- Подсемейство антериковые или венечниковые (Anthericoideae)
- Подсемейство антериковые или венечниковые (Anthericoideae)
-
В подсемействе антериковых (Anthericoideae) известны 31 род и около 630 видов; из них 27 родов распространено в Старом Свете, 1 (антерикум) встречается также и в Америке, 4 рода произрастают только на Американском континенте —- от юга США (штаты Юта и Аризона) до Перу и Чили. В этом подсемействе различают 5 триб. Самой большой из них является триба антериковых (Anthericeae), состоящая из 14 родов, представленных многолетними травами и очень редко небольшими кустарниковидными и полукустарниковидными растениями. Цветки у них звездчатые, колесовидные, редко воронковидные или колокольчатые, со свободными или сросшимися при основании, редко до половины сегментами. Тычинок иногда 3 (анемаррена —- Anemarrhena). Гнезда завязи обычно с многочисленными (до 30), реже несколькими или 2 (роды анемаррена и алекторурус —- Alectorurus) семязачатками. Плод —- коробочка.Наиболее известным родом трибы антериковых является хлорофитум (Chlorophytum). Он включает около 220 видов, большинство которых распространено в тропической Африке, 20 видов встречаются в Южной Африке, около 15 —- на Мадагаскаре, по 1 —- на юге Аравийского полуострова и на острове Сокотра, около 15 видов —- в Индии, несколько видов —- в Таиланде, Индокитае, на полуострове Малакка, в Южном Китае, Австралии и Тасмании. Виды хлорофитума преимущественно мезофильные растения, обитающие во влажных, обычно тенистых местах. На востоке Южной Африки в субтропических вечнозеленых муссонных лесах обычен хлорофитум хохолковый (С. comosum), являющийся важным компонентом их травяного покрова. Это растение имеет довольно необычный облик, благодаря тому что в его соцветии на верхушке цветоноса и боковых ветвей, а нередко и на других частях развиваются новые растения с розетками листьев и короткими корнями. При полегании цветоноса розетки укореняются, и таким путем у этого вида происходит быстрое вегетативное размножение. Хлорофитум хохолковый хорошо знаком читателям, так как он широко распространен в комнатной культуре. Некоторые виды хлорофитума произрастают в аридных районах Южной Африки. В качестве примера можно привести хлорофитум липкий (С. viscosum) —- очень жесткое, опушенное железистыми волосками растение, обитающее по пескам в пустыне Намиб и на полупустынном плоскогорье Намакваленд, а также хлорофитум аридный (С. aridum), встречающийся в кустарниковых саваннах Трансвааля.
 Виды хлорофитума —- многолетние травы с ползучим корневищем, обычно тонкими, но у концов часто клубневидно утолщенными корнями, с розетками листьев и с центральным простым или разветвленным цветоносом. Самый высокий цветонос (до 2 м), а также наиболее длинные (до 140 см) слабосуккулентные листья имеет похожий на алоэ хлорофитум Крука (С. krookianum), встречающийся по сырым местам в восточных районах Южной Африки (рис. 72). Листья у хлорофитумов от линейных до широколанцетных, по краю обычно бахромчатые, а при основании нередко суженные в черешок. Мелкие, белые, реже зеленоватые или розоватые цветки собраны в 1 —- 6-цветковые пучки, расположенные в пазухе прицветников и образующие рыхлые или более или менее густые сложные кисти. Каждый цветок такого пучка сидит непосредственно на оси соцветия в пазухе прицветника. Пучки представляют собой редуцированные боковые ветви соцветия, оси которых подверглись полной редукции. У некоторых видов (например, у хлорофитума капского —- С. capense) иногда развиваются очень короткие оси и цветки на них расположены по спирали. Околоцветник у хлорофитумов со свободными сегментами, обычно расположенными звездчато или отклоненными вниз. Тычинки с крупными пыльниками, прикрепляющимися к нити основанием, при увядании изгибаются назад. Гинецей состоит из 3 плодолистиков, но у хлорофитума клубневого (С. tuberosum), распространенного в тропической Африке и Индии, он образован только 2 плодолистиками. В каждом гнезде завязи имеется от 6 до 30 семязачатков. Коробочка 3-гранная, часто глубоколопастная. Семена крупные, диаметром 2 —- 4 мм, плоские, округлые, черные и блестящие.Цветет хлорофитум в течение всего года, но разные его виды —- в различное время. Цветки в пучках раскрываются последовательно, и первым распускается нижний бутон. При неблагоприятных для цветения условиях рост бутонов может задержаться на продолжительное время, и если они вновь начнут расти и распускаться, то в одних и тех же пучках можно видеть и зрелые плоды, и раскрытые и увядшие цветки, и бутоны. Цветоножки при цветении направлены косо вверх и не изменяют своего положения при плодах. Цветки опыляются пчелами, цветочными мухами и другими насекомыми, привлекаемыми пыльцой и нектаром. Несколько видов хлорофитума разводят как декоративные в комнатах и оранжереях. Из них наиболее распространен хлорофитум хохолковый, имеющий ряд культурных разновидностей, в том числе пестролистную форму с продольными белыми полосами, а также хлорофитум капский.
Виды хлорофитума —- многолетние травы с ползучим корневищем, обычно тонкими, но у концов часто клубневидно утолщенными корнями, с розетками листьев и с центральным простым или разветвленным цветоносом. Самый высокий цветонос (до 2 м), а также наиболее длинные (до 140 см) слабосуккулентные листья имеет похожий на алоэ хлорофитум Крука (С. krookianum), встречающийся по сырым местам в восточных районах Южной Африки (рис. 72). Листья у хлорофитумов от линейных до широколанцетных, по краю обычно бахромчатые, а при основании нередко суженные в черешок. Мелкие, белые, реже зеленоватые или розоватые цветки собраны в 1 —- 6-цветковые пучки, расположенные в пазухе прицветников и образующие рыхлые или более или менее густые сложные кисти. Каждый цветок такого пучка сидит непосредственно на оси соцветия в пазухе прицветника. Пучки представляют собой редуцированные боковые ветви соцветия, оси которых подверглись полной редукции. У некоторых видов (например, у хлорофитума капского —- С. capense) иногда развиваются очень короткие оси и цветки на них расположены по спирали. Околоцветник у хлорофитумов со свободными сегментами, обычно расположенными звездчато или отклоненными вниз. Тычинки с крупными пыльниками, прикрепляющимися к нити основанием, при увядании изгибаются назад. Гинецей состоит из 3 плодолистиков, но у хлорофитума клубневого (С. tuberosum), распространенного в тропической Африке и Индии, он образован только 2 плодолистиками. В каждом гнезде завязи имеется от 6 до 30 семязачатков. Коробочка 3-гранная, часто глубоколопастная. Семена крупные, диаметром 2 —- 4 мм, плоские, округлые, черные и блестящие.Цветет хлорофитум в течение всего года, но разные его виды —- в различное время. Цветки в пучках раскрываются последовательно, и первым распускается нижний бутон. При неблагоприятных для цветения условиях рост бутонов может задержаться на продолжительное время, и если они вновь начнут расти и распускаться, то в одних и тех же пучках можно видеть и зрелые плоды, и раскрытые и увядшие цветки, и бутоны. Цветоножки при цветении направлены косо вверх и не изменяют своего положения при плодах. Цветки опыляются пчелами, цветочными мухами и другими насекомыми, привлекаемыми пыльцой и нектаром. Несколько видов хлорофитума разводят как декоративные в комнатах и оранжереях. Из них наиболее распространен хлорофитум хохолковый, имеющий ряд культурных разновидностей, в том числе пестролистную форму с продольными белыми полосами, а также хлорофитум капский.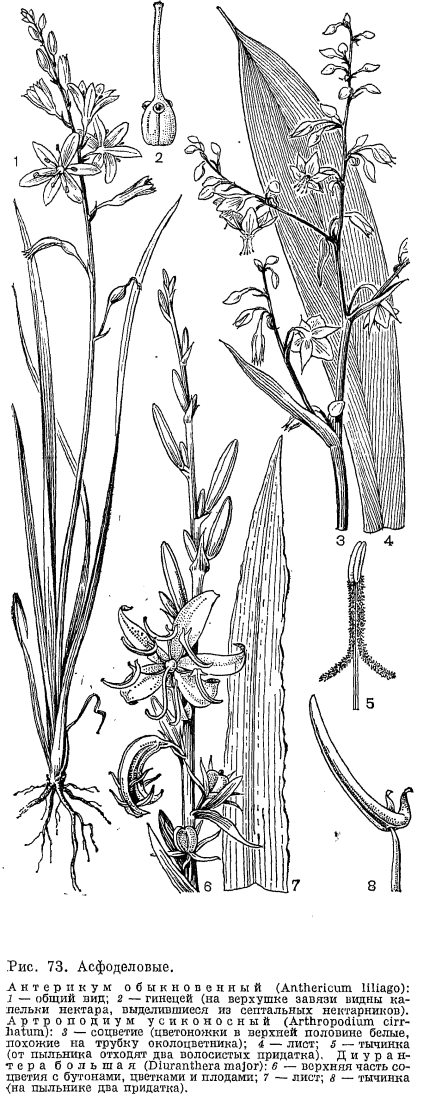 К роду хлорофитум очень близок род антерикум (Anthericum.), отличающийся от него главным образом мелкими, шаровидными в очертании, угловатыми, черными семенами. В род антерикум входит около 250 видов, распространенных преимущественно в тропической Африке. Из них 17 видов произрастают в Южной Африке, 2 —- в Северной Африке, 3 —- в Европе, несколько —- на юге Северной Америки, в Центральной и Южной Америке. Два европейских вида —- антерикум ветвистый (А. rаmosum) и антерикум обыкновенный (A. liliago, рис. 73) —- достигают в своем распространении юга Швеции и, следовательно, дальше других представителей семейства асфоделовых проникают в северные широты. Антерикум ветвистый встречается и в южных, и отчасти центральных районах европейской части СССР. Оба вида обитают по сухим солнечным местам, травянистым и каменистым склонам гор и холмов, сухим разреженным лесам и кустарникам. Европейские виды имеют простые кисти, в то время как у большинства остальных видов они сложные, образованные не одиночными цветками, а пучками цветков. Африканские виды антерикума обитают в условиях, в основном сходных с таковыми у видов хлорофитума. Многие тропические виды растут в горах, по открытым травянистым и лесным склонам, скалам, берегам горных рек. В Южной Африке представители этого рода чаще всего встречаются в горных и равнинных кустарниковых песчаных саваннах. У антерикума обыкновенного пыльники вскрываются одновременно с раскрыванием цветков или сразу после этого. Самоопыление устраняется благодаря разной длине тычинок и столбика, который превышает тычинки примерно на 5 мм и, кроме того, немного отогнут вниз. На верхней поверхности завязи из септальных нектарников крупными каплями выделяется нектар (рис. 73, 2). Аитерикум разводят как декоративное растение в открытом грунте.Следует упомянуть об эндемичном для Африки роде трахиандра (Trachyandra), насчитывающем 50 видов, из которых 45 встречаются в Южной Африке, преимущественно в Капской области, а остальные —- в тропической Африке. Среди видов этого рода имеется несколько кустарниковидных форм, очень редких в подсемействе антериковых. В качестве примера назовем встречающуюся на юго-западе Капской области трахиандру Адамсона (Т. adamsonii) —- кустарник высотой до 180 см, с ветвями, покрытыми твердыми основаниями опавших листьев и несущими на верхушках пучки узких, сизых, слабосуккулентных листьев. Оригинально выглядят некоторые травянистые представители рода, например трахиандра рыхлая (Т. 1аха), обитающая в пустыне Калахари и других засушливых районах Южной Африки. После образования плодов веточки его сильно ветвистого раскидистого соцветия, а также цветоножки дуговидно загибаются книзу, и тогда все соцветие приобретает шаровидную форму. Отрываясь от растения, эти сухие шаровидные соцветия (перекати-поле) перекатываются ветром по земле, разбрасывая семена.Три монотипных рода из трибы антериковых —- анемаррена, тераухия (Terauchia) и алекторурус —- представляют семейство асфоделовых в Восточной Азии. Анемаррена асфоделовидная (Anemarrhena asphodeloides) растет по сухим склонам гор в лесостепных и степных районах Северо-Восточного и Северного Китая и на юге полуострова Корея, образуя иногда своеобразные анемарреновые степи. Она имеет горизонтальное ветвящееся корневище, на конце которого развивается густой пучок злаковидных желобчатых листьев и цветонос длиной до 1 м с негустой кистью мелких невзрачных цветков со слабым сладковатым запахом. Сегменты околоцветника, сросшиеся до половины своей длины, образуют тускло-пурпуровую трубку и желтый или фиолетовый отгиб. Цветки имеют только 3 тычинки, почти полностью сросшиеся нитями с внутренними сегментами околоцветника (рис. 72, 8). Тычинки скрыты в трубке околоцветника и расположены выше столбика. Анемаррена цветет в середине лета. Цветки ее раскрываются после 7 ч вечера и опыляются мелкими ночными бабочками из группы совок (Acronicta psi) и огневок (Pyrausta gracilis).К анемаррене очень близка тераухия анемарренолистная (Terauchia anemarrhenaefolia), встречающаяся на севере полуострова Корея. У этого вида известны только мужские цветки с недоразвитой завязью. Единственный вид алекторуруса —- алекторурус йедоенский (А1еctorurus yedoensis) изредка встречается в Японии, на островах Сикоку и Кюсю, в горах на влажных скалах. Это небольшое двудомное растение с розеткой двурядных вечнозеленых широколинейных листьев и цветоносом высотой 15 —- 40 см, с рыхлым метельчатым соцветием из мелких белых колокольчатых цветков. Семена его снабжены при основании длинными белыми волосками (рис. 72, 7). Эта особенность свойственна только данному роду. Алекторурус занимает в трибе антериковых обособленное положение.
К роду хлорофитум очень близок род антерикум (Anthericum.), отличающийся от него главным образом мелкими, шаровидными в очертании, угловатыми, черными семенами. В род антерикум входит около 250 видов, распространенных преимущественно в тропической Африке. Из них 17 видов произрастают в Южной Африке, 2 —- в Северной Африке, 3 —- в Европе, несколько —- на юге Северной Америки, в Центральной и Южной Америке. Два европейских вида —- антерикум ветвистый (А. rаmosum) и антерикум обыкновенный (A. liliago, рис. 73) —- достигают в своем распространении юга Швеции и, следовательно, дальше других представителей семейства асфоделовых проникают в северные широты. Антерикум ветвистый встречается и в южных, и отчасти центральных районах европейской части СССР. Оба вида обитают по сухим солнечным местам, травянистым и каменистым склонам гор и холмов, сухим разреженным лесам и кустарникам. Европейские виды имеют простые кисти, в то время как у большинства остальных видов они сложные, образованные не одиночными цветками, а пучками цветков. Африканские виды антерикума обитают в условиях, в основном сходных с таковыми у видов хлорофитума. Многие тропические виды растут в горах, по открытым травянистым и лесным склонам, скалам, берегам горных рек. В Южной Африке представители этого рода чаще всего встречаются в горных и равнинных кустарниковых песчаных саваннах. У антерикума обыкновенного пыльники вскрываются одновременно с раскрыванием цветков или сразу после этого. Самоопыление устраняется благодаря разной длине тычинок и столбика, который превышает тычинки примерно на 5 мм и, кроме того, немного отогнут вниз. На верхней поверхности завязи из септальных нектарников крупными каплями выделяется нектар (рис. 73, 2). Аитерикум разводят как декоративное растение в открытом грунте.Следует упомянуть об эндемичном для Африки роде трахиандра (Trachyandra), насчитывающем 50 видов, из которых 45 встречаются в Южной Африке, преимущественно в Капской области, а остальные —- в тропической Африке. Среди видов этого рода имеется несколько кустарниковидных форм, очень редких в подсемействе антериковых. В качестве примера назовем встречающуюся на юго-западе Капской области трахиандру Адамсона (Т. adamsonii) —- кустарник высотой до 180 см, с ветвями, покрытыми твердыми основаниями опавших листьев и несущими на верхушках пучки узких, сизых, слабосуккулентных листьев. Оригинально выглядят некоторые травянистые представители рода, например трахиандра рыхлая (Т. 1аха), обитающая в пустыне Калахари и других засушливых районах Южной Африки. После образования плодов веточки его сильно ветвистого раскидистого соцветия, а также цветоножки дуговидно загибаются книзу, и тогда все соцветие приобретает шаровидную форму. Отрываясь от растения, эти сухие шаровидные соцветия (перекати-поле) перекатываются ветром по земле, разбрасывая семена.Три монотипных рода из трибы антериковых —- анемаррена, тераухия (Terauchia) и алекторурус —- представляют семейство асфоделовых в Восточной Азии. Анемаррена асфоделовидная (Anemarrhena asphodeloides) растет по сухим склонам гор в лесостепных и степных районах Северо-Восточного и Северного Китая и на юге полуострова Корея, образуя иногда своеобразные анемарреновые степи. Она имеет горизонтальное ветвящееся корневище, на конце которого развивается густой пучок злаковидных желобчатых листьев и цветонос длиной до 1 м с негустой кистью мелких невзрачных цветков со слабым сладковатым запахом. Сегменты околоцветника, сросшиеся до половины своей длины, образуют тускло-пурпуровую трубку и желтый или фиолетовый отгиб. Цветки имеют только 3 тычинки, почти полностью сросшиеся нитями с внутренними сегментами околоцветника (рис. 72, 8). Тычинки скрыты в трубке околоцветника и расположены выше столбика. Анемаррена цветет в середине лета. Цветки ее раскрываются после 7 ч вечера и опыляются мелкими ночными бабочками из группы совок (Acronicta psi) и огневок (Pyrausta gracilis).К анемаррене очень близка тераухия анемарренолистная (Terauchia anemarrhenaefolia), встречающаяся на севере полуострова Корея. У этого вида известны только мужские цветки с недоразвитой завязью. Единственный вид алекторуруса —- алекторурус йедоенский (А1еctorurus yedoensis) изредка встречается в Японии, на островах Сикоку и Кюсю, в горах на влажных скалах. Это небольшое двудомное растение с розеткой двурядных вечнозеленых широколинейных листьев и цветоносом высотой 15 —- 40 см, с рыхлым метельчатым соцветием из мелких белых колокольчатых цветков. Семена его снабжены при основании длинными белыми волосками (рис. 72, 7). Эта особенность свойственна только данному роду. Алекторурус занимает в трибе антериковых обособленное положение.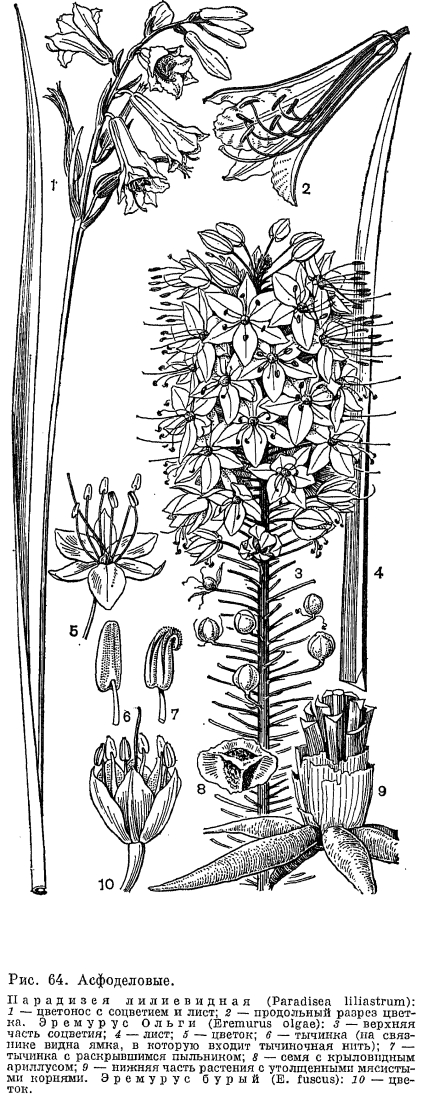 В горах Средней и Южной Европы распространен род парадизея (Paradisea), названный по имени итальянского ботаника конца XVIII и начала XIX в. Д. Парадизи. В род входят 2 вида, из которых более известна парадизея лилиевидная (P. liliastrum), растущая на альпийских и субальпийских лугах в Пиренеях, Юре, Альпах и Апеннинах (рис. 64, 1). Этот вид имеет самые крупные в подсемействе цветки (длиной 3 —- 5 см). Они белые, воронковидные, со свободными сегментами, собранные в 2 —- 20-цветковую более или менее одностороннюю кисть. Их 3 длинные и 3 короткие тычинки с качающимися пыльниками дуговидно изогнуты наверх так же, как и немного превышающий их столбик. Парадизею лилиевидную разводят как декоративное растение, в особенности на альпийских горках.В Новом Свете обитают 3 эндемичных рода из трибы антериковых. Из них 2 —- эремокринум (Eremocrinum), распространенный на юге США (штаты Аризона и Юта), и чилийский род боттионея (Bottionea) —- являются монотипными. Третий, исключительно тропический род эхеандия (Echeandia), объединяет 13 видов, распространенных в Центральной Америке (от Мексики до Панамы) и на севере Южной Америки (Венесуэла и Эквадор). Виды эхеандии —- многолетние травы с утолщенными от основания корнями, длинными линейными листьями и собранными в пучки цветками, образующими на верхушке цветоноса и его боковых ветвей рыхлые кистевидные соцветия. Пыльники у эхеандии срослись в довольно длинную коническую трубку, окружающую столбик, что является приспособлением к перекрестному опылению. Срастание пыльников —- характерная особенность этого рода. Австралийским ботаником П. Бернхардтом и американским Э. Монталво (1977, 1979) детально изучена биология цветения эхеандии крупноплодной (Е. macrocarpa), произрастающей в горах Центральной Америки в дубово-можжевеловых лесах на высоте до 1800 м над уровнем моря. Как отмечают названные авторы, эхеандии в строении андроцея обнаруживают замечательное сходство с двудольными растениями умеренного пояса —- додекатеоном обыкновенным (Dodecatheon meaclia) и пасленом сладко-горьким (Solanum dulcamara), у которых пыльники также образуют конус вокруг столбика, хотя и не срастаются при этом между собой. Указанное сходство обусловливает и одинаковый механизм опыления у этих растений. Цветки эхеандии крупноплодной желтые, с колесовидным околоцветником и во время цветения обращены косо вниз (рис. 72, 6).Они строго протогиничны. В женскую фазу, которая продолжается один день, из пыльникового конуса высовывается столбик. И только после того, как рыльце потеряет способность воспринимать пыльцу и столбик втянется в пыльниковый конус, происходит вскрывание пыльников. Поэтому самоопыление у эхеандии совершенно исключено. Мужская фаза продолжается от 2 до 7 суток в зависимости от погодных условий. Следовательно, каждый отдельный цветок большую часть жизни функционирует как донор пыльцы, а не как ее получатель. Цветки эхеандии не имеют нектара, и насекомые посещают их ради пыльцы. Главными опылителями, в большом количестве встречающиеся на цветках эхеандии, являются шмели —- рабочие особи и царицы. Подлетая к цветку, рабочий шмель обхватывает пыльниковый конус передними парами ног и мандибулами. При вибрации грудных крыльев пыльниковый конус сотрясается и пыльца высыпается из его внутренних продольных щелей на голову, грудь и брюшко насекомого, которое стряхивает ее затем второй парой ног в пыльцевые мешки, находящиеся на задних ногах. Шмели-царицы ведут себя аналогично рабочим особям с той лишь разницей, что они иногда прекращают вибрацию и слизывают выпавшую из конуса пыльцу длинным язычком. Каждую минуту насекомые посещают в среднем 14 цветков, причем 24 с насекомое тратит на полет, а 36 с —- на получение пыльцы путем вибрации. Несмотря на устойчивые взаимоотношения с насекомыми-опылителями, в соцветиях эхеандии крупноплодной образуется мало плодов (в среднем 8 плодов на соцветие, в котором до 66 бутонов). Такая малая продуктивность может быть объяснена двумя причинами. Во-первых, только 15 —- 20% цветков в популяции находятся одновременно в женской фазе. Во-вторых, из 4 —- 5 цветков пазушного пучка только один (нижний) способен после опыления образовать плод, остальные же цветки пучка (их называют вспомогательными) являются функционально пыльниковьми. Они хотя и имеют внешне нормально развитый гинецей, но оказываются не в состоянии завязать плоды. Наличие вспомогательных цветков и большую продолжительность мужской фазы цветка по сравнению с женской Бернхардт и Монталво считают приспособлением, выработанным растениями, чтобы обеспечить насекомых обильной пищей —- пыльцой —- и тем самым постоянно привлекать их к себе и опылять цветки, от которых они ничего не получают (в женской фазе пыльники закрыты). Описанная «стратегия» опыления, как полагают, является выгодной для растения, так как, с одной стороны, она надежно обеспечивает перекрестное опыление, а с другой —- благодаря образованию небольшого количества плодов позволяет растению меньше истощать энергетические резервы и полнее сохранить их для следующей вегетации. Эхеандия крупноплодная цветет с начала июля, в сезон дождей, при температуре 14 —- 15 °С. Вскрывание плодов у нее приурочено к периоду сильных ветров (конец октября —- начало ноября), что способствует более успешному разбрасыванию семян из обращенных вверх коробочек этого растения-баллиста.
В горах Средней и Южной Европы распространен род парадизея (Paradisea), названный по имени итальянского ботаника конца XVIII и начала XIX в. Д. Парадизи. В род входят 2 вида, из которых более известна парадизея лилиевидная (P. liliastrum), растущая на альпийских и субальпийских лугах в Пиренеях, Юре, Альпах и Апеннинах (рис. 64, 1). Этот вид имеет самые крупные в подсемействе цветки (длиной 3 —- 5 см). Они белые, воронковидные, со свободными сегментами, собранные в 2 —- 20-цветковую более или менее одностороннюю кисть. Их 3 длинные и 3 короткие тычинки с качающимися пыльниками дуговидно изогнуты наверх так же, как и немного превышающий их столбик. Парадизею лилиевидную разводят как декоративное растение, в особенности на альпийских горках.В Новом Свете обитают 3 эндемичных рода из трибы антериковых. Из них 2 —- эремокринум (Eremocrinum), распространенный на юге США (штаты Аризона и Юта), и чилийский род боттионея (Bottionea) —- являются монотипными. Третий, исключительно тропический род эхеандия (Echeandia), объединяет 13 видов, распространенных в Центральной Америке (от Мексики до Панамы) и на севере Южной Америки (Венесуэла и Эквадор). Виды эхеандии —- многолетние травы с утолщенными от основания корнями, длинными линейными листьями и собранными в пучки цветками, образующими на верхушке цветоноса и его боковых ветвей рыхлые кистевидные соцветия. Пыльники у эхеандии срослись в довольно длинную коническую трубку, окружающую столбик, что является приспособлением к перекрестному опылению. Срастание пыльников —- характерная особенность этого рода. Австралийским ботаником П. Бернхардтом и американским Э. Монталво (1977, 1979) детально изучена биология цветения эхеандии крупноплодной (Е. macrocarpa), произрастающей в горах Центральной Америки в дубово-можжевеловых лесах на высоте до 1800 м над уровнем моря. Как отмечают названные авторы, эхеандии в строении андроцея обнаруживают замечательное сходство с двудольными растениями умеренного пояса —- додекатеоном обыкновенным (Dodecatheon meaclia) и пасленом сладко-горьким (Solanum dulcamara), у которых пыльники также образуют конус вокруг столбика, хотя и не срастаются при этом между собой. Указанное сходство обусловливает и одинаковый механизм опыления у этих растений. Цветки эхеандии крупноплодной желтые, с колесовидным околоцветником и во время цветения обращены косо вниз (рис. 72, 6).Они строго протогиничны. В женскую фазу, которая продолжается один день, из пыльникового конуса высовывается столбик. И только после того, как рыльце потеряет способность воспринимать пыльцу и столбик втянется в пыльниковый конус, происходит вскрывание пыльников. Поэтому самоопыление у эхеандии совершенно исключено. Мужская фаза продолжается от 2 до 7 суток в зависимости от погодных условий. Следовательно, каждый отдельный цветок большую часть жизни функционирует как донор пыльцы, а не как ее получатель. Цветки эхеандии не имеют нектара, и насекомые посещают их ради пыльцы. Главными опылителями, в большом количестве встречающиеся на цветках эхеандии, являются шмели —- рабочие особи и царицы. Подлетая к цветку, рабочий шмель обхватывает пыльниковый конус передними парами ног и мандибулами. При вибрации грудных крыльев пыльниковый конус сотрясается и пыльца высыпается из его внутренних продольных щелей на голову, грудь и брюшко насекомого, которое стряхивает ее затем второй парой ног в пыльцевые мешки, находящиеся на задних ногах. Шмели-царицы ведут себя аналогично рабочим особям с той лишь разницей, что они иногда прекращают вибрацию и слизывают выпавшую из конуса пыльцу длинным язычком. Каждую минуту насекомые посещают в среднем 14 цветков, причем 24 с насекомое тратит на полет, а 36 с —- на получение пыльцы путем вибрации. Несмотря на устойчивые взаимоотношения с насекомыми-опылителями, в соцветиях эхеандии крупноплодной образуется мало плодов (в среднем 8 плодов на соцветие, в котором до 66 бутонов). Такая малая продуктивность может быть объяснена двумя причинами. Во-первых, только 15 —- 20% цветков в популяции находятся одновременно в женской фазе. Во-вторых, из 4 —- 5 цветков пазушного пучка только один (нижний) способен после опыления образовать плод, остальные же цветки пучка (их называют вспомогательными) являются функционально пыльниковьми. Они хотя и имеют внешне нормально развитый гинецей, но оказываются не в состоянии завязать плоды. Наличие вспомогательных цветков и большую продолжительность мужской фазы цветка по сравнению с женской Бернхардт и Монталво считают приспособлением, выработанным растениями, чтобы обеспечить насекомых обильной пищей —- пыльцой —- и тем самым постоянно привлекать их к себе и опылять цветки, от которых они ничего не получают (в женской фазе пыльники закрыты). Описанная «стратегия» опыления, как полагают, является выгодной для растения, так как, с одной стороны, она надежно обеспечивает перекрестное опыление, а с другой —- благодаря образованию небольшого количества плодов позволяет растению меньше истощать энергетические резервы и полнее сохранить их для следующей вегетации. Эхеандия крупноплодная цветет с начала июля, в сезон дождей, при температуре 14 —- 15 °С. Вскрывание плодов у нее приурочено к периоду сильных ветров (конец октября —- начало ноября), что способствует более успешному разбрасыванию семян из обращенных вверх коробочек этого растения-баллиста.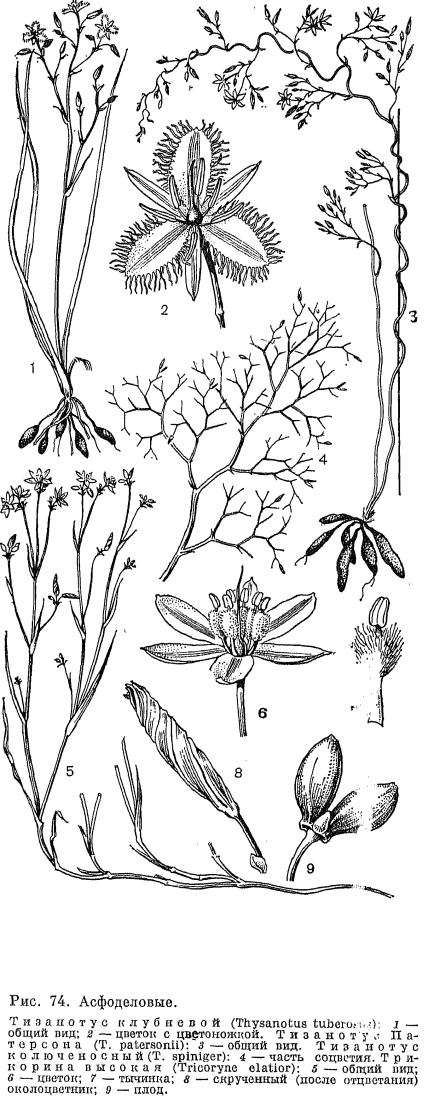 Триба тизанотовых (Thysanoteae) состоит из одного рода тизанотус. Он включает 37 видов, распространенных в Австралии, из них 2 вида встречаются в Новой Гвинее, а 1 —- тизанотус китайский (Thysanotus chinensis) —- в Южном Китае, Таиланде, Вьетнаме, на Филиппинах и в Индонезии. Большинство представителей рода растет в умеренном и субтропическом поясах, часто встречаясь на песчаных равнинах, покрытых верещатниками, на приморских песках, по сухим холмам. Большую экологическую амплитуду имеет распространенный почти по всему ареалу рода тизанотус клубневой (Т. tuberosus), встречающийся от пустынных районов Центральной Австралии, где растет в сообществах ксерофитных злаков, до тропических районов Квинсленда (рис. 74, 1). Все тизанотусы —- многолетние травы с коротким корневищем, от которого отходит пучок многочисленных тонких или (чаще) клубневидно утолщенных корней. Листья прикорневые, линейные, злаковидные, иногда полуцилиндрические или несколько мясистые. У немногих представителей (тизанотуса китайского и тизанотуса многоцветкового —- Т. multiflorus) они вечнозеленые; у некоторых других видов (например, у тизанотуса колюченосного —- Т. spiniger) листья отмирают еще до начала цветения и ассимиляционная функция переходит к цветоносу. У тизанотуса колючего большинство веточек соцветия заканчивается колючим острием, и только немногие из них 1-, 2-цветковым частным соцветием (рис. 74, 4). У единственной в роде лианы —- тизанотуса Патерсона (Т. patersonii) —- цветонос извилистый, вьющийся, длиной до 1 м, с короткими несущими пучки цветков боковыми ветвями (рис. 74, 3). Своеобразное строение имеют цветки тизанотусов: внутренние сегменты распростертого околоцветника резко отличаются от наружных —- они значительно шире благодаря перепончатым, часто фиолетовым и обычно бахромчато-реснитчатым краям (рис. 74, 2). Тычинки, как правило, неравные: 3 внутренние имеют более длинные нити и особенно пыльники; у некоторых видов внутренние тычинки совсем не развиваются. Пыльники линейные, длиной до 1 см, прикрепляются к нити основанием, реже спинкой и вскрываются верхушечной порой. При увядании они обычно спирально скручиваются. В каждом гнезде завязи 2 семязачатка, но иногда их 10 —- 17. При основании семян имеется маленький придаток —- элайосома. Род тизанотус благодаря особенностям строения цветков, а также наличию в хлоренхиме листьев и стеблей рафидных каналов с оксалатом кальция, которых нет у других представителей порядка лилейных, занимает в подсемействе антериковых довольно обособленное положение.В трибе ходжсониоловых (Hodgsonioleae) различают 7 родов, их представители обитают в южном полушарии. Монотипный род паситея (Pasithea) встречается в Южной Америке (юг Перу, Чили), 5 моно- и олиготииных родов —- в Австралии, Тасмании и на востоке Новой Гвинеи и 1 род цесия (Caesia, 12 видов) —- на юге Африки, Мадагаскаре и в Австралии. Все представители этой трибы. —- многолетние травы с прикорневыми, иногда двурядными (виды цесии) листьями и безлистными цветоносами или редко (роды трикорина и коринотека) с облиственными ветвистыми стеблями. Листья обычно линейные, злаковидные, иногда шиловидные, при основании нередко с влагалищами. Цветки мелкие, обычно голубые, редко белые или желтые, собранные в метелку, простую или сложную кисть или в малоцветковые зонтиковидные верхушечные соцветия (например, у трикорины)- Околоцветник звездчатый, со свободными, редко сросшимися внизу в короткую трубку сегментами. При отцветании околоцветник спирально скручивается (рис. 74, 8), что является характерным признаком этой трибы, который свойствен еще только одному австралийскому роду —- арнокринуму (Arnocrinum) из трибы джонсониевых. Скрученный околоцветник более или менее долго сохраняется при развивающемся плоде, но затем опадает. Тычинок обычно 6, пыльники прикрепляются спинкой и вскрываются интрорзно, а у рода агростокринум (Agrostocrinum) —- верхушечной порой. У рода ходжсониола (Hodgsoniola) имеются 3 фертильные и 3 стерильные тычинки. У первых связники удлинены и срослись в цилиндрическую трубку, из которой выступает столбик. У рода трикорина нити тычинок очень густо опушены обращенными вверх волосками (рис. 74, 7). Число семязачатков в каждом гнезде завязи варьирует от многочисленных (у родов паситея и хамесцилла —- Chamaescil1а) до 2 у остальных ее представителей. Роды трикорина и коринотека имеют необычные для семейства асфоделовых плоды. У трикорины в каждом гнезде ее глубоко трехлопастной завязи из 2 семязачатков развивается 1 и, кроме того, 1 или 2 гнезда завязи часто дегенерируют.Образующийся плод состоит из 3 —- 1 односемянных невскрывающихся сухих или сочных долей (рис. 74, 9). Односемянным невскрывающимся часто бывает плод у растений из рода коринотека, когда в их завязи развивается только одно гнездо с 1 семязачатком. Но если развивается больше гнезд и семян, то тогда образуется довольно поздно вскрывающийся плод-коробочка. Таким образом на примере родов трикорина и коринотека можно наглядно видеть возможные пути трансформации коробочки в односемянный невскрывающийся плод. Семена цесии и коринотеки снабжены при основании элайосомами; у цесии элайосомы являются строфиолями, у коринотеки —- карункулами.
Триба тизанотовых (Thysanoteae) состоит из одного рода тизанотус. Он включает 37 видов, распространенных в Австралии, из них 2 вида встречаются в Новой Гвинее, а 1 —- тизанотус китайский (Thysanotus chinensis) —- в Южном Китае, Таиланде, Вьетнаме, на Филиппинах и в Индонезии. Большинство представителей рода растет в умеренном и субтропическом поясах, часто встречаясь на песчаных равнинах, покрытых верещатниками, на приморских песках, по сухим холмам. Большую экологическую амплитуду имеет распространенный почти по всему ареалу рода тизанотус клубневой (Т. tuberosus), встречающийся от пустынных районов Центральной Австралии, где растет в сообществах ксерофитных злаков, до тропических районов Квинсленда (рис. 74, 1). Все тизанотусы —- многолетние травы с коротким корневищем, от которого отходит пучок многочисленных тонких или (чаще) клубневидно утолщенных корней. Листья прикорневые, линейные, злаковидные, иногда полуцилиндрические или несколько мясистые. У немногих представителей (тизанотуса китайского и тизанотуса многоцветкового —- Т. multiflorus) они вечнозеленые; у некоторых других видов (например, у тизанотуса колюченосного —- Т. spiniger) листья отмирают еще до начала цветения и ассимиляционная функция переходит к цветоносу. У тизанотуса колючего большинство веточек соцветия заканчивается колючим острием, и только немногие из них 1-, 2-цветковым частным соцветием (рис. 74, 4). У единственной в роде лианы —- тизанотуса Патерсона (Т. patersonii) —- цветонос извилистый, вьющийся, длиной до 1 м, с короткими несущими пучки цветков боковыми ветвями (рис. 74, 3). Своеобразное строение имеют цветки тизанотусов: внутренние сегменты распростертого околоцветника резко отличаются от наружных —- они значительно шире благодаря перепончатым, часто фиолетовым и обычно бахромчато-реснитчатым краям (рис. 74, 2). Тычинки, как правило, неравные: 3 внутренние имеют более длинные нити и особенно пыльники; у некоторых видов внутренние тычинки совсем не развиваются. Пыльники линейные, длиной до 1 см, прикрепляются к нити основанием, реже спинкой и вскрываются верхушечной порой. При увядании они обычно спирально скручиваются. В каждом гнезде завязи 2 семязачатка, но иногда их 10 —- 17. При основании семян имеется маленький придаток —- элайосома. Род тизанотус благодаря особенностям строения цветков, а также наличию в хлоренхиме листьев и стеблей рафидных каналов с оксалатом кальция, которых нет у других представителей порядка лилейных, занимает в подсемействе антериковых довольно обособленное положение.В трибе ходжсониоловых (Hodgsonioleae) различают 7 родов, их представители обитают в южном полушарии. Монотипный род паситея (Pasithea) встречается в Южной Америке (юг Перу, Чили), 5 моно- и олиготииных родов —- в Австралии, Тасмании и на востоке Новой Гвинеи и 1 род цесия (Caesia, 12 видов) —- на юге Африки, Мадагаскаре и в Австралии. Все представители этой трибы. —- многолетние травы с прикорневыми, иногда двурядными (виды цесии) листьями и безлистными цветоносами или редко (роды трикорина и коринотека) с облиственными ветвистыми стеблями. Листья обычно линейные, злаковидные, иногда шиловидные, при основании нередко с влагалищами. Цветки мелкие, обычно голубые, редко белые или желтые, собранные в метелку, простую или сложную кисть или в малоцветковые зонтиковидные верхушечные соцветия (например, у трикорины)- Околоцветник звездчатый, со свободными, редко сросшимися внизу в короткую трубку сегментами. При отцветании околоцветник спирально скручивается (рис. 74, 8), что является характерным признаком этой трибы, который свойствен еще только одному австралийскому роду —- арнокринуму (Arnocrinum) из трибы джонсониевых. Скрученный околоцветник более или менее долго сохраняется при развивающемся плоде, но затем опадает. Тычинок обычно 6, пыльники прикрепляются спинкой и вскрываются интрорзно, а у рода агростокринум (Agrostocrinum) —- верхушечной порой. У рода ходжсониола (Hodgsoniola) имеются 3 фертильные и 3 стерильные тычинки. У первых связники удлинены и срослись в цилиндрическую трубку, из которой выступает столбик. У рода трикорина нити тычинок очень густо опушены обращенными вверх волосками (рис. 74, 7). Число семязачатков в каждом гнезде завязи варьирует от многочисленных (у родов паситея и хамесцилла —- Chamaescil1а) до 2 у остальных ее представителей. Роды трикорина и коринотека имеют необычные для семейства асфоделовых плоды. У трикорины в каждом гнезде ее глубоко трехлопастной завязи из 2 семязачатков развивается 1 и, кроме того, 1 или 2 гнезда завязи часто дегенерируют.Образующийся плод состоит из 3 —- 1 односемянных невскрывающихся сухих или сочных долей (рис. 74, 9). Односемянным невскрывающимся часто бывает плод у растений из рода коринотека, когда в их завязи развивается только одно гнездо с 1 семязачатком. Но если развивается больше гнезд и семян, то тогда образуется довольно поздно вскрывающийся плод-коробочка. Таким образом на примере родов трикорина и коринотека можно наглядно видеть возможные пути трансформации коробочки в односемянный невскрывающийся плод. Семена цесии и коринотеки снабжены при основании элайосомами; у цесии элайосомы являются строфиолями, у коринотеки —- карункулами.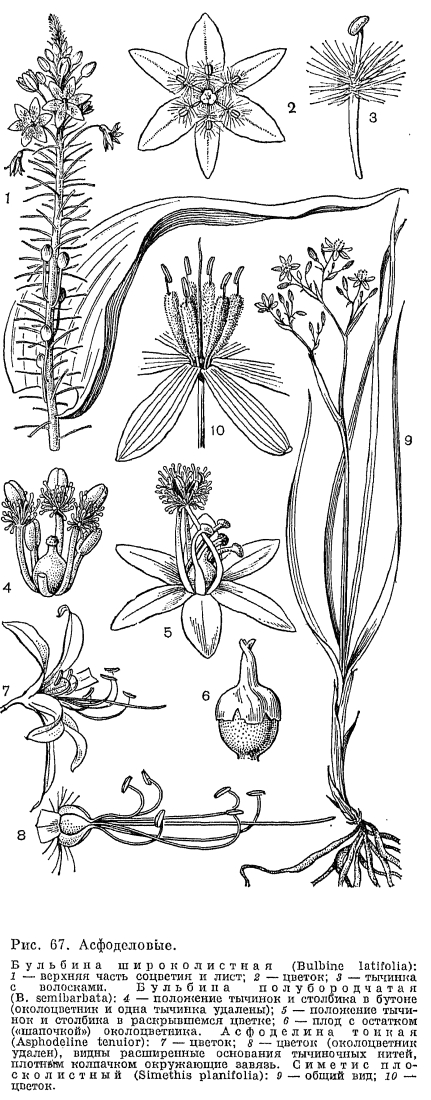 С представителями рассмотренной трибы обнаруживает родство по эмбриологическим признакам род симетис '(Simethis), выделенный X. Хубером (1969) в трибу симетисовых (Simethideae). Единственный вид рода —- симетис плосколистный (S. planifolia) произрастает в Западном Средиземноморье (Алжир, Марокко Тунис, юг Европы до Италии), на западе Франции и юго-западе Ирландии, встречаясь по верещатникам, каменистым местам, разреженным сосновым лесам. Это многолетнее растение с мясистыми шнуровидными корнями, пучком длинных прикорневых влагалищных листьев и цветоносом длиной до 40 см (рис. 67, 9, 10). Соцветие у симетиса рыхлое, метельчатое, с немногими ветвями, несущими пучки из 3 —- 7 цветков. Околоцветник звездчатый, со свободными сегментами, сверху белыми, снизу пурпурными. Тычинки с длинными пыльниками и густо шерстисто-волосистыми нитями, прикрепляющиеся к ямке связника. В каждом гнезде завязи образуются по 2 семязачатка. Плод —- шаровидная коробочка. Семена при основании с белым придатком —- строфиолой.
С представителями рассмотренной трибы обнаруживает родство по эмбриологическим признакам род симетис '(Simethis), выделенный X. Хубером (1969) в трибу симетисовых (Simethideae). Единственный вид рода —- симетис плосколистный (S. planifolia) произрастает в Западном Средиземноморье (Алжир, Марокко Тунис, юг Европы до Италии), на западе Франции и юго-западе Ирландии, встречаясь по верещатникам, каменистым местам, разреженным сосновым лесам. Это многолетнее растение с мясистыми шнуровидными корнями, пучком длинных прикорневых влагалищных листьев и цветоносом длиной до 40 см (рис. 67, 9, 10). Соцветие у симетиса рыхлое, метельчатое, с немногими ветвями, несущими пучки из 3 —- 7 цветков. Околоцветник звездчатый, со свободными сегментами, сверху белыми, снизу пурпурными. Тычинки с длинными пыльниками и густо шерстисто-волосистыми нитями, прикрепляющиеся к ямке связника. В каждом гнезде завязи образуются по 2 семязачатка. Плод —- шаровидная коробочка. Семена при основании с белым придатком —- строфиолой. Очень своеобразна эндемичная для Австралии триба джонсониевых, иногда выделяемая в отдельное семейство. В нее входит 8 родов, включающих многолетние травы, реже кустарничковидные формы. Многие представители трибы являются ксерофильными растениями. Характерным признаком джонсониевых является наличие головчатого, реже зонтиковидного соцветия, окруженного оберткой кроющих листьев; иногда (у рода джонсония —- Johnsonia) соцветие —- простой колос. Другая особенность джонсониевых —- унифациальные (билатеральные или 3-гранные) листовые пластинки. Листья джонсониевых расположены в прикорневых или верхушечных пучках или по всему стеблю. Цветки сидят в пазухе прицветников, причем у некоторых родов каждый цветок имеет еще и прицветничек, расположенный на его вентральной (брюшной) стороне и в разной степени охватывающий цветок (рис. 75). Сегменты околоцветника свободные или сросшиеся в основании или наполовину своей длины, образуя в последнем случае трубку и отгиб (например, у рода бория —- Воrуа). В цветке 6 или 3 тычинки; в каждом гнезде завязи от многих до нескольких семязачатков.В роде джоисония 3 вида, обитающих в Юго-Западной Австралии по сухим приморским пескам. Наиболее известна джонсония обыкновенная (J. lupulina) —- плотнодерновинный многолетник с жесткими двурядно расположенными билатеральными узколинейными листьями и уплощенным стеблем (рис. 75, 1). Джонсонию можно принять за растение из семейства злаков. Злаковидный облик имеют не только дерновины этого вида, но и его соцветие —- густой продолговато-эллиптический колос с крупными скрывающими цветки, тонкокожистыми, как у злаков, чешуями. При основании колоса имеется кроющий лист, направленный вертикально вверх, благодаря чему создается впечатление, что он является продолжением стебля, а соцветие занимает не верхушечное, а боковое положение. Такое расположение нижнего прицветного листа свойственно также некоторым представителям семейств ситниковых и осоковых. Цветки у джонсонии мелкие, со сросшимися внизу и образующими короткую трубку сегментами околоцветника. Тычинок 3, расширенные основания их нитей срослись между собой и с околоцветником (рис. 75, 3). Столбик расположен выше уровня пыльников. Семена джонсонии снабжены элайосомами.Другой род трибы джонсониевых —- совербея (Sowerbaea) состоит из .5 близкородственных видов, дизъюнктивно распространенных по всему побережью Австралии и Тасмании. Растут они преимущественно на сырых и болотистых приморских песках и в субприбрежных верещатниках. Это небольшие, образующие дерновины многолетние травы с прикорневыми листьями, имеющими узкую 3-гранную пластинку и влагалище, а также длинный язычок, расположенный, как и у злаков, на границе между ними (рис. 75, 8 —- 9). По внешнему виду совербея походит на лук. Ее кувшинчатые фиолетовые, розоватые или синие цветки со свободными сегментами околоцветника собраны в довольно густое многоцветковое зонтиковидно-головчатое (как у луков) соцветие, окруженное общей оберткой из нескольких чешуевидных листочков. Каждый цветок располагается в пазухе сильно бахромчато рассеченного прицветника. Для рода характерно наличие 3 тычинок, супротивных внутренним сегментам околоцветника, и 3 чередующихся с ними стаминодиев. Тычинки с очень короткими нитями и удлиненными раздвоенными пыльниками. Столбик расположен значительно выше тычинок, и поэтому самоопыления у видов этого рода, по-видимому, не происходит. В каждом гнезде завязи 2 —- 6 семязачатков.Род бория (Воrуа, рис. 75, 5 —- 7) включает низкие, сильно ксерофилизованные кустарничковидные растения, с очень жесткими колючими 3-гранными листовыми пластинками и широкими короткими влагалищами. Плотные головчатые соцветия окружены черепитчатой оберткой из жестких кожистых чешуй. Супротивно кожистым прицветникам, в пазухах которых сидят цветки, на вентральной стороне последних располагаются чешуевидные прицветнички, охватывающие своими завернутыми краями нижнюю часть цветка (рис. 75, 6). Цветки мелкие, желтоватые, с околоцветником, дифференцированным на длинную трубку из сросшихся частей сегментов и колесовидный отгиб. Тычинок 6, прикрепленных тонкими нитями к зеву околоцветника, что является необычным для этого семейства. Столбик расположен на одном уровне с пыльниками. Семязачатки в каждом гнезде завязи многочисленные. В род бория входят 3 вида, встречающихся в Западной Австралии и Квинсленде. Бория блестящая (В. nitida) замечательна тем, что может сохранять жизнеспособность даже после сильного пересыхания. Она обитает в аридных районах юго-запада Австралии. Это низкий (высотой 3 —- 15 см) кустарничек с очень жесткими трехгранными игловидными листьями, скученными на верхушке побегов. Пластинки листьев со временем опадают, а их короткие расширенные черепитчато налегающие друг на друга основания остаются на ветвях, придавая им своеобразный чешуйчатый вид. Листья бории блестящей имеют ярко выраженное ксероморфное строение: массивное кольцо склеренхимы под эпидермой, очень глубокое расположение устьиц. При наступлении летней засухи этому виду приходится существовать в течение многих недель в отсутствии влаги. Что же происходит в это время с растением? Его листовые пластинки, прежде расположенные более или менее горизонтально, поднимаются вверх, образуя плотный жесткий пучок, в котором молодые листья и верхушечная почка оказываются под прикрытием старых листьев. Продольные бороздки листа с находящимися в них устьицами закрываются благодаря сцеплению зубцевидных выростов на эпидермальных клетках. Эти приспособления несколько замедляют, но не приостанавливают процесс потери воды растением. Продолжая все больше и больше обезвоживаться, листья становятся вначале ярко-оранжевыми, а затем соломенно-желтыми. Такие растения производят впечатление совершенно засохших, безжизненных. Но при наступлении периода дождей они вновь становятся зелеными. Специальные наблюдения в природе показали, что если засохшие растения, находившиеся без воды в течение нескольких недель, полить, то через 24ч после полива листья возвращаются в прежнее, почти горизонтальное положение и начинают зеленеть от основания. Через 16 суток после полива листья становятся зелеными больше чем наполовину своей длины, и на этом процесс их позеленения обычно останавливается. Экспериментально установлено, что бория блестящая может обходиться без воды в течение четырех лет!
Очень своеобразна эндемичная для Австралии триба джонсониевых, иногда выделяемая в отдельное семейство. В нее входит 8 родов, включающих многолетние травы, реже кустарничковидные формы. Многие представители трибы являются ксерофильными растениями. Характерным признаком джонсониевых является наличие головчатого, реже зонтиковидного соцветия, окруженного оберткой кроющих листьев; иногда (у рода джонсония —- Johnsonia) соцветие —- простой колос. Другая особенность джонсониевых —- унифациальные (билатеральные или 3-гранные) листовые пластинки. Листья джонсониевых расположены в прикорневых или верхушечных пучках или по всему стеблю. Цветки сидят в пазухе прицветников, причем у некоторых родов каждый цветок имеет еще и прицветничек, расположенный на его вентральной (брюшной) стороне и в разной степени охватывающий цветок (рис. 75). Сегменты околоцветника свободные или сросшиеся в основании или наполовину своей длины, образуя в последнем случае трубку и отгиб (например, у рода бория —- Воrуа). В цветке 6 или 3 тычинки; в каждом гнезде завязи от многих до нескольких семязачатков.В роде джоисония 3 вида, обитающих в Юго-Западной Австралии по сухим приморским пескам. Наиболее известна джонсония обыкновенная (J. lupulina) —- плотнодерновинный многолетник с жесткими двурядно расположенными билатеральными узколинейными листьями и уплощенным стеблем (рис. 75, 1). Джонсонию можно принять за растение из семейства злаков. Злаковидный облик имеют не только дерновины этого вида, но и его соцветие —- густой продолговато-эллиптический колос с крупными скрывающими цветки, тонкокожистыми, как у злаков, чешуями. При основании колоса имеется кроющий лист, направленный вертикально вверх, благодаря чему создается впечатление, что он является продолжением стебля, а соцветие занимает не верхушечное, а боковое положение. Такое расположение нижнего прицветного листа свойственно также некоторым представителям семейств ситниковых и осоковых. Цветки у джонсонии мелкие, со сросшимися внизу и образующими короткую трубку сегментами околоцветника. Тычинок 3, расширенные основания их нитей срослись между собой и с околоцветником (рис. 75, 3). Столбик расположен выше уровня пыльников. Семена джонсонии снабжены элайосомами.Другой род трибы джонсониевых —- совербея (Sowerbaea) состоит из .5 близкородственных видов, дизъюнктивно распространенных по всему побережью Австралии и Тасмании. Растут они преимущественно на сырых и болотистых приморских песках и в субприбрежных верещатниках. Это небольшие, образующие дерновины многолетние травы с прикорневыми листьями, имеющими узкую 3-гранную пластинку и влагалище, а также длинный язычок, расположенный, как и у злаков, на границе между ними (рис. 75, 8 —- 9). По внешнему виду совербея походит на лук. Ее кувшинчатые фиолетовые, розоватые или синие цветки со свободными сегментами околоцветника собраны в довольно густое многоцветковое зонтиковидно-головчатое (как у луков) соцветие, окруженное общей оберткой из нескольких чешуевидных листочков. Каждый цветок располагается в пазухе сильно бахромчато рассеченного прицветника. Для рода характерно наличие 3 тычинок, супротивных внутренним сегментам околоцветника, и 3 чередующихся с ними стаминодиев. Тычинки с очень короткими нитями и удлиненными раздвоенными пыльниками. Столбик расположен значительно выше тычинок, и поэтому самоопыления у видов этого рода, по-видимому, не происходит. В каждом гнезде завязи 2 —- 6 семязачатков.Род бория (Воrуа, рис. 75, 5 —- 7) включает низкие, сильно ксерофилизованные кустарничковидные растения, с очень жесткими колючими 3-гранными листовыми пластинками и широкими короткими влагалищами. Плотные головчатые соцветия окружены черепитчатой оберткой из жестких кожистых чешуй. Супротивно кожистым прицветникам, в пазухах которых сидят цветки, на вентральной стороне последних располагаются чешуевидные прицветнички, охватывающие своими завернутыми краями нижнюю часть цветка (рис. 75, 6). Цветки мелкие, желтоватые, с околоцветником, дифференцированным на длинную трубку из сросшихся частей сегментов и колесовидный отгиб. Тычинок 6, прикрепленных тонкими нитями к зеву околоцветника, что является необычным для этого семейства. Столбик расположен на одном уровне с пыльниками. Семязачатки в каждом гнезде завязи многочисленные. В род бория входят 3 вида, встречающихся в Западной Австралии и Квинсленде. Бория блестящая (В. nitida) замечательна тем, что может сохранять жизнеспособность даже после сильного пересыхания. Она обитает в аридных районах юго-запада Австралии. Это низкий (высотой 3 —- 15 см) кустарничек с очень жесткими трехгранными игловидными листьями, скученными на верхушке побегов. Пластинки листьев со временем опадают, а их короткие расширенные черепитчато налегающие друг на друга основания остаются на ветвях, придавая им своеобразный чешуйчатый вид. Листья бории блестящей имеют ярко выраженное ксероморфное строение: массивное кольцо склеренхимы под эпидермой, очень глубокое расположение устьиц. При наступлении летней засухи этому виду приходится существовать в течение многих недель в отсутствии влаги. Что же происходит в это время с растением? Его листовые пластинки, прежде расположенные более или менее горизонтально, поднимаются вверх, образуя плотный жесткий пучок, в котором молодые листья и верхушечная почка оказываются под прикрытием старых листьев. Продольные бороздки листа с находящимися в них устьицами закрываются благодаря сцеплению зубцевидных выростов на эпидермальных клетках. Эти приспособления несколько замедляют, но не приостанавливают процесс потери воды растением. Продолжая все больше и больше обезвоживаться, листья становятся вначале ярко-оранжевыми, а затем соломенно-желтыми. Такие растения производят впечатление совершенно засохших, безжизненных. Но при наступлении периода дождей они вновь становятся зелеными. Специальные наблюдения в природе показали, что если засохшие растения, находившиеся без воды в течение нескольких недель, полить, то через 24ч после полива листья возвращаются в прежнее, почти горизонтальное положение и начинают зеленеть от основания. Через 16 суток после полива листья становятся зелеными больше чем наполовину своей длины, и на этом процесс их позеленения обычно останавливается. Экспериментально установлено, что бория блестящая может обходиться без воды в течение четырех лет!
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.
.