- Порядок ламинариевые (Laminariales)
- Порядок ламинариевые (Laminariales)
-
Ламинариевые имеют гетероморфный цикл развития с обязательным чередованием спорофитов и гаметофитов.Зрелые спорофиты — крупные растения высотой 0,5—6 м, для отдельных видов обычны размеры в 20 м. Самые крупные ламинариевые достигают 50 м. Слоевище ламинариевых имеет одну или несколько пластин, расположенных на простом или разветвленном стволе. Слоевище прикрепляется к грунту диском или разветвленными ризоидами. Внутри пластины и ствола можно различить меристодерму и кору, состоящие из окрашенных клеток, промежуточный слой из крупных бесцветных клеток и сердцевину. У одних ламинариевых (подпорядок филлариевые) сердцевина состоит из плотно сомкнутых рядов цилиндрических нешироких клеток с оболочками обычной толщины и рядов широких клеток с толстыми боковыми стенками. Сердцевина других ламинариевых (подпорядок ламинариевые) образована неплотно расположенными клеточными нитями: одни из них идут поперек слоевища от клеток промежуточного слоя; другие ориентированы по длинной оси слоевища, причем клетки последних очень длинные и узкие и только у поперечных перегородок они воронкообразно расширяются (рис. 139,1, 2).
 Эти нити называют трубчатыми нитями, а их поперечные перегородки — ситовидными пластинами, так как в них находятся поры. В краевой части сердцевины крупных ламинариевых, таких, как макроцистис и нереоцистис, среди тонких-нитей располагаются ситовидные трубки. Они выделяются клетками большого диаметра по всей длине, их поперечными перегородками служат ситовидные пластины. В коре или промежуточном слое многих ламинариевых расположены слизистые каналы, образующие в слоевище сеть, петли которой вытянуты по его продольной оси. В некоторых участках слизистых каналов, особенно в местах их ветвления, располагаются группы секреторных клеток (рис. 140), содержащих большое количество диктиосом. Роль секреторных клеток и слизистых каналов ламинариевых пока не ясна; видимо, они выполняют какую-то физиологическую функцию. Количество их в слоевищах одного вида изменяется в зависимости от условий произрастания, и иногда они могут совершенно не развиваться.
Эти нити называют трубчатыми нитями, а их поперечные перегородки — ситовидными пластинами, так как в них находятся поры. В краевой части сердцевины крупных ламинариевых, таких, как макроцистис и нереоцистис, среди тонких-нитей располагаются ситовидные трубки. Они выделяются клетками большого диаметра по всей длине, их поперечными перегородками служат ситовидные пластины. В коре или промежуточном слое многих ламинариевых расположены слизистые каналы, образующие в слоевище сеть, петли которой вытянуты по его продольной оси. В некоторых участках слизистых каналов, особенно в местах их ветвления, располагаются группы секреторных клеток (рис. 140), содержащих большое количество диктиосом. Роль секреторных клеток и слизистых каналов ламинариевых пока не ясна; видимо, они выполняют какую-то физиологическую функцию. Количество их в слоевищах одного вида изменяется в зависимости от условий произрастания, и иногда они могут совершенно не развиваться. Для представителей семейства алариевых характерно присутствие в поверхностном слое пластин крупных клеток, называемых гландулярными; они наполнены фукозаном. В молодых частях слоевища эти клетки желтые, в старых частях они темно-бурые или черные.Рост слоевищ спорофитов осуществляется посредством интеркалярной зоны роста. Она занимает основание пластины и вершину ствола. Если ствол разветвленный и имеется несколько пластинок, то интеркалярная зона роста находится в основании каждой пластины. Интеркалярная зона роста не отграничена резко от остальной части слоевища. Деление клеток и соответственно рост наблюдается и за ее пределами.Наряду с процессами роста в пластинах спорофита ламинариевых происходят процессы разрушения верхнего конца. Нанося на пластину через определенные расстояния отверстия и измеряя, как эти расстояния увеличиваются со временем, установили, что за время роста пластина обновляется по длине 1—5 раз, а по массе до 20 раз. Слоевища ламинариевых обладают наибольшей скоростью роста, достигающей более 10 см в сутки. У нереоцистиса Лютке наблюдалось удлинение ствола на 13,5 см в сутки, не считая роста пластин. Особенно быстро слоевища ламинариевых растут в конце зимы и весной, когда температура воды нередко держится около 0°С. В период бурного роста пластина содержит много воды и поэтому, если смять ее рукой, легко трескается. С повышением температуры воды скорость роста замедляется и усиливаются процессы синтеза и накопления органических веществ. Пластины становятся толще, темнее и перестают трескаться при сминании. Наиболее интенсивно накопление органических веществ происходит в основании пластины. В то же время наибольшая скорость фотосинтеза отмечается в верхней ее части. Наличие сердцевины с хорошо развитыми проводящими элементами обеспечивает транспортировку веществ в пределах слоевища.Большинство ламинариевых — многолетние растения. Ежегодно после завершения выхода спор пластинчатая часть слоевища целиком разрушается. Этот процесс начинается с вершины. Тогда же или несколько позднее в интеркалярной зоне роста начинается интенсивное деление клеток и происходит закладка новой пластины. Она появляется между стволом и старой пластиной. У одних ламинариевых между двумя пластинами хорошо выражен пережим, у других он отсутствует. Обычно новая пластина шире старой. По мере роста новой пластины старая отодвигается все дальше вверх, иногда ее остатки могут долго сохраняться на вершине. При низкой температуре воды разрушение старой пластины происходит медленно. В Арктике у ламинарии копытной (Laminaria solidungula) на вершине новой пластины можно наблюдать цепочку из трех-четырех старых пластин. Как свидетельство многолетнего роста некоторых ламинариевых, в их стволах заметны кольца. Лучше всего они выражены в самом низу — в наиболее старой части ствола.
Для представителей семейства алариевых характерно присутствие в поверхностном слое пластин крупных клеток, называемых гландулярными; они наполнены фукозаном. В молодых частях слоевища эти клетки желтые, в старых частях они темно-бурые или черные.Рост слоевищ спорофитов осуществляется посредством интеркалярной зоны роста. Она занимает основание пластины и вершину ствола. Если ствол разветвленный и имеется несколько пластинок, то интеркалярная зона роста находится в основании каждой пластины. Интеркалярная зона роста не отграничена резко от остальной части слоевища. Деление клеток и соответственно рост наблюдается и за ее пределами.Наряду с процессами роста в пластинах спорофита ламинариевых происходят процессы разрушения верхнего конца. Нанося на пластину через определенные расстояния отверстия и измеряя, как эти расстояния увеличиваются со временем, установили, что за время роста пластина обновляется по длине 1—5 раз, а по массе до 20 раз. Слоевища ламинариевых обладают наибольшей скоростью роста, достигающей более 10 см в сутки. У нереоцистиса Лютке наблюдалось удлинение ствола на 13,5 см в сутки, не считая роста пластин. Особенно быстро слоевища ламинариевых растут в конце зимы и весной, когда температура воды нередко держится около 0°С. В период бурного роста пластина содержит много воды и поэтому, если смять ее рукой, легко трескается. С повышением температуры воды скорость роста замедляется и усиливаются процессы синтеза и накопления органических веществ. Пластины становятся толще, темнее и перестают трескаться при сминании. Наиболее интенсивно накопление органических веществ происходит в основании пластины. В то же время наибольшая скорость фотосинтеза отмечается в верхней ее части. Наличие сердцевины с хорошо развитыми проводящими элементами обеспечивает транспортировку веществ в пределах слоевища.Большинство ламинариевых — многолетние растения. Ежегодно после завершения выхода спор пластинчатая часть слоевища целиком разрушается. Этот процесс начинается с вершины. Тогда же или несколько позднее в интеркалярной зоне роста начинается интенсивное деление клеток и происходит закладка новой пластины. Она появляется между стволом и старой пластиной. У одних ламинариевых между двумя пластинами хорошо выражен пережим, у других он отсутствует. Обычно новая пластина шире старой. По мере роста новой пластины старая отодвигается все дальше вверх, иногда ее остатки могут долго сохраняться на вершине. При низкой температуре воды разрушение старой пластины происходит медленно. В Арктике у ламинарии копытной (Laminaria solidungula) на вершине новой пластины можно наблюдать цепочку из трех-четырех старых пластин. Как свидетельство многолетнего роста некоторых ламинариевых, в их стволах заметны кольца. Лучше всего они выражены в самом низу — в наиболее старой части ствола. Спорофиты всех ламинариевых образуют органы бесполого размножения одного типа — одноклеточные одногнездные спорангии, которые развиваются на поверхности слоевища среди одноклеточных булавовидных парафиз, содержащих в верхнем, расширенном конце хлоропласты и физоды (рис. 141). Лишь когда парафизы достигнут почти полного развития, начинают расти спорангии. У всех ламинариевых они имеют сходную форму — эллиптическую или цилиндрическую. Число зооспор в спорангиях колеблется в зависимости от вида и отчасти от внешних условий от 16 до 128. Образование зооспор сопровождается мейозом. При развитии спорангиев на пластинчатых частях слоевища появляются большие по площади пятна (или одно пятно), слегка (на доли миллиметра) поднимающиеся над поверхностью и незначительно выделяющиеся окраской. После созревания и выхода зооспор происходит разрушение спороносных участков пластин, на их месте образуются отверстия, а вскоре разрушается и вся пластина. Массовое развитие спорангиев у ламинариевых начинается в конце лета — начале осени. В холодных морях они сохраняются на слоевищах до начала следующего лета. На единичных экземплярах спорангии могут появляться в начале лета, а также в иные времена года.Зооспоры ламинариевых имеют типичное для подвижных клеток бурых водорослей строение. Они грушевидные, длиной 5—10 мкм, с двумя жгутиками, отходящими сбоку. Передний жгутик перистый, задний — бичевидный. Отчетливо видимая оболочка у зооспор отсутствует. Внутри, как правило, содержится по одному хлоропласту. Относительно существования глазка у зооспор ламинариевых имеются противоречивые сведения. При наиболее подходящих температурных условиях зооспоры способны к активному движению в течение двух суток. При высокой температуре (для ламинариевых морей СССР это +20°С и выше) они проявляют активность всего несколько минут или вообще не способны двигаться. Если за время активного движения зооспора не найдет место для прикрепления, она погибает. Направление движения зооспор определяется интенсивностью света. При высокой освещенности зооспоры проявляют отрицательный фототаксис. Встретив твердую поверхность, зооспора прикрепляется передним жгутиком, который сокращается и подтягивает тело зооспоры к субстрату. После этого зооспора теряет жгутики и округляется, у нее появляется ясно видимая оболочка, и зооспора превращается в эмбриоспору, которая прорастает без периода покоя.В течение 1—2 суток эмбриоспора дает вырост — проростковую трубку, в которую переходит все содержимое. Направление роста проростковой трубки определяется светом и расстоянием от других эмбриоспор. При небольшом расстоянии между эмбриоспорами их проростковые трубки направлены в разные стороны. Протоплазма, переместившаяся в проростковую трубку, отделяется от опустевшей эмбриоспоры поперечной перегородкой (рис. 142). Так образуется первая клетка гаметофита. Оболочка эмбриоспоры долго сохраняется у гаметофита, часто даже тогда, когда на нем появляется спорофит. Гаметофиты большинства ламинариевых, за редким исключением, раздельнополые и различаются морфологически. Различия между мужскими и женскими гаметофитами заметны уже на стадии первой клетки. У женского гаметофита она короче и шире. В каждом спорангии ламинариевых образуется равное количество спор, дающих мужские и женские гаметофиты. Повышенная температура воды в первую очередь вызывает гибель женских спор.
Спорофиты всех ламинариевых образуют органы бесполого размножения одного типа — одноклеточные одногнездные спорангии, которые развиваются на поверхности слоевища среди одноклеточных булавовидных парафиз, содержащих в верхнем, расширенном конце хлоропласты и физоды (рис. 141). Лишь когда парафизы достигнут почти полного развития, начинают расти спорангии. У всех ламинариевых они имеют сходную форму — эллиптическую или цилиндрическую. Число зооспор в спорангиях колеблется в зависимости от вида и отчасти от внешних условий от 16 до 128. Образование зооспор сопровождается мейозом. При развитии спорангиев на пластинчатых частях слоевища появляются большие по площади пятна (или одно пятно), слегка (на доли миллиметра) поднимающиеся над поверхностью и незначительно выделяющиеся окраской. После созревания и выхода зооспор происходит разрушение спороносных участков пластин, на их месте образуются отверстия, а вскоре разрушается и вся пластина. Массовое развитие спорангиев у ламинариевых начинается в конце лета — начале осени. В холодных морях они сохраняются на слоевищах до начала следующего лета. На единичных экземплярах спорангии могут появляться в начале лета, а также в иные времена года.Зооспоры ламинариевых имеют типичное для подвижных клеток бурых водорослей строение. Они грушевидные, длиной 5—10 мкм, с двумя жгутиками, отходящими сбоку. Передний жгутик перистый, задний — бичевидный. Отчетливо видимая оболочка у зооспор отсутствует. Внутри, как правило, содержится по одному хлоропласту. Относительно существования глазка у зооспор ламинариевых имеются противоречивые сведения. При наиболее подходящих температурных условиях зооспоры способны к активному движению в течение двух суток. При высокой температуре (для ламинариевых морей СССР это +20°С и выше) они проявляют активность всего несколько минут или вообще не способны двигаться. Если за время активного движения зооспора не найдет место для прикрепления, она погибает. Направление движения зооспор определяется интенсивностью света. При высокой освещенности зооспоры проявляют отрицательный фототаксис. Встретив твердую поверхность, зооспора прикрепляется передним жгутиком, который сокращается и подтягивает тело зооспоры к субстрату. После этого зооспора теряет жгутики и округляется, у нее появляется ясно видимая оболочка, и зооспора превращается в эмбриоспору, которая прорастает без периода покоя.В течение 1—2 суток эмбриоспора дает вырост — проростковую трубку, в которую переходит все содержимое. Направление роста проростковой трубки определяется светом и расстоянием от других эмбриоспор. При небольшом расстоянии между эмбриоспорами их проростковые трубки направлены в разные стороны. Протоплазма, переместившаяся в проростковую трубку, отделяется от опустевшей эмбриоспоры поперечной перегородкой (рис. 142). Так образуется первая клетка гаметофита. Оболочка эмбриоспоры долго сохраняется у гаметофита, часто даже тогда, когда на нем появляется спорофит. Гаметофиты большинства ламинариевых, за редким исключением, раздельнополые и различаются морфологически. Различия между мужскими и женскими гаметофитами заметны уже на стадии первой клетки. У женского гаметофита она короче и шире. В каждом спорангии ламинариевых образуется равное количество спор, дающих мужские и женские гаметофиты. Повышенная температура воды в первую очередь вызывает гибель женских спор.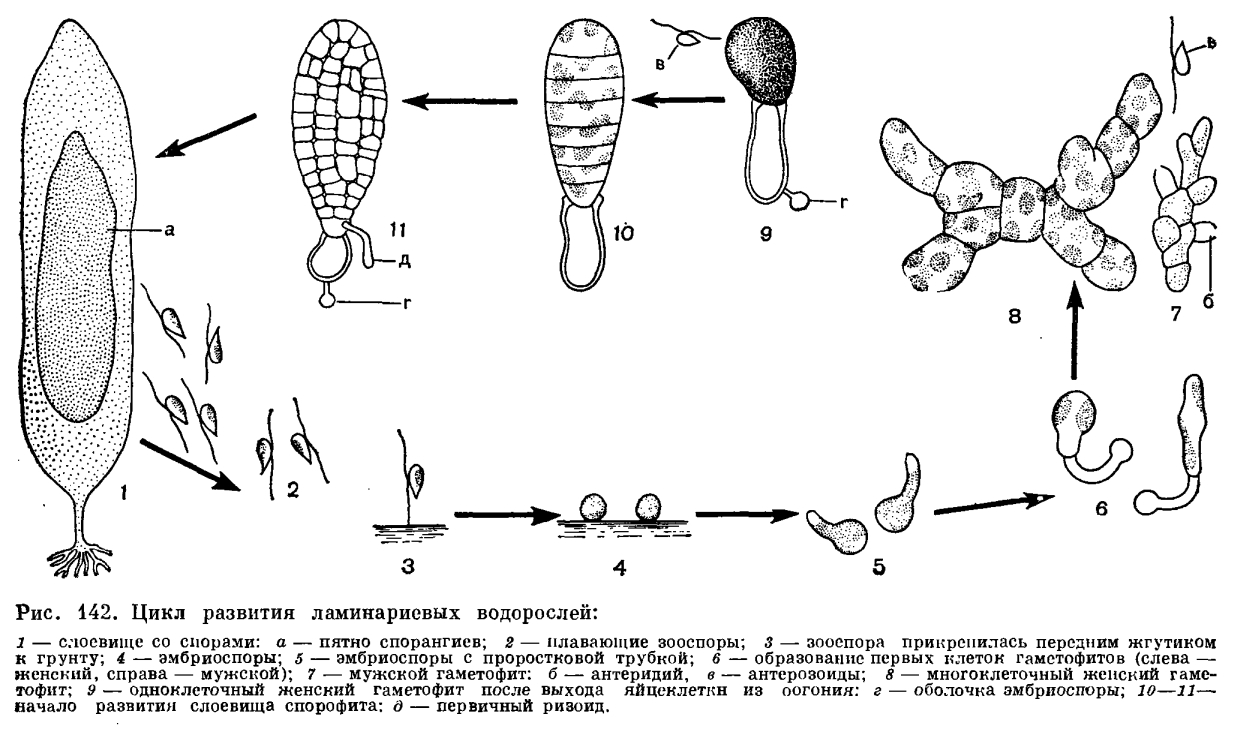 Гаметофиты ламинариевых многоклеточные, в виде однорядных разветвленных стелющихся нитей; женские гаметофиты могут быть одноклеточными. Мужские гаметофиты сильнее разветвлены и мельче женских, с более мелкими бледно окрашенными клетками. Обычно гаметофиты существуют не более одного — четырех месяцев до созревания гаметангиев. Женские многоклеточные гаметофиты не превышают в диаметре 300 мкм. При температуре, слишком высокой для образования гаметангиев, гаметофиты могут жить больше года и достигать в диаметре 1 см и более. В экспериментальных условиях гаметофиты ламинариевых способны к вегетативному размножению. Если разрезать многоклеточный гаметофит на части, из них вырастут новые гаметофиты. Это используется при всякого рода исследованиях, когда необходим генетически однородный материал. В природных условиях такое размножение едва ли возможно.Половой процесс у ламинариевых — оогамия. Антеридии имеют вид мелких бесцветных клеток, располагающихся группами или по одной на поверхности вегетативных клеток. В антеридиях образуется по одному антерозоиду. После выхода антерозоидов мужские гаметофиты погибают. У многоклеточных женских гаметофитов в оогонии превращаются большей частью конечные клетки ветвей, иногда и предыдущая клетка. Таким образом, один гаметофит несет несколько оогониев. В случае одноклеточных женских гаметофитов единственная клетка со временем превращается в единственный оогоний. Образование оогониев сопровождается увеличением размеров клеток, содержимое их окрашивается в темно-бурый цвет. В каждом оогонии формируется по одной яйцеклетке. При созревании она выходит через отверстие на вершине оогония, но не отделяется, а остается прикрепленной к краям отверстия. В таком положении происходит ее оплодотворение и рост спорофита. После выхода всех яйцеклеток гаметофит отмирает. Однако оболочки его клеток еще долго сохраняются в основании спорофитов. На месте одного многоклеточного женского гаметофита развивается несколько спорофитов, которые располагаются по его периферии. Женские гаметофиты не предоставляют никаких питательных веществ развивающимся спорофитам, они обеспечивают им только место для прикрепления на грунте. Это имеет важное значение для существования водорослей, поскольку твердые грунты занимают в море небольшую площадь, и в довершении всего их поверхность в значительной мере занята другими водорослями, а также прикрепленными животными. Основная масса зачатков морских донных водорослей (спор и гамет) погибает из-за того, что они не попадают на подходящую поверхность.Оплодотворенная яйцеклетка, оставаясь прикрепленной к гаметофиту, вырабатывает оболочку и без периода покоя трогается в рост. Вначале образуется однорядная нить примерно из семи клеток, затем наряду с поперечными перегородками появляются продольные и образуется однослойная пластинка. Из нижних клеток пластинки вырастают тонкие ризоиды. По мере роста пластинки в ней увеличивается число слоев клеток, происходит их специализация, появляются ствол и многорядные ризоиды. Если яйцеклетка почему-либо оказывается не прикрепленной к оболочке оогония, то оплодотворение происходит, но развитие спорофита идет ненормально. Образуется шарообразное скопление клеток, которое существует около месяца и погибает.В лабораторных условиях удавалось выращивать гаметофиты апоспорически, минуя зооспоры. При содержании стерильных спорофитов в аквариуме в течение трех-четырех месяцев у них отмирали клетки. Несколько клеток оставалось живыми, и они развивались в однорядные разветвленные нити, на которых образовывались оогонии и антеридии. Поскольку клетки спорофитов диплоидные, то и гаметофиты получались диплоидными. Яйцеклетки таких гаметофитов после оплодотворения развивались в тетраплоидные спорофиты. Таким же путем удавалось выращивать гаметофиты из клеток гаплоидных спорофитов, полученных посредством партеногенеза (из неоплодотворенных яйцеклеток) и апогамии (из вегетативных клеток гаметофитов). Партеногенетическому развитию спорофитов предшествует массовая дегенерация яйцеклеток после выхода их из оогониев в условиях отсутствия мужских гаметофитов. В нормальные спорофиты при этом партеногенетически развивались немногие яйцеклетки. Апогамные спорофиты вырастают из конечных клеток зрелых ветвей мужских гаметофитов.Ламинариевые обычны для .холодных вод северного и южного полушарий, где даже в самые теплые месяцы температура воды держится выше +20°С всего лишь в течение двух-трех недель. Причем в северном полушарии растет большее число родов и видов. В последнее время ламинариевые (виды рода ламинария) обнаружены в тропических водах около Бразилии и в Филиппинском море, но растут они там на глубине 70—85 м, где температура воды ниже, чем у поверхности.Ламинариевые встречаются по вертикали от поверхности моря до глубины 200 м (Laminaria rodriguezii в Средиземном море). В литоральной зоне они растут в основном в нижнем горизонте литорали в тех местах, где во время отлива подвергаются действию брызг, или там, где постоянно преобладают пасмурная погода и высокая влажность воздуха. Так, ламинариевые занимают большие площади на литорали Командорских островов, отличающихся как раз такими погодными условиями.Таким крупным водорослям, как ламинариевые, нужны условия, при которых обеспечивалось бы интенсивное поступление питательных веществ к слоевищам. Этому соответствуют места с постоянным сильным течением или прибоем. Там, где нет достаточных по силе течений или прибоя, ламинариевые растут в водах, обогащенных питательными веществами (но не до степени загрязнения) за счет разного рода стоков, содержащих много органики, или за счет неглубокого залегания илистых грунтов. Поэтому заросли ламинариевых встречаются у открытых берегов, особенно у мысов, далеко вдающихся в море. Заросли же видов, способных расти в спокойных местах, располагаются по соседству с илистыми грунтами. Это приводит к тому, что ламинариевые можно найти на глубине, в условиях пониженной освещенности, но при наличии ила среди камней, а в том же месте у берега, на меньшей глубине, их нет,— если там отсутствует постоянный прибой, а дно имеет большой уклон, из-за чего органические остатки опускаются глубже. Точно так же заросли ламинариевых легче встретить в местах с мутной водой, подверженной интенсивному движению, чем в местах с прозрачной водой, находящейся в менее активном движении.
Гаметофиты ламинариевых многоклеточные, в виде однорядных разветвленных стелющихся нитей; женские гаметофиты могут быть одноклеточными. Мужские гаметофиты сильнее разветвлены и мельче женских, с более мелкими бледно окрашенными клетками. Обычно гаметофиты существуют не более одного — четырех месяцев до созревания гаметангиев. Женские многоклеточные гаметофиты не превышают в диаметре 300 мкм. При температуре, слишком высокой для образования гаметангиев, гаметофиты могут жить больше года и достигать в диаметре 1 см и более. В экспериментальных условиях гаметофиты ламинариевых способны к вегетативному размножению. Если разрезать многоклеточный гаметофит на части, из них вырастут новые гаметофиты. Это используется при всякого рода исследованиях, когда необходим генетически однородный материал. В природных условиях такое размножение едва ли возможно.Половой процесс у ламинариевых — оогамия. Антеридии имеют вид мелких бесцветных клеток, располагающихся группами или по одной на поверхности вегетативных клеток. В антеридиях образуется по одному антерозоиду. После выхода антерозоидов мужские гаметофиты погибают. У многоклеточных женских гаметофитов в оогонии превращаются большей частью конечные клетки ветвей, иногда и предыдущая клетка. Таким образом, один гаметофит несет несколько оогониев. В случае одноклеточных женских гаметофитов единственная клетка со временем превращается в единственный оогоний. Образование оогониев сопровождается увеличением размеров клеток, содержимое их окрашивается в темно-бурый цвет. В каждом оогонии формируется по одной яйцеклетке. При созревании она выходит через отверстие на вершине оогония, но не отделяется, а остается прикрепленной к краям отверстия. В таком положении происходит ее оплодотворение и рост спорофита. После выхода всех яйцеклеток гаметофит отмирает. Однако оболочки его клеток еще долго сохраняются в основании спорофитов. На месте одного многоклеточного женского гаметофита развивается несколько спорофитов, которые располагаются по его периферии. Женские гаметофиты не предоставляют никаких питательных веществ развивающимся спорофитам, они обеспечивают им только место для прикрепления на грунте. Это имеет важное значение для существования водорослей, поскольку твердые грунты занимают в море небольшую площадь, и в довершении всего их поверхность в значительной мере занята другими водорослями, а также прикрепленными животными. Основная масса зачатков морских донных водорослей (спор и гамет) погибает из-за того, что они не попадают на подходящую поверхность.Оплодотворенная яйцеклетка, оставаясь прикрепленной к гаметофиту, вырабатывает оболочку и без периода покоя трогается в рост. Вначале образуется однорядная нить примерно из семи клеток, затем наряду с поперечными перегородками появляются продольные и образуется однослойная пластинка. Из нижних клеток пластинки вырастают тонкие ризоиды. По мере роста пластинки в ней увеличивается число слоев клеток, происходит их специализация, появляются ствол и многорядные ризоиды. Если яйцеклетка почему-либо оказывается не прикрепленной к оболочке оогония, то оплодотворение происходит, но развитие спорофита идет ненормально. Образуется шарообразное скопление клеток, которое существует около месяца и погибает.В лабораторных условиях удавалось выращивать гаметофиты апоспорически, минуя зооспоры. При содержании стерильных спорофитов в аквариуме в течение трех-четырех месяцев у них отмирали клетки. Несколько клеток оставалось живыми, и они развивались в однорядные разветвленные нити, на которых образовывались оогонии и антеридии. Поскольку клетки спорофитов диплоидные, то и гаметофиты получались диплоидными. Яйцеклетки таких гаметофитов после оплодотворения развивались в тетраплоидные спорофиты. Таким же путем удавалось выращивать гаметофиты из клеток гаплоидных спорофитов, полученных посредством партеногенеза (из неоплодотворенных яйцеклеток) и апогамии (из вегетативных клеток гаметофитов). Партеногенетическому развитию спорофитов предшествует массовая дегенерация яйцеклеток после выхода их из оогониев в условиях отсутствия мужских гаметофитов. В нормальные спорофиты при этом партеногенетически развивались немногие яйцеклетки. Апогамные спорофиты вырастают из конечных клеток зрелых ветвей мужских гаметофитов.Ламинариевые обычны для .холодных вод северного и южного полушарий, где даже в самые теплые месяцы температура воды держится выше +20°С всего лишь в течение двух-трех недель. Причем в северном полушарии растет большее число родов и видов. В последнее время ламинариевые (виды рода ламинария) обнаружены в тропических водах около Бразилии и в Филиппинском море, но растут они там на глубине 70—85 м, где температура воды ниже, чем у поверхности.Ламинариевые встречаются по вертикали от поверхности моря до глубины 200 м (Laminaria rodriguezii в Средиземном море). В литоральной зоне они растут в основном в нижнем горизонте литорали в тех местах, где во время отлива подвергаются действию брызг, или там, где постоянно преобладают пасмурная погода и высокая влажность воздуха. Так, ламинариевые занимают большие площади на литорали Командорских островов, отличающихся как раз такими погодными условиями.Таким крупным водорослям, как ламинариевые, нужны условия, при которых обеспечивалось бы интенсивное поступление питательных веществ к слоевищам. Этому соответствуют места с постоянным сильным течением или прибоем. Там, где нет достаточных по силе течений или прибоя, ламинариевые растут в водах, обогащенных питательными веществами (но не до степени загрязнения) за счет разного рода стоков, содержащих много органики, или за счет неглубокого залегания илистых грунтов. Поэтому заросли ламинариевых встречаются у открытых берегов, особенно у мысов, далеко вдающихся в море. Заросли же видов, способных расти в спокойных местах, располагаются по соседству с илистыми грунтами. Это приводит к тому, что ламинариевые можно найти на глубине, в условиях пониженной освещенности, но при наличии ила среди камней, а в том же месте у берега, на меньшей глубине, их нет,— если там отсутствует постоянный прибой, а дно имеет большой уклон, из-за чего органические остатки опускаются глубже. Точно так же заросли ламинариевых легче встретить в местах с мутной водой, подверженной интенсивному движению, чем в местах с прозрачной водой, находящейся в менее активном движении.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.
.