- Подсемейство лилейные (Lilioideae)
- Подсемейство лилейные (Lilioideae)
-
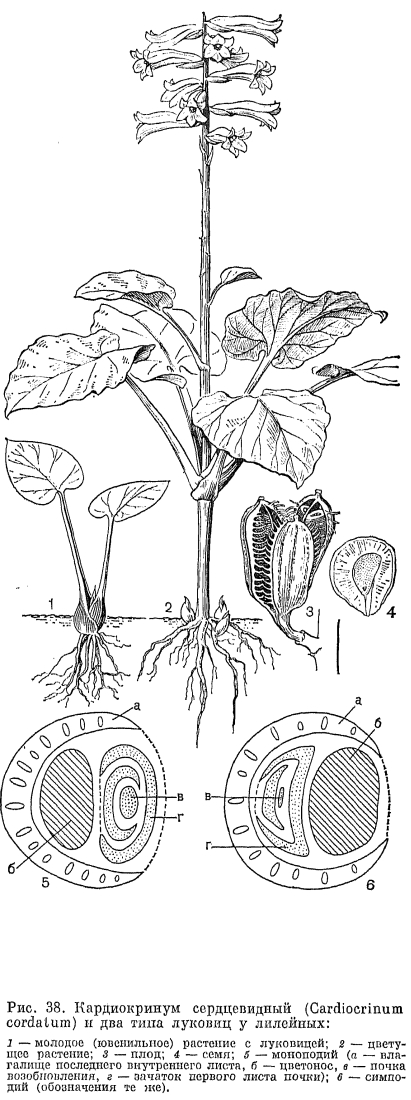
Подсемейство собственно лилейные (Lilioideae) имеет 10 родов (примерно 470 видов), относящихся к 4 трибам. Представители подсемейства встречаются только в северном полушарии. Луковицы у них составлены низовыми чешуями, так как базальные листья отсутствуют, за исключением рода кардиокринум и нескольких видов лилий. Цветоносный стебель облиственный. Сегменты околоцветника свободные, а нектарники тепальные. Пыльники прикреплены спинкой, качающиеся (триба лилейных) или прикреплены основанием, вращающиеся вокруг оси (трибы тюльпановых и гейджиевых). Нити тычинок свободные. Семена обычно плоские. Из анатомических признаков характерно отсутствие угловых клеток эпидермы. Рафиды оксалата кальция отсутствуют. Характерно наличие алкалоидов, и по химическому составу эта группа гомогенная (К. Вильямс, 1975). Однородность группы подтверждается и серологическими данными (В. С. Чупов и Н. Г. Кутявина, 1980). Триба лилейных (Lilieae) объединяет 5 родов, центром развития которых является Восточная Азия. Здесь целиком сосредоточены древние роды кардиокринум и номохарис (Nomocharis), а также наиболее древние представители родов лилия (Lilium), нотолирион (Notholirion) и рябчик. В Восточной Азии сохранились промежуточные звенья между родами —- виды с признаками обоих родов. У представителей трибы лилейных луковицы составлены незамкнутыми низовыми (кроме родов кардиокринум и нотолирион) чешуями, специализированных покровных чешуй нет. Коробочка прямостоячая, семена плоские, дисковидные, с крыловидной каймой, распространяемые по способу баллистов.Род кардиокринум (Cardiocrinum) насчитывает 3-4 вида, произрастающих в Гималаях, Китае и в Японии. Ранее его рассматривали как секцию или подрод рода лилия, однако совершенно отличный жизненный цикл, иное строение луковицы, черешковые сердцевидные листья с сетчатым жилкованием и несколько зигоморфные цветки подтверждают его родовую самостоятельность. Эти травы высотой 1,5-4 м являются монокарпиками, т. е. цветут и плодоносят единственный раз, после чего все растение отмирает. Луковица кардиокринума образована разросшимися основаниями черешков листьев; она не имеет покровных чешуй, находится почти на поверхности почвы и на свету приобретает темно-зеленый цвет (рис. 38, 1). Когда начинает расти цветоносный стебель, чешуи луковицы, расходуя на него питательные вещества, постепенно высыхают, так что к моменту цветения луковицы уже не существует. У основания стебля остаются лишь дочерние луковички, образовавшиеся в пазухах чешуи материнской луковицы. Обычно листья у кардиокринумов только стеблевые, которые собраны в ложную мутовку; у кардиокринума гигантского имеется приземная розетка и стеблевые листья очередные. Листовые пластинки достигают в диаметре 30-45 см, длина черешков около 16 см. Цветки в числе от 4 до 24 на коротких цветоножках, горизонтальные, трубчатые, длиной 12-15 см, с опадающими прицветниками, белые, снаружи зеленоватые, ароматные. Коробочка по краям растрескавшихся гнезд имеет характерные для представителей трибы лилейных выросты эпидермы в виде зубчиков (рис. 38, 3). Семена снабжены широким (до 5 мм) крыловидным краем (рис. 38, 4). В роде, по мнению одних авторов, 3 вида, по мнению других —- 4, так как кардиокринум Глена (C. glehnii), встречающийся на Сахалине и Курильских островах, часто присоединяют к японскому виду кардиокринум сердцевидный (C. cordatum). Аборигены острова Хоккайдо получают из луковиц этого растения пищевой крахмал. Все виды рода —- высокодекоративные растения.
 В роде лилия (Lilium) от 80 до 100 видов, широко распространенных в умеренной зоне северного полушария. Виды рода распространены на территории между 68° с. ш. (низовья реки Енисея) и 11° с. ш. (Южная Индия). Наиболее протяженный ареал (от Португалии до верховьев реки Лены в Сибири) у самого северного вида лилий —- саранки, или лилии кудреватой (L. martagon), которая имеет несколько разновидностей с цветками различной окраски. Большинство видов встречается в предгорьях и горных районах, в лесах, в субальпийском высокотравье вдоль ручьев, на заболоченных местах или открытых травянистых склонах. Особенно богат видами лилий (около 40) Западный Китай, Юго-Восточный Тибет и Северная Бирма.
В роде лилия (Lilium) от 80 до 100 видов, широко распространенных в умеренной зоне северного полушария. Виды рода распространены на территории между 68° с. ш. (низовья реки Енисея) и 11° с. ш. (Южная Индия). Наиболее протяженный ареал (от Португалии до верховьев реки Лены в Сибири) у самого северного вида лилий —- саранки, или лилии кудреватой (L. martagon), которая имеет несколько разновидностей с цветками различной окраски. Большинство видов встречается в предгорьях и горных районах, в лесах, в субальпийском высокотравье вдоль ручьев, на заболоченных местах или открытых травянистых склонах. Особенно богат видами лилий (около 40) Западный Китай, Юго-Восточный Тибет и Северная Бирма. Луковицы лилий многолетние, черепитчатые (чешуи незамкнутые и узкие), составлены низовыми чешуями. Лишь у нескольких видов в луковице имеются еще и листовые чешуи. Чешуи цельные или членистые, обычно в числе 8-40, но иногда их может быть 100-120, как, например, в луковице лилии Кессельринга (L. kesselringianum, табл. 6, 3), достигающей в диаметре 25 см и состоящей из 7 годичных циклов. По местонахождению почки возобновления и направлению роста побега у лилий различают ортотропные и плагиотропные луковицы (М. В. Баранова, 1981). У ортотропных луковиц почка возобновления расположена у основания цветоносного стебля, рост нового побега идет в вертикальном направлении и замещающая луковица формируется внутри материнской. У плагиотропных луковиц почка возобновления находится в пазухе нижележащих чешуй, рост нового побега идет в горизонтальном направлении и замещающая луковица располагается сбоку от материнской. Если плагиотропная луковица имеет несколько годичных побегов, она приобретает вид цепочки из нескольких луковиц, как, например, у лилии малой (L. parvum, рис. 39, 14) и некоторых других эндемичных видов западного побережья Северной Америки. Если же, кроме замещающей луковицы, имеются еще и дочерние, то образуется гнездо, которое с годами может увеличиваться до нескольких сотен луковиц, как, например, у лилии леопардовой (L. pardalinum). У некоторых видов замещающая луковица выносится за пределы материнской на толстом столоне, как у лилии канадской (L. canadense, рис. 39, 15) и других видов восточного побережья Северной Америки. От донца луковицы у лилий отходят многочисленные и сильные многолетние разветвленные корни. Некоторые из корней или их верхние участки являются втягивающими. Многие виды имеют корни на подземной части цветоносного стебля, над луковицей. Они дополнительно питают мощный цветущий стебель.
Луковицы лилий многолетние, черепитчатые (чешуи незамкнутые и узкие), составлены низовыми чешуями. Лишь у нескольких видов в луковице имеются еще и листовые чешуи. Чешуи цельные или членистые, обычно в числе 8-40, но иногда их может быть 100-120, как, например, в луковице лилии Кессельринга (L. kesselringianum, табл. 6, 3), достигающей в диаметре 25 см и состоящей из 7 годичных циклов. По местонахождению почки возобновления и направлению роста побега у лилий различают ортотропные и плагиотропные луковицы (М. В. Баранова, 1981). У ортотропных луковиц почка возобновления расположена у основания цветоносного стебля, рост нового побега идет в вертикальном направлении и замещающая луковица формируется внутри материнской. У плагиотропных луковиц почка возобновления находится в пазухе нижележащих чешуй, рост нового побега идет в горизонтальном направлении и замещающая луковица располагается сбоку от материнской. Если плагиотропная луковица имеет несколько годичных побегов, она приобретает вид цепочки из нескольких луковиц, как, например, у лилии малой (L. parvum, рис. 39, 14) и некоторых других эндемичных видов западного побережья Северной Америки. Если же, кроме замещающей луковицы, имеются еще и дочерние, то образуется гнездо, которое с годами может увеличиваться до нескольких сотен луковиц, как, например, у лилии леопардовой (L. pardalinum). У некоторых видов замещающая луковица выносится за пределы материнской на толстом столоне, как у лилии канадской (L. canadense, рис. 39, 15) и других видов восточного побережья Северной Америки. От донца луковицы у лилий отходят многочисленные и сильные многолетние разветвленные корни. Некоторые из корней или их верхние участки являются втягивающими. Многие виды имеют корни на подземной части цветоносного стебля, над луковицей. Они дополнительно питают мощный цветущий стебель.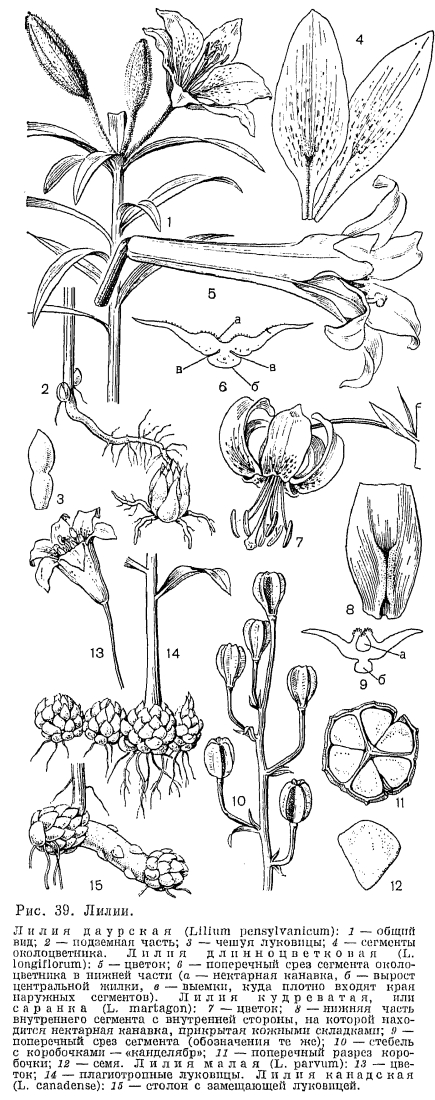 Стебель у лилий обычно прямостоячий от основания, но у некоторых восточноазиатских видов он сначала как бы блуждает под землей и может появиться на поверхности на расстоянии 60 см от луковицы. Листья обычно стеблевые, в ложных мутовках или очередные, обычно без черешков, лишь у нескольких видов имеются базальные листья (лилия белоснежная —- L. candidum и др.). Цветки горизонтальные и трубчатые (как у лилии длинноцветковой- L. longiflorum, рис. 39, 5), прямостоячие и кубковидные (как у лилии даурской, названной при первоописании по недоразумению лилией пенсильванской —- L. pensylvanicum, рис. 39, 1), поникшие и чалмовидные (как у саранки, рис. 39, 7). На форме околоцветника и основана традиционная система рода. Сегменты околоцветника варьируют у различных видов по форме, размеру и окраске. У большинства лилий и наружные и внутренние сегменты имеют нектарники в виде желобка или канавки, которая может быть открытой и голой (лилия длинноцветковая), закрытой с двух сторон кожными складками (саранка, рис. 39, 8) или окаймленной сосочками (лилия Генри —- L. henryi, табл. 4, 3). У лилии луковиценосной (L. bulbiferum), встречающейся в Центральной Европе, на концах листьев и у бутонов на кончиках внешних сегментов расположены примитивные экстрафлоральные нектарники, где весной выделяется сахаросодержащая жидкость, привлекающая муравьев (Е. Рицек, 1974). С наружной стороны сегментов у лилий имеются выросты центральной жилки, разной формы у различных видов (рис. 39, 6). В выемку между выростом жилки и спинкой внутренних сегментов плотно входят края наружных сегментов, в результате чего трубка околоцветника не распадается. Пыльники прикреплены к нитям спинками, качающиеся. Столбик значительно выше цилиндрической завязи. Рыльце головчатое, 3-лопастное.
Стебель у лилий обычно прямостоячий от основания, но у некоторых восточноазиатских видов он сначала как бы блуждает под землей и может появиться на поверхности на расстоянии 60 см от луковицы. Листья обычно стеблевые, в ложных мутовках или очередные, обычно без черешков, лишь у нескольких видов имеются базальные листья (лилия белоснежная —- L. candidum и др.). Цветки горизонтальные и трубчатые (как у лилии длинноцветковой- L. longiflorum, рис. 39, 5), прямостоячие и кубковидные (как у лилии даурской, названной при первоописании по недоразумению лилией пенсильванской —- L. pensylvanicum, рис. 39, 1), поникшие и чалмовидные (как у саранки, рис. 39, 7). На форме околоцветника и основана традиционная система рода. Сегменты околоцветника варьируют у различных видов по форме, размеру и окраске. У большинства лилий и наружные и внутренние сегменты имеют нектарники в виде желобка или канавки, которая может быть открытой и голой (лилия длинноцветковая), закрытой с двух сторон кожными складками (саранка, рис. 39, 8) или окаймленной сосочками (лилия Генри —- L. henryi, табл. 4, 3). У лилии луковиценосной (L. bulbiferum), встречающейся в Центральной Европе, на концах листьев и у бутонов на кончиках внешних сегментов расположены примитивные экстрафлоральные нектарники, где весной выделяется сахаросодержащая жидкость, привлекающая муравьев (Е. Рицек, 1974). С наружной стороны сегментов у лилий имеются выросты центральной жилки, разной формы у различных видов (рис. 39, 6). В выемку между выростом жилки и спинкой внутренних сегментов плотно входят края наружных сегментов, в результате чего трубка околоцветника не распадается. Пыльники прикреплены к нитям спинками, качающиеся. Столбик значительно выше цилиндрической завязи. Рыльце головчатое, 3-лопастное. Цветки лилий опыляются бабочками или мотыльками. Саранку, например, опыляют преимущественно ночные бабочки-бражники, а иногда также дневные бабочки. Приманкой для первых служит усиливающийся к ночи аромат цветков, для вторых —- грязно-пурпуровый с темными пятнышками околоцветник. Нектарный желобок длиной 10-15 мм, с многочисленными мелкими выростами-сосочками имеет на наружном конце вход шириной 1 мм. Порхающий бражник, просовывая свой тонкий длинный хоботок в нектарник, касается брюшком, вымазанным пыльцой другого растения, сначала выдвинутого рыльца, а затем покрытых пыльцой пыльников. Так как во время цветения столбик у лилий изгибается вверх настолько, что рыльце соприкасается с двумя верхними тычинками, возможно и самоопыление цветков. У лилии белоснежной самоопыление исключено, так как столбик не изгибается и рыльце находится на 20-25 мм выше пыльников. Лилия малая, встречающаяся на альпийских лугах в горах Сьерра-Невады в Калифорнии, имеет характерные орнитофильные трубчатые, от оранжевых до темно-красных тонов цветки, опыляемые колибри. Однако колибри кормятся часто и на энтомофильных цветках, например лилии Гумбольдта (L. humboldtii). Стебель с коробочками у лилий имеет вид канделябра. Коробочки бескрылые или крылатые, с многочисленными дисковидными семенами в каждом гнезде.Лилии хорошо размножаются и вегетативно: дочерними луковичками, которые образуются в пазухах чешуи материнской луковицы; чешуями или их члениками, а также луковичками-бульбочками, которые образуются на надземной части стебля в пазухах листьев (у лилии луковиценосной) или на подземной части стебля в зоне стеблевых корней.Лилии —- первоклассные декоративные растения, и начало их культуры восходит к далекому прошлому. Древнейшая из культивируемых лилий —- лилия белоснежная, известная также как лилия Мадонны, лилия Благовещения, в диком состоянии встречается кое-где в Ливане, Палестине и Сирии на известняковых скалах, среди средиземноморской растительности. Ее разводили и до сих пор разводят из-за красивых и ароматных цветков и для получения эфирного масла и лекарственных препаратов. Первые изображения этой лилии встречаются на Критских вазах и фресках начиная с 1750 г. до н. э., а затем у древних ассирийцев, египтян, греков и римлян. Древние римляне считали, что после розы нет цветка более прекрасного, чем лилия. От древних римлян лилия попала в завоеванные ими страны Европы. В раннем средневековье ее выращивали монахи в монастырях сначала как лекарственное растение. Белизна ее цветков считалась символом правды, чистоты и непорочности, поэтому у ранних христиан цветки лилии белоснежной стали атрибутами девы Марии-богоматери. Начиная с эпохи итальянского Возрождения и ранней фламандской живописи в картинах на библейский сюжет «Благовещение» изображался архангел Гавриил, приносящий мадонне белую лилию. Карл Линней, давший ботаническое название этой лилии, следовал латинским поэтам, которые начиная с Вергилия (70-19 гг. до н. э.) называли ее белоснежной. С XVI в. в культуру ввели и другие виды лилий. С конца XVIII в. начались работы по гибридизации. В начале XX в. с нахождением в Западном Китае лилии царственной (L. regale) значительно увеличивается число сортов лилий, достигшее теперь 2000. В Восточной Азии лилии разводят как овощную культуру, ради съедобных луковиц. Использовали в пищу и луковицы сибирских видов. В луковице лилии даурской обнаружены сердечные гликозиды. В народной медицине использовали и луковицы саранки, которая, согласно старой сибирской легенде; возникла из сердца казачьего атамана Ермака, погибшего в 1585 г. в бою с ханом Кучумом на реке Иртыш.В роде нотолирион (Notholirion) 5 видов, которые встречаются в Восточной и Западной Азии на высокогорных лугах. Три из них приурочены к Восточной Азии. Нотолирион Томсона (N. thomsonianum) произрастает в сравнительно сухих областях, на солнечных склонах от Центральных Гималаев до Северо-Западного Афганистана. Самый западный вид —- нотолирион Коэ (N. koeiei) —- найден на горных болотах хребта Загрос в Западном Иране. Виды нотолириона- монокарпические растения, которые отличаются от лилий строением луковицы и отчетливо 3-раздельным рыльцем. Луковица нотолириона состоит из влагалищ базальных листьев и низовых чешуй, в пазухах которых образуются дочерние луковички, остающиеся в земле после отмирания растения. Покровными чешуями служат сухие основания листьев. Листья базальные и стеблевые. Цветки трубчатые или колокольчатые, иногда немного зигоморфные, протандричные (табл. 5, 1). В основании сегментов околоцветника имеется нектарная ямка, как у видов рябчика. Семена мелкие, бескрылые.
Цветки лилий опыляются бабочками или мотыльками. Саранку, например, опыляют преимущественно ночные бабочки-бражники, а иногда также дневные бабочки. Приманкой для первых служит усиливающийся к ночи аромат цветков, для вторых —- грязно-пурпуровый с темными пятнышками околоцветник. Нектарный желобок длиной 10-15 мм, с многочисленными мелкими выростами-сосочками имеет на наружном конце вход шириной 1 мм. Порхающий бражник, просовывая свой тонкий длинный хоботок в нектарник, касается брюшком, вымазанным пыльцой другого растения, сначала выдвинутого рыльца, а затем покрытых пыльцой пыльников. Так как во время цветения столбик у лилий изгибается вверх настолько, что рыльце соприкасается с двумя верхними тычинками, возможно и самоопыление цветков. У лилии белоснежной самоопыление исключено, так как столбик не изгибается и рыльце находится на 20-25 мм выше пыльников. Лилия малая, встречающаяся на альпийских лугах в горах Сьерра-Невады в Калифорнии, имеет характерные орнитофильные трубчатые, от оранжевых до темно-красных тонов цветки, опыляемые колибри. Однако колибри кормятся часто и на энтомофильных цветках, например лилии Гумбольдта (L. humboldtii). Стебель с коробочками у лилий имеет вид канделябра. Коробочки бескрылые или крылатые, с многочисленными дисковидными семенами в каждом гнезде.Лилии хорошо размножаются и вегетативно: дочерними луковичками, которые образуются в пазухах чешуи материнской луковицы; чешуями или их члениками, а также луковичками-бульбочками, которые образуются на надземной части стебля в пазухах листьев (у лилии луковиценосной) или на подземной части стебля в зоне стеблевых корней.Лилии —- первоклассные декоративные растения, и начало их культуры восходит к далекому прошлому. Древнейшая из культивируемых лилий —- лилия белоснежная, известная также как лилия Мадонны, лилия Благовещения, в диком состоянии встречается кое-где в Ливане, Палестине и Сирии на известняковых скалах, среди средиземноморской растительности. Ее разводили и до сих пор разводят из-за красивых и ароматных цветков и для получения эфирного масла и лекарственных препаратов. Первые изображения этой лилии встречаются на Критских вазах и фресках начиная с 1750 г. до н. э., а затем у древних ассирийцев, египтян, греков и римлян. Древние римляне считали, что после розы нет цветка более прекрасного, чем лилия. От древних римлян лилия попала в завоеванные ими страны Европы. В раннем средневековье ее выращивали монахи в монастырях сначала как лекарственное растение. Белизна ее цветков считалась символом правды, чистоты и непорочности, поэтому у ранних христиан цветки лилии белоснежной стали атрибутами девы Марии-богоматери. Начиная с эпохи итальянского Возрождения и ранней фламандской живописи в картинах на библейский сюжет «Благовещение» изображался архангел Гавриил, приносящий мадонне белую лилию. Карл Линней, давший ботаническое название этой лилии, следовал латинским поэтам, которые начиная с Вергилия (70-19 гг. до н. э.) называли ее белоснежной. С XVI в. в культуру ввели и другие виды лилий. С конца XVIII в. начались работы по гибридизации. В начале XX в. с нахождением в Западном Китае лилии царственной (L. regale) значительно увеличивается число сортов лилий, достигшее теперь 2000. В Восточной Азии лилии разводят как овощную культуру, ради съедобных луковиц. Использовали в пищу и луковицы сибирских видов. В луковице лилии даурской обнаружены сердечные гликозиды. В народной медицине использовали и луковицы саранки, которая, согласно старой сибирской легенде; возникла из сердца казачьего атамана Ермака, погибшего в 1585 г. в бою с ханом Кучумом на реке Иртыш.В роде нотолирион (Notholirion) 5 видов, которые встречаются в Восточной и Западной Азии на высокогорных лугах. Три из них приурочены к Восточной Азии. Нотолирион Томсона (N. thomsonianum) произрастает в сравнительно сухих областях, на солнечных склонах от Центральных Гималаев до Северо-Западного Афганистана. Самый западный вид —- нотолирион Коэ (N. koeiei) —- найден на горных болотах хребта Загрос в Западном Иране. Виды нотолириона- монокарпические растения, которые отличаются от лилий строением луковицы и отчетливо 3-раздельным рыльцем. Луковица нотолириона состоит из влагалищ базальных листьев и низовых чешуй, в пазухах которых образуются дочерние луковички, остающиеся в земле после отмирания растения. Покровными чешуями служат сухие основания листьев. Листья базальные и стеблевые. Цветки трубчатые или колокольчатые, иногда немного зигоморфные, протандричные (табл. 5, 1). В основании сегментов околоцветника имеется нектарная ямка, как у видов рябчика. Семена мелкие, бескрылые. Представители рода номохарис (Nomocharis) встречаются на высокогорных лугах или пастбищах в Восточной Азии. В роде, по-видимому, около 6 видов. Ранее к роду номохарис присоединяли виды с признаками и лилий и рябчика, которые теперь относят к роду лилия (С. Лян, 1980). Луковицы у номохариса такого же строения, как и у лилий, но цветки совершенно плоские, блюдцевидные (табл. 5, 2). Сегменты околоцветника широко распростертые от основания, с пятнышками. Основания внутренних сегментов и тычиночные нити темноокрашенные и образуют заметный «глазок» в центре цветка. Наружные сегменты обычно без нектарников. В основании внутренних сегментов в середине темного пятна имеется короткий нектароносный желобок, от которого веерообразно расходятся складки ткани. Тычиночные нити с мясистым цилиндрическим или кеглевидным основанием и игловидной верхушкой. Столбик почти равен по высоте завязи.Виды номохариса и нотолириона —- декоративные растения.Род рябчик, или фритиллария (Fritillaria), насчитывает около 100 видов, распространенных так же, как лилия, в северном полушарии. Часть видов, близких лилиям, встречается в лесах Восточной Азии. Большая часть наиболее подвинутых видов сосредоточена в Западной Азии. В отличие от остальных родов трибы, имеющих длительный период вегетации, рябчики —- типичные эфемероиды. Луковицы их ежегодно возобновляются, составлены низовыми чешуями, покровных чешуй обычно не имеют. У представителей подрода лилиориза (Liliorhiza) луковицы черепитчатые, рыхлые, с многочисленными мелкими чешуями.
Представители рода номохарис (Nomocharis) встречаются на высокогорных лугах или пастбищах в Восточной Азии. В роде, по-видимому, около 6 видов. Ранее к роду номохарис присоединяли виды с признаками и лилий и рябчика, которые теперь относят к роду лилия (С. Лян, 1980). Луковицы у номохариса такого же строения, как и у лилий, но цветки совершенно плоские, блюдцевидные (табл. 5, 2). Сегменты околоцветника широко распростертые от основания, с пятнышками. Основания внутренних сегментов и тычиночные нити темноокрашенные и образуют заметный «глазок» в центре цветка. Наружные сегменты обычно без нектарников. В основании внутренних сегментов в середине темного пятна имеется короткий нектароносный желобок, от которого веерообразно расходятся складки ткани. Тычиночные нити с мясистым цилиндрическим или кеглевидным основанием и игловидной верхушкой. Столбик почти равен по высоте завязи.Виды номохариса и нотолириона —- декоративные растения.Род рябчик, или фритиллария (Fritillaria), насчитывает около 100 видов, распространенных так же, как лилия, в северном полушарии. Часть видов, близких лилиям, встречается в лесах Восточной Азии. Большая часть наиболее подвинутых видов сосредоточена в Западной Азии. В отличие от остальных родов трибы, имеющих длительный период вегетации, рябчики —- типичные эфемероиды. Луковицы их ежегодно возобновляются, составлены низовыми чешуями, покровных чешуй обычно не имеют. У представителей подрода лилиориза (Liliorhiza) луковицы черепитчатые, рыхлые, с многочисленными мелкими чешуями.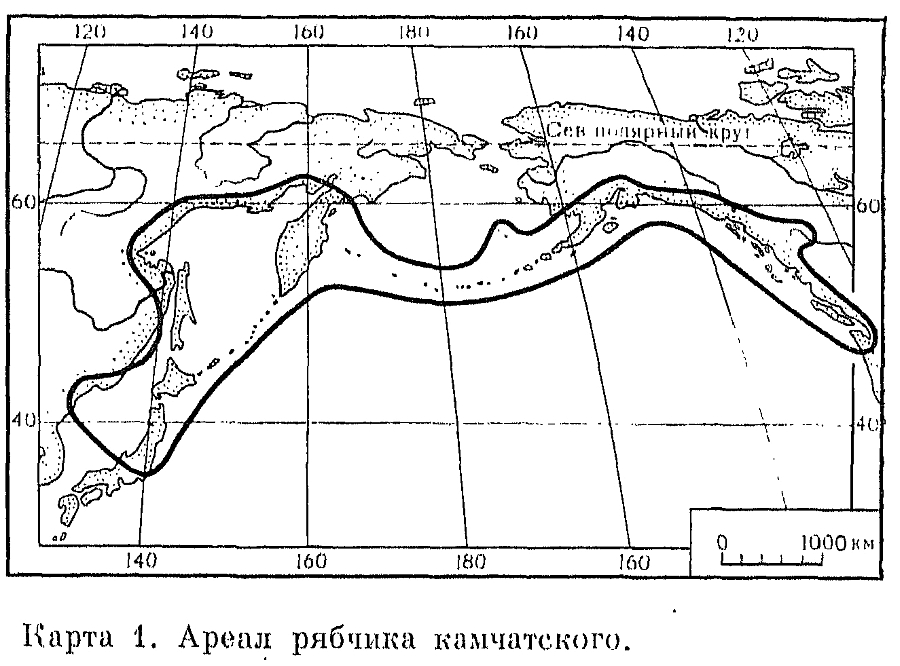 У рябчика камчатского (Fritillaria camschatcensis, рис. 40, 7) и других лесных восточноазиатских видов этой секции чешуи сильно вздутые и похожи на зерна риса. Они легко отделяются от донца и укореняются, имитируя луковички-детки. На самом деле это разбухшие основания низовых чешуй, у которых верхняя часть осталась тонкой и отсохла, о чем свидетельствует рубец. Замещающая луковица у этих видов выносится наружу на толстом столоне. Втягивающих корней у них нет, и луковицы залегают у поверхности почвы. Подавляющее большинство рябчиков имеет луковицы, составленные 1-4 мясистыми широкими чешуями, причем у видов типового подрода луковицы полутуникатные, из 2-3 несросшихся или едва сросшихся чешуй, а у видов подрода петилиум (Petilium), подрода терезия (Theresia) и у рябчика Северцова (Fritillaria sewerzowii) луковица туникатная, из 1-4 массивных, сросшихся целиком или наполовину чешуй (М. В. Баранова, О. И. Захарьева, 1981). У этих растений аридных местообитаний луковица покрыта высохшими чешуями прошлых лет. Многочисленные втягивающие корни способны втянуть луковицу на глубину 25 см и таким образом защитить почку возобновления от высыхания. Замещающая луковица образуется внутри материнской. Стебли у рябчика с очередными или мутовчатыми листьями. Прицветные листья прямостоячие, иногда многочисленные (рябчик императорский —- F. imperialis, табл. 7, 1), иногда спирально закрученные (рябчик русский —- F. ruthenica, рис. 40, 8). Цветки одиночные или их несколько, иногда до 30 (рябчик персидский —- F. persica), собранные в кисть или зонтик (мутовка). Околоцветник актиноморфный или зигоморфный (подрод ринопеталум —- Rhinopetalum), обычно колокольчатый, у некоторых видов с шахматным рисунком. На внутренней стороне сегментов в их основании имеются нектарные ямки (рис. 40, 9, 10, 14, 15), иногда у верхнего сегмента нектарник выпячивается наружу в виде особенно крупного шпорца (подрод ринопеталум, рис. 40, 13); встречаются и бороздчатые нектарники, как у лилий (рябчик камчатский, рис. 40, 2). Пыльники прикрепляются к нити основаниями или несколько выше основания.
У рябчика камчатского (Fritillaria camschatcensis, рис. 40, 7) и других лесных восточноазиатских видов этой секции чешуи сильно вздутые и похожи на зерна риса. Они легко отделяются от донца и укореняются, имитируя луковички-детки. На самом деле это разбухшие основания низовых чешуй, у которых верхняя часть осталась тонкой и отсохла, о чем свидетельствует рубец. Замещающая луковица у этих видов выносится наружу на толстом столоне. Втягивающих корней у них нет, и луковицы залегают у поверхности почвы. Подавляющее большинство рябчиков имеет луковицы, составленные 1-4 мясистыми широкими чешуями, причем у видов типового подрода луковицы полутуникатные, из 2-3 несросшихся или едва сросшихся чешуй, а у видов подрода петилиум (Petilium), подрода терезия (Theresia) и у рябчика Северцова (Fritillaria sewerzowii) луковица туникатная, из 1-4 массивных, сросшихся целиком или наполовину чешуй (М. В. Баранова, О. И. Захарьева, 1981). У этих растений аридных местообитаний луковица покрыта высохшими чешуями прошлых лет. Многочисленные втягивающие корни способны втянуть луковицу на глубину 25 см и таким образом защитить почку возобновления от высыхания. Замещающая луковица образуется внутри материнской. Стебли у рябчика с очередными или мутовчатыми листьями. Прицветные листья прямостоячие, иногда многочисленные (рябчик императорский —- F. imperialis, табл. 7, 1), иногда спирально закрученные (рябчик русский —- F. ruthenica, рис. 40, 8). Цветки одиночные или их несколько, иногда до 30 (рябчик персидский —- F. persica), собранные в кисть или зонтик (мутовка). Околоцветник актиноморфный или зигоморфный (подрод ринопеталум —- Rhinopetalum), обычно колокольчатый, у некоторых видов с шахматным рисунком. На внутренней стороне сегментов в их основании имеются нектарные ямки (рис. 40, 9, 10, 14, 15), иногда у верхнего сегмента нектарник выпячивается наружу в виде особенно крупного шпорца (подрод ринопеталум, рис. 40, 13); встречаются и бороздчатые нектарники, как у лилий (рябчик камчатский, рис. 40, 2). Пыльники прикрепляются к нити основаниями или несколько выше основания.
, Цветки рябчика протогиничные. У рябчика камчатского и других американских видов цветки имеют неприятный запах и опыляются мухами. Восточносредиземноморские виды опыляются шмелями, осами и мелкими мухами. В Калифорнии наблюдается замечательный параллелизм в строении цветков рябчика отогнутого (F. recurva) и лилии малой. Оба вида имеют трубчатые багряно-красные цветки с большим количеством нектара, привлекающего опылителей-колибри. Рябчик императорский имеет признаки цветка, опыляемого птицами (Г. Куглер, 1970). У подавляющего большинства видов рода преобладает семенное размножение и лишь у столонообразующих восточноазиатских видов оно почти полностью подавлено и компенсируется вегетативным размножением с помощью большого числа (до 50) легко опадающих чешуй.Рябчики используют как декоративные и лекарственные растения. Многие виды ядовиты, так как содержат алкалоиды. Несмотря на горечь, луковицы некоторых видов съедобны. Так, луковицы рябчика камчатского, встречающегося по обоим побережьям (азиатскому и американскому) Тихого океана, служили пищей камчадалам и индейцам. Камчадалы, называвшие растения «сараной», выкапывали луковицы мотыгой или забирали их из кладовых полевки-экономки (Microtus oeconomus). Сушеные чешуи луковиц, нанизанные на веревки, вывозили с Камчатки, где растения росли в изобилии, в Америку и продавали индейцам, которые называли их «северо-западным рисом».Триба тюльпановых (Tulipeae), более подвинутая, чем триба лилейных, включает 2 рода, у представителей которых луковицы ежегодно возобновляются, и составлены 1 —- 5 низовыми замкнутыми чешуями.Род тюльпан (Tulipa) насчитывает, вероятно, около 100 видов, но некоторые авторы доводят их число до 140 (З. М. Силина, 1977). Тюльпаны распространены в Евразии и Северной Африке, в областях с жарким и сухим летом и небольшим количеством осадков весной и осенью, в пустынях, полупустынях и степях, редко в лесу. Они растут на равнинах и во всех поясах гор, но редко вблизи ледников, на различных почвах: от плотного лесса до сыпучих песков, на нейтральных до засоленных, но не кислых почвах. Луковицы тюльпанов залегают на глубине до 45 см и более. Они состоят из 2-5 мясистых запасающих чешуй, одетых специализированными покровными кожистыми или пленчатыми чешуями, которые с внутренней стороны бывают голыми, прижато-щетинистыми или шерстисто-войлочными. У ювенильных (до первого цветения) растений ежегодно формирующиеся замещающие луковицы выносятся за пределы материнской луковицы и углубляются при помощи вертикальных (у некоторых видов горизонтальных) полых столонов, внутри которых они находятся (рис. 41, 9, 10). У некоторых видов сохраняется цепочка отмерших луковиц, которые иногда ошибочно принимают за «четковидные луковицы» (рис. 41, 8). Кроме замещающей луковицы, может образоваться 1-2 дочерних, при этом или одна из них, или обе в столонах (рис. 41). Стебель тюльпанов с 2-4 (реже с 1 и до 12) листьями и с 1-2 (реже 3-15) прямостоячими цветками, хотя бутоны у некоторых видов бывают поникшие. Сегменты околоцветника без нектарников, опадают при плодах. С изменением температуры среды происходит движение сегментов (термонастии). При повышении температуры цветки широко раскрываются благодаря расширению клеток на внутренней стороне сегментов, при понижении температуры закрываются вследствие расширения клеток наружной стороны. Для тюльпанов характерна многоцветная окраска цветов (полихроизм). Так, например, даже в одной популяции степного тюльпана Шренка (Т. schrenkii) встречаются растения с цветками самой разной окраски —- белой, желтой, розовой, красной, фиолетовой, с пятном, при основании черным или желтым или без пятна. Тычинки часто с расширенными к основанию, иногда толстыми нитями, которые оканчиваются острием, входящим в углубление основания пыльника, отчего пыльники вращаются вокруг оси. Рыльце 3-лопастное, чаще сидячее; столбик более или менее длинный.
Цветки рябчика протогиничные. У рябчика камчатского и других американских видов цветки имеют неприятный запах и опыляются мухами. Восточносредиземноморские виды опыляются шмелями, осами и мелкими мухами. В Калифорнии наблюдается замечательный параллелизм в строении цветков рябчика отогнутого (F. recurva) и лилии малой. Оба вида имеют трубчатые багряно-красные цветки с большим количеством нектара, привлекающего опылителей-колибри. Рябчик императорский имеет признаки цветка, опыляемого птицами (Г. Куглер, 1970). У подавляющего большинства видов рода преобладает семенное размножение и лишь у столонообразующих восточноазиатских видов оно почти полностью подавлено и компенсируется вегетативным размножением с помощью большого числа (до 50) легко опадающих чешуй.Рябчики используют как декоративные и лекарственные растения. Многие виды ядовиты, так как содержат алкалоиды. Несмотря на горечь, луковицы некоторых видов съедобны. Так, луковицы рябчика камчатского, встречающегося по обоим побережьям (азиатскому и американскому) Тихого океана, служили пищей камчадалам и индейцам. Камчадалы, называвшие растения «сараной», выкапывали луковицы мотыгой или забирали их из кладовых полевки-экономки (Microtus oeconomus). Сушеные чешуи луковиц, нанизанные на веревки, вывозили с Камчатки, где растения росли в изобилии, в Америку и продавали индейцам, которые называли их «северо-западным рисом».Триба тюльпановых (Tulipeae), более подвинутая, чем триба лилейных, включает 2 рода, у представителей которых луковицы ежегодно возобновляются, и составлены 1 —- 5 низовыми замкнутыми чешуями.Род тюльпан (Tulipa) насчитывает, вероятно, около 100 видов, но некоторые авторы доводят их число до 140 (З. М. Силина, 1977). Тюльпаны распространены в Евразии и Северной Африке, в областях с жарким и сухим летом и небольшим количеством осадков весной и осенью, в пустынях, полупустынях и степях, редко в лесу. Они растут на равнинах и во всех поясах гор, но редко вблизи ледников, на различных почвах: от плотного лесса до сыпучих песков, на нейтральных до засоленных, но не кислых почвах. Луковицы тюльпанов залегают на глубине до 45 см и более. Они состоят из 2-5 мясистых запасающих чешуй, одетых специализированными покровными кожистыми или пленчатыми чешуями, которые с внутренней стороны бывают голыми, прижато-щетинистыми или шерстисто-войлочными. У ювенильных (до первого цветения) растений ежегодно формирующиеся замещающие луковицы выносятся за пределы материнской луковицы и углубляются при помощи вертикальных (у некоторых видов горизонтальных) полых столонов, внутри которых они находятся (рис. 41, 9, 10). У некоторых видов сохраняется цепочка отмерших луковиц, которые иногда ошибочно принимают за «четковидные луковицы» (рис. 41, 8). Кроме замещающей луковицы, может образоваться 1-2 дочерних, при этом или одна из них, или обе в столонах (рис. 41). Стебель тюльпанов с 2-4 (реже с 1 и до 12) листьями и с 1-2 (реже 3-15) прямостоячими цветками, хотя бутоны у некоторых видов бывают поникшие. Сегменты околоцветника без нектарников, опадают при плодах. С изменением температуры среды происходит движение сегментов (термонастии). При повышении температуры цветки широко раскрываются благодаря расширению клеток на внутренней стороне сегментов, при понижении температуры закрываются вследствие расширения клеток наружной стороны. Для тюльпанов характерна многоцветная окраска цветов (полихроизм). Так, например, даже в одной популяции степного тюльпана Шренка (Т. schrenkii) встречаются растения с цветками самой разной окраски —- белой, желтой, розовой, красной, фиолетовой, с пятном, при основании черным или желтым или без пятна. Тычинки часто с расширенными к основанию, иногда толстыми нитями, которые оканчиваются острием, входящим в углубление основания пыльника, отчего пыльники вращаются вокруг оси. Рыльце 3-лопастное, чаще сидячее; столбик более или менее длинный.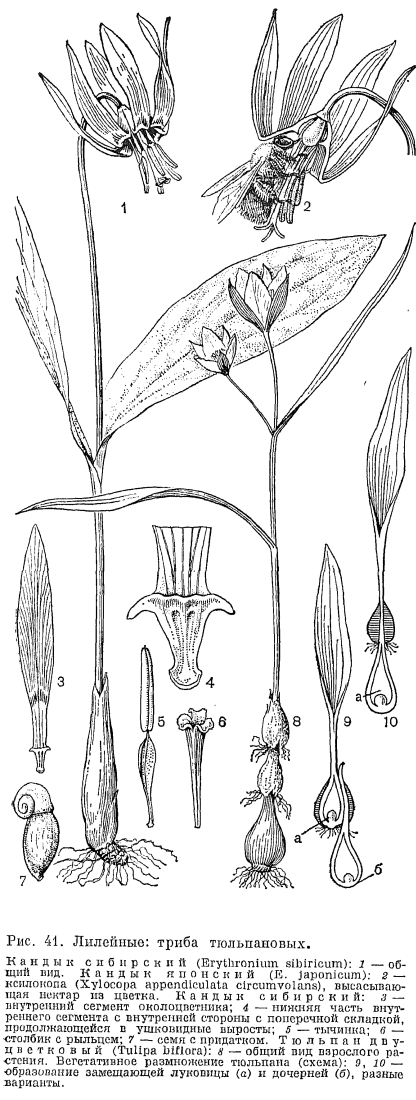 У тюльпанов ярко выражена протандрия. Цветки тюльпанов опыляются мелкими пчелами, мухами, собирающими пыльцу. Нектарники, подобные нектарникам безвременника, имеются лишь у тюльпана лесного (T. sylvestris, Э. Дауманн, 1970). Весенние эфемероиды тюльпаны в период массового цветения покрывают открытые горные или степные склоны красными, желтыми, пестрыми коврами. Прямостоячие коробочки тюльпанов содержат многочисленные плоские, треугольные коричневые семена, распространяемые по способу баллистов, что характерно для растений открытых пространств. Некоторые виды размножаются вегетативно дочерними луковичками преимущественно в ювенильный период. Тюльпаны —- прекрасные декоративные растения. Первые культурные тюльпаны попали в Западную Европу в XVI в. из Константинополя. Их называли турецкими тюльпанами, прародителем их был тюльпан Шренка, привозимый из Кафы (нынешней Феодосии), входившей тогда в состав Османской империи (М. Хоог, 1973). Голландия стала страной промышленного выращивания культурных сортов тюльпанов. В начале XIX в. стало известно о существовании настоящего очага видового разнообразия тюльпанов в Средней Азии (около 75 видов). Заслуга в интродукции многих декоративных растений Средней Азии и тюльпанов в первую очередь принадлежит директору ботанического сада в Санкт-Петербурге Эдуарду Регелю. В настоящее время насчитывается 2900 промышленных сортов тюльпанов, объединенных в 15 классов; 3 класса из них составляют разновидности и гибриды замечательных среднеазиатских видов: тюльпана Кауфмана (T. kaufmanniana), тюльпана Фостера (T. fosteriana) и тюльпана Грейга (T. greigii), который считается самым красивым тюльпаном в мире.Род кандык, или эритрониум (Erythronium), насчитывает 24 вида, из которых 15 произрастают на западе, 5 —- на востоке Северной Америки, а 4 вида —- евразиатских. Эритрониумы —- лесные горные растения, они встречаются от предгорий до альпийских лугов в хвойных, смешанных и лиственных лесах, реже на высокогорных лугах. Луковица кандыка сибирского (E. sibiricum) и кандыка кавказского (E. caucasicum) состоит из одной запасающей чешуи, образовавшейся из 3 сросшихся на ранних стадиях онтогенеза между собой и с цветоносом в одну замкнутую чешую (Н. И. Шорина и А. Г. Куклина, 1976). Покровные чешуи луковицы образуются из высохших запасающих чешуй. У основания луковицы кандыка сибирского сохраняются донца луковиц прошлых лет, образуя членистое (до 6 члеников) корневище (Г. В. Скакунов, 1974, рис. 41, 1). У ювенильных растений замещающая луковица углубляется в почву при помощи шпорообразно удлиняющегося влагалища низового листа, обладающего положительным геотропизмом.Эритрониумы —- ранневесенние эфемероиды. Весной появляется стебель с 2-3 часто пятнистыми листьями и обычно одним поникшим розовым, лиловым, белым или желтым цветком. Сегменты околоцветника сближены в нижней части и образуют трубку, а в верхней отогнуты кверху, как у цикламена. В пасмурную погоду и с наступлением сумерек сегменты опускаются, предохраняя пыльцу от намокания. Нижняя часть сегментов обычно окрашена иначе, а на месте отгиба сегментов имеются различные метки: пятна, крапинки, штрихи. У некоторых видов в основании внутренних сегментов имеется поперечная, разделенная на 4 лопасти складка, которая продолжается с каждой стороны сегмента в виде ушковидных отростков (рис. 41, 4). Тычинки эритрониумов имеют подвижные пыльники, как у тюльпанов, и нити, иногда сильно уплощенные (кандык сибирский и кандык европейский, или собачий зуб, —- E. denscanis, рис. 41, 5); у некоторых видов тычинки обоих кругов разной длины (кандык японский —- E. japonicum). Соотношение длины зрелого столбика и тычинок варьирует у разных видов.Цветок эритрониумов имеет интересные приспособления к насекомоопылению, что особенно ярко выражено у близкого к кандыку сибирскому кандыка японского (Ф. Утех и С. Кавано, 1975). У этого растения нектар в большом количестве выделяется секреторной тканью в основании сегментов. Для того чтобы нектар не вытекал из опрокинутого околоцветника, существует система приспособлений, образующая как бы колпак вокруг завязи. Ниже лопастей складки, по краям основания сегментов имеются продольные вздутия, благодаря которым колпак вокруг завязи плотно прижат к ее стенкам и не позволяет вытекать нектару. Чтобы получить нектар, надо отстранить сегменты и нити тычинок от завязи, с чем успешно справляется ксилокопа —- самая крупная пчела японской фауны перепончатокрылых. Лиловые отвернутые части сегментов околоцветника сильно отражают ультрафиолетовые лучи и разительно контрастируют с пурпурным зубчатым ободком в зеве трубки, таким заметным в видимом спектре. В сочетании эти свойства околоцветника указывают путь насекомым с трехцветным зрением. Действия пчелы точные и быстрые. Удерживаясь на тычинках и столбике, она головкой отстраняет сегменты околоцветника, сжимающие завязь, и открывает выход нектару, который высасывает своим хоботком (рис. 41, 2). В это время волосистая поверхность тела и лапок пчелы покрывается пыльцой. Эффект усиливается благодаря тому, что пыльники, открывающиеся двумя продольными щелями, вращаются на оси, а также благодаря различной длине тычиночных нитей, в результате чего образовавшаяся зона пыльников почти вдвое превышает длину одного пыльника. Цветки кандыка японского слабо протандричны. Лопасти рыльца раскрываются только тогда, когда длина рыльца превысит длину тычинок с созревшими пыльниками, что исключает самоопыление.Коробочки прямостоячие, с небольшим числом семян, стебли при плодах полегающие. У евразиатских видов семена продолговато-яйцевидные, желтые, с хорошо выраженным бесцветным придатком в области халазы (рис. 41, 7). Придаток, содержащий жирные масла, поедают муравьи, которые способствуют распространению семян. По своей морфологической природе придаток представляет ариллоид, вырост наружного интегумента (Г. А. Комар, 1978). Основной способ размножения видов —- семенной, некоторые размножаются вегетативно. Луковицы некоторых видов используют в пищу и как лекарственные средства. Все виды рода —- ранневесенние декоративные растения.Последней, наиболее подвинутой трибой подсемейства лилейных является триба гейджиевых (Gageeae). Это приоритетное название трибы (Г. Руи, 1910), ранее известной как триба ллойдиевых (Lloydieae, Ф. Буксбаум, 1937). В трибе гейджиевых 2 рода, которые характеризуются ежегодно возобновляющейся луковицей, состоящей из одной замкнутой низовой чешуи, покрытой сухими влагалищами прошлогодних листьев.
У тюльпанов ярко выражена протандрия. Цветки тюльпанов опыляются мелкими пчелами, мухами, собирающими пыльцу. Нектарники, подобные нектарникам безвременника, имеются лишь у тюльпана лесного (T. sylvestris, Э. Дауманн, 1970). Весенние эфемероиды тюльпаны в период массового цветения покрывают открытые горные или степные склоны красными, желтыми, пестрыми коврами. Прямостоячие коробочки тюльпанов содержат многочисленные плоские, треугольные коричневые семена, распространяемые по способу баллистов, что характерно для растений открытых пространств. Некоторые виды размножаются вегетативно дочерними луковичками преимущественно в ювенильный период. Тюльпаны —- прекрасные декоративные растения. Первые культурные тюльпаны попали в Западную Европу в XVI в. из Константинополя. Их называли турецкими тюльпанами, прародителем их был тюльпан Шренка, привозимый из Кафы (нынешней Феодосии), входившей тогда в состав Османской империи (М. Хоог, 1973). Голландия стала страной промышленного выращивания культурных сортов тюльпанов. В начале XIX в. стало известно о существовании настоящего очага видового разнообразия тюльпанов в Средней Азии (около 75 видов). Заслуга в интродукции многих декоративных растений Средней Азии и тюльпанов в первую очередь принадлежит директору ботанического сада в Санкт-Петербурге Эдуарду Регелю. В настоящее время насчитывается 2900 промышленных сортов тюльпанов, объединенных в 15 классов; 3 класса из них составляют разновидности и гибриды замечательных среднеазиатских видов: тюльпана Кауфмана (T. kaufmanniana), тюльпана Фостера (T. fosteriana) и тюльпана Грейга (T. greigii), который считается самым красивым тюльпаном в мире.Род кандык, или эритрониум (Erythronium), насчитывает 24 вида, из которых 15 произрастают на западе, 5 —- на востоке Северной Америки, а 4 вида —- евразиатских. Эритрониумы —- лесные горные растения, они встречаются от предгорий до альпийских лугов в хвойных, смешанных и лиственных лесах, реже на высокогорных лугах. Луковица кандыка сибирского (E. sibiricum) и кандыка кавказского (E. caucasicum) состоит из одной запасающей чешуи, образовавшейся из 3 сросшихся на ранних стадиях онтогенеза между собой и с цветоносом в одну замкнутую чешую (Н. И. Шорина и А. Г. Куклина, 1976). Покровные чешуи луковицы образуются из высохших запасающих чешуй. У основания луковицы кандыка сибирского сохраняются донца луковиц прошлых лет, образуя членистое (до 6 члеников) корневище (Г. В. Скакунов, 1974, рис. 41, 1). У ювенильных растений замещающая луковица углубляется в почву при помощи шпорообразно удлиняющегося влагалища низового листа, обладающего положительным геотропизмом.Эритрониумы —- ранневесенние эфемероиды. Весной появляется стебель с 2-3 часто пятнистыми листьями и обычно одним поникшим розовым, лиловым, белым или желтым цветком. Сегменты околоцветника сближены в нижней части и образуют трубку, а в верхней отогнуты кверху, как у цикламена. В пасмурную погоду и с наступлением сумерек сегменты опускаются, предохраняя пыльцу от намокания. Нижняя часть сегментов обычно окрашена иначе, а на месте отгиба сегментов имеются различные метки: пятна, крапинки, штрихи. У некоторых видов в основании внутренних сегментов имеется поперечная, разделенная на 4 лопасти складка, которая продолжается с каждой стороны сегмента в виде ушковидных отростков (рис. 41, 4). Тычинки эритрониумов имеют подвижные пыльники, как у тюльпанов, и нити, иногда сильно уплощенные (кандык сибирский и кандык европейский, или собачий зуб, —- E. denscanis, рис. 41, 5); у некоторых видов тычинки обоих кругов разной длины (кандык японский —- E. japonicum). Соотношение длины зрелого столбика и тычинок варьирует у разных видов.Цветок эритрониумов имеет интересные приспособления к насекомоопылению, что особенно ярко выражено у близкого к кандыку сибирскому кандыка японского (Ф. Утех и С. Кавано, 1975). У этого растения нектар в большом количестве выделяется секреторной тканью в основании сегментов. Для того чтобы нектар не вытекал из опрокинутого околоцветника, существует система приспособлений, образующая как бы колпак вокруг завязи. Ниже лопастей складки, по краям основания сегментов имеются продольные вздутия, благодаря которым колпак вокруг завязи плотно прижат к ее стенкам и не позволяет вытекать нектару. Чтобы получить нектар, надо отстранить сегменты и нити тычинок от завязи, с чем успешно справляется ксилокопа —- самая крупная пчела японской фауны перепончатокрылых. Лиловые отвернутые части сегментов околоцветника сильно отражают ультрафиолетовые лучи и разительно контрастируют с пурпурным зубчатым ободком в зеве трубки, таким заметным в видимом спектре. В сочетании эти свойства околоцветника указывают путь насекомым с трехцветным зрением. Действия пчелы точные и быстрые. Удерживаясь на тычинках и столбике, она головкой отстраняет сегменты околоцветника, сжимающие завязь, и открывает выход нектару, который высасывает своим хоботком (рис. 41, 2). В это время волосистая поверхность тела и лапок пчелы покрывается пыльцой. Эффект усиливается благодаря тому, что пыльники, открывающиеся двумя продольными щелями, вращаются на оси, а также благодаря различной длине тычиночных нитей, в результате чего образовавшаяся зона пыльников почти вдвое превышает длину одного пыльника. Цветки кандыка японского слабо протандричны. Лопасти рыльца раскрываются только тогда, когда длина рыльца превысит длину тычинок с созревшими пыльниками, что исключает самоопыление.Коробочки прямостоячие, с небольшим числом семян, стебли при плодах полегающие. У евразиатских видов семена продолговато-яйцевидные, желтые, с хорошо выраженным бесцветным придатком в области халазы (рис. 41, 7). Придаток, содержащий жирные масла, поедают муравьи, которые способствуют распространению семян. По своей морфологической природе придаток представляет ариллоид, вырост наружного интегумента (Г. А. Комар, 1978). Основной способ размножения видов —- семенной, некоторые размножаются вегетативно. Луковицы некоторых видов используют в пищу и как лекарственные средства. Все виды рода —- ранневесенние декоративные растения.Последней, наиболее подвинутой трибой подсемейства лилейных является триба гейджиевых (Gageeae). Это приоритетное название трибы (Г. Руи, 1910), ранее известной как триба ллойдиевых (Lloydieae, Ф. Буксбаум, 1937). В трибе гейджиевых 2 рода, которые характеризуются ежегодно возобновляющейся луковицей, состоящей из одной замкнутой низовой чешуи, покрытой сухими влагалищами прошлогодних листьев.
, Род гейджия, или гусиный лук (Gagea), насчитывает около 70 видов, распространенных в умеренных областях Евразии и Северной Африки, от лесотундры и ледников в горах до полупустынь. Гусиные луки —- ранневесенние эфемероиды. Их желтые звездчатые цветки покрывают весной горные луга, щебнистые склоны и трещины скал, встречаются в степи, иногда на засоленной почве и на известняках, отдельными куртинами в лиственных лесах и на газонах в парках или, как сорняки, в посевах. Это самые мелкие растения в подсемействе лилейных —- высотой от 3 до 35 см. Они имеют одну луковицу или 2-3 в том случае, если у материнской луковицы образуется 1 или 2 луковички-детки, сохраняющиеся при материнском растении. У основания материнской луковицы иногда за ряд лет скапливается множество мелких луковичек-деток, образующих гнездо разновозрастных луковиц. Покровные чешуи сетчато-волокнистые, пленчатые или кожистые. У некоторых видов подрода платиспермум (Platyspermum), обитающих в сравнительно сухих местах, имеются корни двоякого рода: обычные нитевидные придаточные корни, идущие от середины донца луковицы вертикально вниз (положительный геотропизм), и жесткие склерифицированные корни по краям донца, идущие сначала вниз, а затем горизонтально и вверх (нулевой или отрицательный геотропизм, рис. 42, 1). Они оплетают луковицу и вместе с песчинками между ними образуют вокруг нее как бы капсулу, которая, возможно, защищает луковицу от высыхания в летний период. Базальные листья (1 или 2) обычно превышают соцветие, плоские, с килем или дудчатые (полые внутри). Подсоцветных листьев 1-3(5), реже стебель равномерно облиственный. Соцветия зонтиковидные, малоцветковые. Цветки часто на неравных цветоножках. Сегменты околоцветника снаружи зеленые, внутри желтые, в основании с нектарной ямкой. Тычинки так же, как у видов тюльпана и эритрониума, имеют пыльники, прикрепленные основаниями, подвижные (рис. 42, 2). Цветки слабо протогиничные, без запаха. Опылителей —- маленьких мух, жуков и пчел —- привлекает нектар, который скапливается в виде капель между основаниями сегментов и тычиночных нитей. Семена плоские, треугольные (подрод платиспермум) или яйцевидные (типовой подрод), причем у некоторых видов, например у гусиного лука желтого (G. lutea, табл. 8, 7) и гусиного лука сетчатого (G. reticulata), семена с элайосомами (рис. 42, 4). По своей морфологической природе это одна из разновидностей ариллоида —- строфиола, т. е. разрастание ткани наружного интегумента семени вдоль семенного шва. Гусиные луки интенсивно размножаются и вегетативно, с помощью луковичек, которые образуются на донце луковицы, в пазухах базальных или стеблевых листьев или иногда на месте бутонов. У одних видов усиленное вегетативное размножение происходит до первого цветения растения, у других способность к вегетативному размножению сохраняется и во взрослом состоянии. К роду гусиный лук близок род ллойдия (Lloydia), насчитывающий примерно 20 видов. Ллойдии —- высокогорные растения (ореофиты). Большинство видов встречается в Восточной Азии (7 видов в Китае), несколько —- в Восточном Средиземноморье. Ареал ллойдии поздней (L. serotina) охватывает территорию от гор Уэльса в Англии, горы Евразии до запада Северной Америки и Арктическую провинцию. В. Грейтер (1975) склонен принимать род ллойдия как монотипный, с одним видом ллойдия поздняя. По облику ллойдии похожи на гусиные луки, но луковица у них продолговатая и окутана волокнистыми влагалищами прошлогодних листьев. У основания луковицы могут сохраняться донца прошлогодних луковиц, образуя членистое корневище. Цветки одиночные или по 2-3, воронковидные. Сегменты околоцветника беловатые, с пурпуровыми жилками и темными основаниями, опушенные по жилкам (ллойдия тибетская —- L. tibetica, рис. 42, 5, 6), или с нектарниками (ллойдия поздняя), или без опушения и нектарников. Тычинки с подвижными пыльниками, как у гусиных луков. Нектарники ллойдии поздней представляют собой поперечную складку выше основания сегмента, над которой в углублении выделяется и задерживается нектар. Протандричные цветки ллойдии поздней со свободно лежащим нектаром опыляются короткохоботковыми насекомыми. Стебли при плодах не полегают. Коробочка растрескивается в верхней части гнезд, рассеивая плоские, полулунной формы семена по способу баллистов.К подсемейству лилейных, возможно, относится и монотипная триба медеоловых (Medeoleae) с единственным родом и видом медеола вирджинская (Medeola virginiana). Медеола растет в лесах в восточной части Северной Америки и по облику похожа на триллиум (Trillium) или вороний глаз (Paris). Вот почему эти роды объединяли ранее в трибу парисовых (Parideae) или в семейство триллиевые (Trilliaceae), пока Р. Берг (1962) не установил, что медеолу гораздо правильнее отнести к подсемейству лилейных. У медеолы симподиальный, ежегодно замещающийся клубень (уникальный тип в пределах семейства). Надземный стебель имеет 2 ложные мутовки листьев и верхушечное соцветие —- зонтик, обычно из 3 цветков на поникших цветоножках, которые становятся прямостоячими при плодах. Цветки мелкие, без запаха. Сегменты околоцветника одинаковые, зеленоватые, тычинки красноватые, завязь с 3 рыльцами. Пыльники находятся ниже широко раскинутых рылец и самоопыление не происходит. Плод —- ягода; семена распространяются, по-видимому, птицами.
Род гейджия, или гусиный лук (Gagea), насчитывает около 70 видов, распространенных в умеренных областях Евразии и Северной Африки, от лесотундры и ледников в горах до полупустынь. Гусиные луки —- ранневесенние эфемероиды. Их желтые звездчатые цветки покрывают весной горные луга, щебнистые склоны и трещины скал, встречаются в степи, иногда на засоленной почве и на известняках, отдельными куртинами в лиственных лесах и на газонах в парках или, как сорняки, в посевах. Это самые мелкие растения в подсемействе лилейных —- высотой от 3 до 35 см. Они имеют одну луковицу или 2-3 в том случае, если у материнской луковицы образуется 1 или 2 луковички-детки, сохраняющиеся при материнском растении. У основания материнской луковицы иногда за ряд лет скапливается множество мелких луковичек-деток, образующих гнездо разновозрастных луковиц. Покровные чешуи сетчато-волокнистые, пленчатые или кожистые. У некоторых видов подрода платиспермум (Platyspermum), обитающих в сравнительно сухих местах, имеются корни двоякого рода: обычные нитевидные придаточные корни, идущие от середины донца луковицы вертикально вниз (положительный геотропизм), и жесткие склерифицированные корни по краям донца, идущие сначала вниз, а затем горизонтально и вверх (нулевой или отрицательный геотропизм, рис. 42, 1). Они оплетают луковицу и вместе с песчинками между ними образуют вокруг нее как бы капсулу, которая, возможно, защищает луковицу от высыхания в летний период. Базальные листья (1 или 2) обычно превышают соцветие, плоские, с килем или дудчатые (полые внутри). Подсоцветных листьев 1-3(5), реже стебель равномерно облиственный. Соцветия зонтиковидные, малоцветковые. Цветки часто на неравных цветоножках. Сегменты околоцветника снаружи зеленые, внутри желтые, в основании с нектарной ямкой. Тычинки так же, как у видов тюльпана и эритрониума, имеют пыльники, прикрепленные основаниями, подвижные (рис. 42, 2). Цветки слабо протогиничные, без запаха. Опылителей —- маленьких мух, жуков и пчел —- привлекает нектар, который скапливается в виде капель между основаниями сегментов и тычиночных нитей. Семена плоские, треугольные (подрод платиспермум) или яйцевидные (типовой подрод), причем у некоторых видов, например у гусиного лука желтого (G. lutea, табл. 8, 7) и гусиного лука сетчатого (G. reticulata), семена с элайосомами (рис. 42, 4). По своей морфологической природе это одна из разновидностей ариллоида —- строфиола, т. е. разрастание ткани наружного интегумента семени вдоль семенного шва. Гусиные луки интенсивно размножаются и вегетативно, с помощью луковичек, которые образуются на донце луковицы, в пазухах базальных или стеблевых листьев или иногда на месте бутонов. У одних видов усиленное вегетативное размножение происходит до первого цветения растения, у других способность к вегетативному размножению сохраняется и во взрослом состоянии. К роду гусиный лук близок род ллойдия (Lloydia), насчитывающий примерно 20 видов. Ллойдии —- высокогорные растения (ореофиты). Большинство видов встречается в Восточной Азии (7 видов в Китае), несколько —- в Восточном Средиземноморье. Ареал ллойдии поздней (L. serotina) охватывает территорию от гор Уэльса в Англии, горы Евразии до запада Северной Америки и Арктическую провинцию. В. Грейтер (1975) склонен принимать род ллойдия как монотипный, с одним видом ллойдия поздняя. По облику ллойдии похожи на гусиные луки, но луковица у них продолговатая и окутана волокнистыми влагалищами прошлогодних листьев. У основания луковицы могут сохраняться донца прошлогодних луковиц, образуя членистое корневище. Цветки одиночные или по 2-3, воронковидные. Сегменты околоцветника беловатые, с пурпуровыми жилками и темными основаниями, опушенные по жилкам (ллойдия тибетская —- L. tibetica, рис. 42, 5, 6), или с нектарниками (ллойдия поздняя), или без опушения и нектарников. Тычинки с подвижными пыльниками, как у гусиных луков. Нектарники ллойдии поздней представляют собой поперечную складку выше основания сегмента, над которой в углублении выделяется и задерживается нектар. Протандричные цветки ллойдии поздней со свободно лежащим нектаром опыляются короткохоботковыми насекомыми. Стебли при плодах не полегают. Коробочка растрескивается в верхней части гнезд, рассеивая плоские, полулунной формы семена по способу баллистов.К подсемейству лилейных, возможно, относится и монотипная триба медеоловых (Medeoleae) с единственным родом и видом медеола вирджинская (Medeola virginiana). Медеола растет в лесах в восточной части Северной Америки и по облику похожа на триллиум (Trillium) или вороний глаз (Paris). Вот почему эти роды объединяли ранее в трибу парисовых (Parideae) или в семейство триллиевые (Trilliaceae), пока Р. Берг (1962) не установил, что медеолу гораздо правильнее отнести к подсемейству лилейных. У медеолы симподиальный, ежегодно замещающийся клубень (уникальный тип в пределах семейства). Надземный стебель имеет 2 ложные мутовки листьев и верхушечное соцветие —- зонтик, обычно из 3 цветков на поникших цветоножках, которые становятся прямостоячими при плодах. Цветки мелкие, без запаха. Сегменты околоцветника одинаковые, зеленоватые, тычинки красноватые, завязь с 3 рыльцами. Пыльники находятся ниже широко раскинутых рылец и самоопыление не происходит. Плод —- ягода; семена распространяются, по-видимому, птицами.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.
.