвощом болотным, реже хвощами приречным и полевым. При поедании сена с большим количеством указанных видов (свежие растения на пастбищах коровы обычно в рот не берут) отмечаются быстрое исхудание животных, падение удоев и жирности молока. В конце концов животные, если не заменить засоренный хвощами корм, могут погибнуть от истощения. У овец при поедании хвоща полевого отмечаются исхудание и прекращение роста шерсти.Ядовитое начало в хвощах, вызывающее хроническое отравление животных, сродни тиаминазе — ферменту, разрушающему витамин B1. Случаи острого отравления животных, по-видимому, связаны с наличием у хвощей сапонинов (эквизетонин) и флавоновых гликозидов, количество которых в растении варьирует в зависимости от места обитания, времени года и многих других условий, и этим, возможно, объясняется противоречивость сведений о ядовитости того или иного вида для разных животных в разных областях.Гораздо меньше сведений о ядовитости для скота хвощей из подрода гиппохете. Более того, экспериментальным путем и достоверными наблюдениями в природе подтверждена большая кормовая ценность многих видов этого подрода для лошадей, коров, северного и пятнистого оленей, кабанов. Но следует отметить, что дикие и домашние животные (при вольном выпасе) используют хвощи подрода гиппохете в пищу главным образом после наступления сильных морозов осенью и далее зимой, а также очень рано весной. Летом они их не поедают. Это объясняется изменчивостью химического состава хвощей в течение года. Например, накопленный за лето хвощом зимующим крахмал превращается в сахара при наступлении низких температур.О ядовитости хвощей для человека сведений нет. Напротив, молодые, чуть сладковатые спороносные побеги хвоща полевого и крахмалоносные клубни этого вида ранее широко использовало в пищу бедное население Евразии и Северной Америки. Многие хвощи применяли и сейчас применяют в народной медицине, а хвощ полевой входит в отечественную государственную фармакопею.До XX в. полевой и лесной хвощи использовали для окрашивания шерсти в серо-желтый цвет. Индейцы Северной Америки ранее употребляли корневища хвоща болотного для плетения корзин. Жесткие стебли хвоща зимующего и сходных видов широко использовали в Европе и Северной Америке вместо наждачной бумаги для технических надобностей: для полировки мебели и рога, для очистки металлических частей при лужении и паянии и просто для чистки металлической посуды.
Из споры после первого деления образуются положительно геотропичный ризоид и заростковая клетка (клетка гаметофита), которая многократно делится и дает начало зеленому гаметофиту (рис. 88). При слабом освещении или в загущенных посевах гаметофит имеет вид однорядной зеленой нити — хлоронемы. При достаточном освещении нитевидная стадия в развитии гаметофита обычно не выражена, и он сразу принимает вид однослойной пластинки, нарастающей с помощью верхушечной меристемы. Некоторые клетки пластинчатого гаметофита дают начало вторичным ризоидам. Спустя некоторое время двухмерный пластинчатый гаметофит превращается в трехмерный гаметофит с более или менее массивной распростертой по субстрату базальной частью — подушкой. На верхней стороне подушки развиваются прямостоячие зеленые пластинки длиной до 1—4 мм, а на нижней — располагаются бесцветные, длиной до 1 см ризоиды, прикрепляющие гаметофит к почве и высасывающие из нее воду с минеральными солями. При хороших условиях освещения и питания по краю подушки вскоре дифференцируется меристема, в результате деятельности которой увеличивается как объем подушки, так и число пластинок и ризоидов.При проращивании одиночных спор на стерильной питательной среде удалось выяснить, что у хвощей имеется три типа гаметофитов: мужские, женские и обоеполые, различающиеся между собой по скорости роста, темпам развития и морфологии. Размеры мужских гаметофитов у хвощей варьируют в диаметре от 1 до 10 мм, женских — от 3 до 30 мм. У одного и того же вида мужские гаметофиты при равных условиях культуры мельче женских. Первые антеридии появляются на мужских гаметофитах в зависимости от вида хвоща спустя 20 — 110 дней после посева спор, а первые архегонии на женских гаметофитах — спустя 30 — 130 дней.При формировании мужского гаметофита краевая меристема образует небольшое число пластинок, а затем дает начало антеридиальной ветви — резко изогнутой кверху лопасти, на нижней стороне которой по краю возникают антеридии.У гаметофитов, находимых в природе, число антеридиев обычно невелико. В культуре старые гаметофиты могут иметь несколько ветвей с десятками и сотнями антеридиев. Антеридии могут возникать и на кончиках пластинок, а иногда пластинки прорастают и дают начало более или менее массивному телу с краевой меристемой. Эта краевая меристема или может дать начало новым мужским ветвям с собственной меристемой, или из нее у немногих видов образуется подушка с пластинками и с архегониями, и, таким образом, первоначально мужской гаметофит превращается в обоеполый.При формировании женского гаметофита краевая меристема обычно не прекращает своей деятельности по образованию подушки, пластинок и ризоидов, но, когда подушка достигнет определенной величины, из некоторых клеток меристемы возникают архегонии. Зрелые архегонии располагаются у основания пластинок. У неветвистых гаметофитов спустя некоторое время меристема по всему краю подушки начинает формировать антеридии. Если гаметофиты ветвистые, то краевая меристема только некоторых ветвей перерождается и дает начало антеридиальным ветвям с антеридиями, в основании которых затем возникает антеридиальная меристема. Таким образом, первоначально женский гаметофит превращается в обоеполый, несущий одновременно и архегонии и антеридии. Постепенно все ветви женского гаметофита перерождаются в антеридиальные, и после отмирания архегониев гаметофит становится функционально мужским.В культуре число архегониев на одном гаметофите варьирует от 20 до нескольких сотен; у гаметофитов, находимых в природе, число архегониев нередко много меньше.Следует отметить, что женские гаметофиты всех видов хвоща могут стать обоеполыми, и это резко контрастирует с неспособностью мужских гаметофитов подавляющего большинства представителей этого рода формировать архегонии.Гаметофиты хвощей очень чувствительны к воздействию окружающих условий. В опыте со строго контролируемыми и постоянными факторами внешней среды нередко можно заметить различия в морфологии гаметофитов разных видов и подродов. У гаметофитов, выросших в природе, видовые различия подметить трудно или вовсе не удается, да и сами дикорастущие гаметофиты иногда очень резко отличаются от культивируемых по величине подушки, числу и длине лопастей и пластинок, скорости развития, числу гаметангиев и т. д. В природе гаметофиты могут не успеть до наступления неблагоприятной погоды приступить к формированию гаметангиев другого пола, и популяция, таким образом, будет состоять только из мужских и женских гаметофитов.Обычно количество мужских гаметофитов в популяции очень сильно варьирует (от 3 до 100%) в зависимости и от вида хвоща, и от разнообразных условий среды. В целом условия, благоприятствующие росту молодых гаметофитов, приводят к формированию большего числа женских гаметофитов, а при неблагоприятных условиях роста образуется больше мужских. Из долго хранившихся спор, обладающих малой энергией прорастания, возникает больше мужских гаметофитов, чем при тех же условиях из свежесобранных спор.Зрелые антеридии разных видов хвоща отличаются друг от друга формой полости, в которой лежат сперматоциты, степенью своего возвышения над поверхностью гаметофита, числом, формой и расположением клеток крышечки.Хвощи обладают крупными и очень сложно устроенными сперматозоидами. Локомоторный (двигательный) аппарат сперматозоида состоит из спирально скрученного опорного тела, или блефаропласта, с многочисленными (около сотни) жгутиками, направленными у живого сперматозоида назад и в сторону. Каждый жгутик волнообразно изгибается и описывает в пространстве коническую поверхность (рис. 88). Продолжительность плавания сперматозоидов, направление и характер их движения (длина и амплитуда волны) зависят от состава и температуры воды.Зрелый архегоний состоит из погруженного в ткань гаметофита брюшка и выступающей над его поверхностью трех-, четырехъярусной шейки. В брюшке находится яйцеклетка, над которой лежат брюшная канальцевая клетка и две шейковые канальцевые клетки (рис. 88).Оплодотворение у хвощей совершается только при наличии капельно-жидкой воды на поверхности гаметофита (во время обильной росы, дождя). Слизь, выступающая из шейки архегония, в воде разбухает и выделяет в нее вещества, привлекающие сперматозоиды, которые подплывают к шейке и проникают в ее канал, но только один из сперматозоидов сливается с яйцеклеткой.В результате оплодотворения образуется зигота, которая сразу же, прорастая, дает начало предзародышу спорофита, позднее развивающемуся в зародыш. Зародыш хвощей всасывает пищу из клеток гаметофита основанием первого листового влагалища.Возникающий из зародыша проросток обычно невелик. Его высота лишь изредка превышает 10 см. Стебель проростка несет 10—15 листовых влагалищ с тремя листовыми зубцами каждое. У нижнего узла первичного побега образуется почка, которая дает начало следующему, уже более мощному побегу со своим собственным корнем и листьями, собранными обычно в мутовку по четыре. У основания второго побега возникает затем более мощный третий побег и т. д. Первые междоузлия третьего и последующего побегов обычно изгибаются вниз и углубляются в почву, образуя нисходящее корневище, которое затем переходит в горизонтальное. В конечном счете образуется система подземных разветвленных корневищ и надземных побегов.Все до сих пор изученные в цитологическом отпошении виды хвоща имеют одинаковое хромосомное число — n=108. Столь высокое и стабильное хромосомное число в пределах рода установилось, по всей вероятности, очень давно.Хвощи характеризуются очень большой морфологической пластичностью. Например, у недавно поселившегося на хорошо освещенном участке с нарушенным растительным покровом хвоща полевого стебли обычно низкие, распростертые, но позже, когда разовьется окружающая растительность, от более зрелых корневищ того же клона могут отходить мощные прямостоячие стебли с ветвистыми веточками. При приближении к северным границам ареала этого же вида спороносные побеги, буроватые у растений умеренной зоны, сменяются зеленеющими, а потом и ветвистыми зелеными спороносными побегами тундровых растений. Очень изменчивы бывают побеги и других видов. Эти возникающие под влиянием многообразных факторов внешней среды и нередко в пределах одного клона модификации являлись и являются наряду с многочисленными гибридами и наследуемыми уродствами одной из причин разногласий в оценке объема рода.Большинство современных систематиков считает, что род хвощ (Equisetum) невелик по объему и включает около 20 крайне полиморфных видов, объединяемых в два подрода — эквизетум (Equisetum) и гиппохете (Hippochaete). Некоторые исследователи считают целесообразным выделять эти подроды в самостоятельные роды. К подроду эквизетум из упомянутых уже в тексте видов относятся хвощи боготский, болотный, большой, лесной, луговой, полевой, приречный; к подроду гиппохете — хвощи ветвистый, зимующий, камышковый, многощетинковый.Хвощи довольно широко распространены на Земле. Однако южное полушарие намного беднее ими, чем северное. Совсем нет аборигенных видов хвощей в Австралии, Новой Зеландии и в тропической Африке. В тропиках преимущественно обитают виды из подрода гиппохете, а из подрода эквизетум только два — хвощи боготский (в Цептральной и Южной Америке) и раскидистый — Е. diffusum (на полуострове Индостан). Хвощи подрода эквизетум — жители главным образом умеренной и арктической зон северного полушария.Хвощи обладают признаками как гигрофитов (слабое развитие водопроводящей системы и хорошо развитая воздухоносная ткань), так и ксерофитов (погруженные под поверхность эпидермы замыкающие клетки устьиц, закрытые устьица на старых участках стебля, сильно развитая механическая ткань, фотосинтезирующие стебли, редукция листьев). У некоторых видов хвоща (например, у приречного) на первый план выступают признаки гигрофитов; у других (например, у хвоща ветвистого) — ксерофитов; третьи (например, хвощ луговой) можно отнести к мезофитам — растениям, обитающим в местах со средней степенью увлажнения почвы; четвертые правильнее характеризовать как гигромезофиты (например, хвощ болотный) или ксеромезофиты (например, хвощ полевой). Некоторые хвощи (например, хвощ камышковый) могут жить на холодной и сырой почве и являются психрофитами. Нередко один вид хвоща обладает широкой экологической амплитудой; например, хвощ большой в одних местах ведет себя как психромезофит, в других — как гигрофит.Хвощи можно встретить в разных растительных зонах и сообществах, но в любом случае близ воды или в местах с достаточным содержанием влаги в почве или с относительно неглубоким залеганием грунтовых вод. В крайних условиях существования хвощи, по-видимому, размножаются исключительно вегетативно.Входя в состав пионерных растительных группировок и захватывая территории с нарушенным естественным растительным покровом, хвощи нередко образуют чистые или почти чистые заросли в тех местах, где другие растения не могут жить, например, из-за обилия воды или, напротив, из-за ее недостатка в тех слоях почвы, где расположена корневая система этих растений. Раз поселившись на какой-нибудь территории, хвощи благодаря наличию глубоко залегающих корневищ, масса которых превышает массу надземных частей растения в несколько раз, успешно противостоят таким неблагоприятным воздействиям внешней среды, как засухи, лесные пожары и т. п., и успешно конкурируют с другими растениями, долго удерживая захваченную территорию.Поселяясь на местах с нарушенным растительным покровом, хвощи представляют собой широко распространенные и трудно искореняемые сорняки пастбищ и полей, особенно вновь осваиваемых полей с кислыми почвами, недавно вышедшими из-под леса или из-под луга.Среди видов подрода эквизетум встречаются ядовитые для домашнего скота растения, но сведения об их ядовитости довольно противоречивы. В качестве опасных видов для лошадей указывают обычно болотный, полевой и приречный хвощи; попадаясь в большом количестве в сене, они могут явиться причиной болезни эквизетоза, которая была известна в России под названием «шатуна» или «пьяной болезни». Отравления крупного рогатого скота в Европе и Северной Америке обычно связывают с хвощом болотным, реже хвощами приречным и полевым. При поедании сена с большим количеством указанных видов (свежие растения на пастбищах коровы обычно в рот не берут) отмечаются быстрое исхудание животных, падение удоев и жирности молока. В конце концов животные, если не заменить засоренный хвощами корм, могут погибнуть от истощения. У овец при поедании хвоща полевого отмечаются исхудание и прекращение роста шерсти.Ядовитое начало в хвощах, вызывающее хроническое отравление животных, сродни тиаминазе — ферменту, разрушающему витамин B1. Случаи острого отравления животных, по-видимому, связаны с наличием у хвощей сапонинов (эквизетонин) и флавоновых гликозидов, количество которых в растении варьирует в зависимости от места обитания, времени года и многих других условий, и этим, возможно, объясняется противоречивость сведений о ядовитости того или иного вида для разных животных в разных областях.Гораздо меньше сведений о ядовитости для скота хвощей из подрода гиппохете. Более того, экспериментальным путем и достоверными наблюдениями в природе подтверждена большая кормовая ценность многих видов этого подрода для лошадей, коров, северного и пятнистого оленей, кабанов. Но следует отметить, что дикие и домашние животные (при вольном выпасе) используют хвощи подрода гиппохете в пищу главным образом после наступления сильных морозов осенью и далее зимой, а также очень рано весной. Летом они их не поедают. Это объясняется изменчивостью химического состава хвощей в течение года. Например, накопленный за лето хвощом зимующим крахмал превращается в сахара при наступлении низких температур.О ядовитости хвощей для человека сведений нет. Напротив, молодые, чуть сладковатые спороносные побеги хвоща полевого и крахмалоносные клубни этого вида ранее широко использовало в пищу бедное население Евразии и Северной Америки. Многие хвощи применяли и сейчас применяют в народной медицине, а хвощ полевой входит в отечественную государственную фармакопею.До XX в. полевой и лесной хвощи использовали для окрашивания шерсти в серо-желтый цвет. Индейцы Северной Америки ранее употребляли корневища хвоща болотного для плетения корзин. Жесткие стебли хвоща зимующего и сходных видов широко использовали в Европе и Северной Америке вместо наждачной бумаги для технических надобностей: для полировки мебели и рога, для очистки металлических частей при лужении и паянии и просто для чистки металлической посуды.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.
.