- Отдел сине-зеленые водоросли (Cyanophyta)
- Отдел сине-зеленые водоросли (Cyanophyta)
-
Сине-зеленые водоросли (Cyanophyta), дробянки, точнее, фикохромовые дробянки (Schizophyceae), слизевые водоросли (Myxophyceae) — сколько различных названий получила от исследователей эта группа древнейших автотрофных растений! Страсти не утихли и до сих пор. Немало таких ученых, которые готовы исключить сине-зеленых из числа водорослей, а некоторые — вообще из царства растений. И не так, «с легкой руки», а с полной уверенностью, что они делают это на серьезной научной основе. «Виноваты» в такой судьбе сине-зеленые водоросли сами. Крайне своеобразное строение клеток, колоний и нитей, интересная биология, большой филогенетический возраст— все эти признаки отдельно и вместе взятые дают основу для множества трактовок систематики этой группы организмов.Нет сомнений в том, что сине-зеленые водоросли — старейшая группа среди автотрофных организмов и среди организмов вообще. Остатки подобных им организмов найдены среди строматолитов (известковые образования с бугорчатой поверхностью и концентрически слоистым внутренним строением из докембрийских отложений), возраст которых составлял около трех миллиардов лет. Химический анализ обнаружил в этих остатках продукты разложения хлорофилла. Второе серьезное доказательство древности сине-зеленых водорослей — строение их клеток. Вместе с бактериями они объединены в одну группу под названием доядерных организмов (Procaryota). Разные систематики по-разному оценивают ранг этой группы — от класса до самостоятельного царства организмов, в зависимости от того, какое значение они придают отдельным признакам или уровню клеточного строения. В систематике сине-зеленых водорослей еще много неясного, большие разногласия возникают на каждом уровне их исследования.Сине-зеленые водоросли встречаются во всевозможных и почти невозможных для существования местообитаниях, по всем континентам и водоемам Земли.Строение клеток. По форме вегетативных клеток сине-зеленые водоросли можно разделить на две основные группы:1) виды с более или менее шаровидными клетками (шаровидные, широкоэллипсоидные, груше- и яйцевидные);2) виды с клетками, сильно вытянутыми (или сжатыми) в одном направлении (удлиненно-эллипсоидные, веретеновидные, цилиндрические — от короткоцилиндрических и бочонковидных до удлиненно-цилиндрических). Клетки живут отдельно, а иногда соединяются в колонии или образуют нити (последние также могут жить отдельно или образовывать дерновинки или студенистые колонии).Клетки имеют довольно толстые стенки. В сущности, протопласт окружен здесь четырьмя оболочковыми слоями: двухслойная клеточная оболочка покрыта сверху внешней волнистой мембраной, а между протопластом и оболочкой находится еще и внутренняя клеточная мембрана. В образовании поперечной перегородки между клетками в нитях участвуют только внутренний слой оболочки и внутренняя мембрана; внешняя мембрана и внешний слой оболочки туда не заходят.Строение клеточной стенки и другие микроструктуры клеток сине-зеленых водорослей изучали с помощью электронного микроскопа (рис. 49).
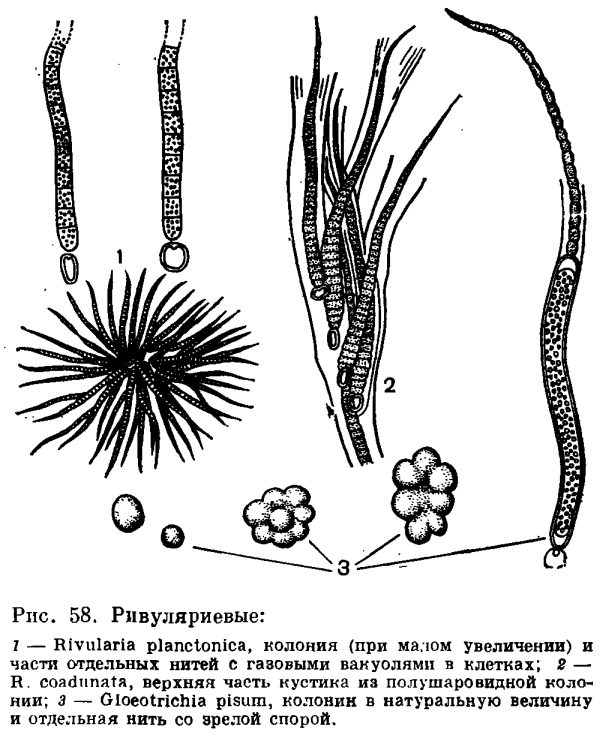
 В клеточной оболочке хотя и содержится целлюлоза, но основную роль играют пектиновые вещества и слизевые полисахариды. У одних видов клеточные оболочки хорошо ослизняются и содержат даже пигменты; у других вокруг клеток образуется специальный слизистый чехол, иногда самостоятельный вокруг каждой клетки, но чаще сливающийся в общий чехол, окружающий группу или весь ряд клеток, называемый у нитчатых форм специальным термином — трихом. У многих сине-зеленых водорослей трихомы окружены настоящими чехлами — влагалищами. Как клеточные, так и настоящие чехлы состоят из тонких переплетающихся волокон. Они могут быть гомогенными или слоистыми: слоистость у нитей с обособленными основаниями и верхушкой бывает параллельной или косой, иногда даже воронкообразной. Настоящие чехлы растут путем наложения новых слоев слизи друг на друга или внедрения новых слоев между старыми. У некоторых ностоковых (Nostoc, Anabaena) клеточные чехлы образуются путем выделения слизи через поры в оболочках.Протопласт сине-зеленых водорослей лишен оформленного ядра и ранее считался диффузным, разделенным лишь на окрашенную периферическую часть — хроматоплазму — и лишенную окраски центральную часть — цептроплазму. Однако различными методами микроскопии и цитохимии, а также ультрацентрифугированием было доказано, что такое разделение может быть только условным. Клетки сине-зеленых водорослей содержат хорошо выраженные структурные элементы, и разное их расположение обусловливает различия между центро- и хроматоплазмой. Некоторые авторы выделяют теперь в протопласте сине-зеленых водорослей три составные части:1) нуклеоплазму;2) фотосинтетические пластины (ламеллы);3) рибосомы и другие цитоплазматические гранулы.Но так как нуклеоплазма занимает район центроплазмы, а ламеллы и другие составные части расположены в районе хроматоплазмы, содержащей пигменты, то нельзя считать ошибкой и старое, классическое разграничение (рибосомы встречаются в обеих частях протопласта).Пигменты, сосредоточенные в периферической части протопласта, локализованы в пластинчатых образованиях — ламеллах, которые располагаются в хроматоплазме по-разному: хаотично, бывают упакованы в гранулы или ориентированы радиально. Подобные системы ламелл теперь нередко называют парахроматофорами.В хроматоплазме, кроме ламелл и рибосом, встречаются еще эктопласты (цианофициновые зерна, состоящие из липопротеидов) и различного рода кристаллы. В зависимости от физиологического состояния и возраста клеток все эти структурные элементы могут сильно изменяться вплоть до полного исчезновения.Центроплазма клеток сине-зеленых водорослей состоит из гиалоплазмы и разнообразных палочек, фибрилл и гранул. Последние представляют собой хроматиновые элементы, которые окрашиваются ядерными красителями. Гиалоплазму и хроматиновые элементы вообще можно считать аналогом ядра, поскольку в этих элементах содержится ДНК; они при делении клеток делятся продольно, и половинки поровну распределяются по дочерним клеткам. Но, в отличие от типичного ядра, в клетках сине-зеленых водорослей вокруг хроматиновых элементов никогда не удается обнаружить ядерной оболочки и ядрышек. Это — ядроподобное образование в клетке, и называют его нуклеоидом. В нем встречаются и рибосомы, содержащие РНК, вакуоли и полифосфатные гранулы.Установлено, что у нитчатых форм между клетками имеются плазмодесмы. Иногда соединены между собой также системы ламелл соседних клеток. Поперечные перегородки в трихоме ни в коем случае нельзя считать кусками мертвого вещества. Это живая составная часть клетки, которая постоянно участвует в ее жизненных процессах подобно перипласту жгутиковых организмов.Протоплазма сине-зеленых водорослей более густая, чем у других групп растений; она неподвижна и очень редко содержит вакуоли, наполненные клеточным соком. Вакуоли появляются только в старых клетках, и возникновение их всегда приводит к гибели клетки. Зато в клетках сине-зеленых водорослей часто встречаются газовые вакуоли (псевдовакуоли). Это полости в протоплазме, наполненные азотом и придающие клетке в проходящем свете микроскопа черно-бурый или почти черный цвет. Встречаются они у некоторых видов почти постоянно, но есть и такие виды, у которых их не обнаруживают. Присутствие или отсутствие их часто считается таксономически важным признаком, но, конечно, еще далеко не все о газовых вакуолях нам известно. Чаще всего встречаются они в клетках у таких видов, которые ведут планктонный образ жизни (представители родов Anabaena, Aphanizomenon, Rivularia, Microcystis и др., рис. 50, 58,1).
В клеточной оболочке хотя и содержится целлюлоза, но основную роль играют пектиновые вещества и слизевые полисахариды. У одних видов клеточные оболочки хорошо ослизняются и содержат даже пигменты; у других вокруг клеток образуется специальный слизистый чехол, иногда самостоятельный вокруг каждой клетки, но чаще сливающийся в общий чехол, окружающий группу или весь ряд клеток, называемый у нитчатых форм специальным термином — трихом. У многих сине-зеленых водорослей трихомы окружены настоящими чехлами — влагалищами. Как клеточные, так и настоящие чехлы состоят из тонких переплетающихся волокон. Они могут быть гомогенными или слоистыми: слоистость у нитей с обособленными основаниями и верхушкой бывает параллельной или косой, иногда даже воронкообразной. Настоящие чехлы растут путем наложения новых слоев слизи друг на друга или внедрения новых слоев между старыми. У некоторых ностоковых (Nostoc, Anabaena) клеточные чехлы образуются путем выделения слизи через поры в оболочках.Протопласт сине-зеленых водорослей лишен оформленного ядра и ранее считался диффузным, разделенным лишь на окрашенную периферическую часть — хроматоплазму — и лишенную окраски центральную часть — цептроплазму. Однако различными методами микроскопии и цитохимии, а также ультрацентрифугированием было доказано, что такое разделение может быть только условным. Клетки сине-зеленых водорослей содержат хорошо выраженные структурные элементы, и разное их расположение обусловливает различия между центро- и хроматоплазмой. Некоторые авторы выделяют теперь в протопласте сине-зеленых водорослей три составные части:1) нуклеоплазму;2) фотосинтетические пластины (ламеллы);3) рибосомы и другие цитоплазматические гранулы.Но так как нуклеоплазма занимает район центроплазмы, а ламеллы и другие составные части расположены в районе хроматоплазмы, содержащей пигменты, то нельзя считать ошибкой и старое, классическое разграничение (рибосомы встречаются в обеих частях протопласта).Пигменты, сосредоточенные в периферической части протопласта, локализованы в пластинчатых образованиях — ламеллах, которые располагаются в хроматоплазме по-разному: хаотично, бывают упакованы в гранулы или ориентированы радиально. Подобные системы ламелл теперь нередко называют парахроматофорами.В хроматоплазме, кроме ламелл и рибосом, встречаются еще эктопласты (цианофициновые зерна, состоящие из липопротеидов) и различного рода кристаллы. В зависимости от физиологического состояния и возраста клеток все эти структурные элементы могут сильно изменяться вплоть до полного исчезновения.Центроплазма клеток сине-зеленых водорослей состоит из гиалоплазмы и разнообразных палочек, фибрилл и гранул. Последние представляют собой хроматиновые элементы, которые окрашиваются ядерными красителями. Гиалоплазму и хроматиновые элементы вообще можно считать аналогом ядра, поскольку в этих элементах содержится ДНК; они при делении клеток делятся продольно, и половинки поровну распределяются по дочерним клеткам. Но, в отличие от типичного ядра, в клетках сине-зеленых водорослей вокруг хроматиновых элементов никогда не удается обнаружить ядерной оболочки и ядрышек. Это — ядроподобное образование в клетке, и называют его нуклеоидом. В нем встречаются и рибосомы, содержащие РНК, вакуоли и полифосфатные гранулы.Установлено, что у нитчатых форм между клетками имеются плазмодесмы. Иногда соединены между собой также системы ламелл соседних клеток. Поперечные перегородки в трихоме ни в коем случае нельзя считать кусками мертвого вещества. Это живая составная часть клетки, которая постоянно участвует в ее жизненных процессах подобно перипласту жгутиковых организмов.Протоплазма сине-зеленых водорослей более густая, чем у других групп растений; она неподвижна и очень редко содержит вакуоли, наполненные клеточным соком. Вакуоли появляются только в старых клетках, и возникновение их всегда приводит к гибели клетки. Зато в клетках сине-зеленых водорослей часто встречаются газовые вакуоли (псевдовакуоли). Это полости в протоплазме, наполненные азотом и придающие клетке в проходящем свете микроскопа черно-бурый или почти черный цвет. Встречаются они у некоторых видов почти постоянно, но есть и такие виды, у которых их не обнаруживают. Присутствие или отсутствие их часто считается таксономически важным признаком, но, конечно, еще далеко не все о газовых вакуолях нам известно. Чаще всего встречаются они в клетках у таких видов, которые ведут планктонный образ жизни (представители родов Anabaena, Aphanizomenon, Rivularia, Microcystis и др., рис. 50, 58,1).
, Нет сомнения в том, что газовые вакуоли у этих водорослей служат своеобразным приспособлением к уменьшению удельного веса, т. е. к улучшению «парения» в толще воды. И все же их наличие совсем необязательно, и даже у таких типичных планктеров, как Microcystis aeruginosa и М. flosaquae, можно наблюдать (особенно осенью) почти полное исчезновение газовых вакуолей. У некоторых видов они появляются и исчезают внезапно, часто по неизвестным причинам. У ностока сливовидного (Nostoc pruniforme, табл. 3, 9), крупные колонии которого всегда живут на дне водоемов, они появляются в природных условиях весной, вскоре после таяния льда. Обычно зеленоватокоричневые колонии приобретают тогда сероватый, иногда даже молочный оттенок и в течение нескольких дней полностью расплываются. Микроскопирование водоросли в зтой стадии показывает, что все клетки ностока набиты газовыми вакуолями (рис. 50) и стали черновато-коричневыми, похожими на клетки планктонных анабен. В зависимости от условий газовые вакуоли сохраняются до десяти дней, но в конце концов исчезают; начинается образование слизистого чехла вокруг клеток и их интенсивное деление. Каждая нить или даже кусок нити дает начало новому организму (колонии). Подобную картину можно наблюдать и при прорастании спор эпифитных или планктонных видов глеотрихии. Иногда газовые вакуоли появляются только в некоторых клетках трихома, например в меристемальпой зоне, где происходит интенсивное деление клеток и могут возникать гормогонии, выходу которых газовые вакуоли каким-то образом помогают.
Нет сомнения в том, что газовые вакуоли у этих водорослей служат своеобразным приспособлением к уменьшению удельного веса, т. е. к улучшению «парения» в толще воды. И все же их наличие совсем необязательно, и даже у таких типичных планктеров, как Microcystis aeruginosa и М. flosaquae, можно наблюдать (особенно осенью) почти полное исчезновение газовых вакуолей. У некоторых видов они появляются и исчезают внезапно, часто по неизвестным причинам. У ностока сливовидного (Nostoc pruniforme, табл. 3, 9), крупные колонии которого всегда живут на дне водоемов, они появляются в природных условиях весной, вскоре после таяния льда. Обычно зеленоватокоричневые колонии приобретают тогда сероватый, иногда даже молочный оттенок и в течение нескольких дней полностью расплываются. Микроскопирование водоросли в зтой стадии показывает, что все клетки ностока набиты газовыми вакуолями (рис. 50) и стали черновато-коричневыми, похожими на клетки планктонных анабен. В зависимости от условий газовые вакуоли сохраняются до десяти дней, но в конце концов исчезают; начинается образование слизистого чехла вокруг клеток и их интенсивное деление. Каждая нить или даже кусок нити дает начало новому организму (колонии). Подобную картину можно наблюдать и при прорастании спор эпифитных или планктонных видов глеотрихии. Иногда газовые вакуоли появляются только в некоторых клетках трихома, например в меристемальпой зоне, где происходит интенсивное деление клеток и могут возникать гормогонии, выходу которых газовые вакуоли каким-то образом помогают.
,Газовые вакуоли образуются на границе хромато- и центроплазмы и по очертаниям совсем неправильны. У некоторых видов, живущих в верхних слоях придонного ила (в сапропеле), в частности у видов осциллатории, крупные газовые вакуоли располагаются в клетках по сторонам поперечных перегородок. Экспериментально установлено, что появление таких вакуолей бывает вызвано уменьшением в среде количества растворенного кислорода, с прибавлением в среду продуктов сероводородного брожения. Можно предполагать, что такие вакуоли возникают в качестве хранилищ или мест отложения газов, которые выделяются при ферментативных процессах, происходящих в клетке.Состав пигментного аппарата у сине-зеленых водорослей очень пестрый, у них найдено около 30 различных внутриклеточных пигментов. Они относятся к четырем группам — к хлорофиллам, каротинам, ксантофиллам и билипротеинам. Из хлорофиллов пока достоверно доказано наличие хлорофилла а; из каротиноидов — α, β и ε-каротинов; из ксантофиллов — эхинеона, зеаксантина, криптоксаитина, миксоксантофилла и др., а из билипротеинов — с-фикоцианина, с-фикоэритрина и аллофикоцианина. Весьма характерно для синезеленых водорослей наличие последней группы пигментов (встречающейся еще у багрянок и некоторых криптомонад) и отсутствие хлорофилла b. Последнее еще раз свидетельствует о том, что сине-зеленые водоросли являются древней группой, отделившейся и пошедшей по самостоятельному пути развития еще до возникновения в ходе эволюции хлорофилла b, участие которого в фотохимических реакциях фотосинтеза дает наиболее высокий коэффициент полезного действия.Разнообразием и своеобразным составом фотоассимилирующих пигментных систем объясняется устойчивость сине-зеленых водорослей к воздействию продолжительного затемнения и анаэробиоза. Этим же частично объясняется и существование их в крайних условиях обитания — в пещерах, богатых сероводородом слоях придонного ила, в минеральных источниках.Продуктом фотосинтеза в клетках сипе-зеленых водорослей является гликопротеид, который возникает в хроматоплазме и там же отлагается. Гликопротеид похож на гликоген — от раствора иода в йодистом калии он приобретает коричневый цвет. Между фотосинтетическими ламеллами обнаружены полисахаридные зернышки. Цианофициновые зерна во внешнем слое хроматоплазмы состоят из липопротеидов. Волютиновые зерна в центроплазме представляют собой запасные вещества белкового происхождения. В плазме обитателей серных водоемов появляются зернышки серы.Пестротой пигментного состава можно объяснить и разнообразие цвета клеток и трихомов сине-зеленых водорослей. Окраска их варьирует от чисто-сине-зеленой до фиолетовой или красноватой, иногда до пурпурной или коричневато-красной, от желтой до бледно-голубой или почти черной. Цвет протопласта зависит от систематического положения вида, а также от возраста клеток и условий существования. Очень часто он маскируется цветом слизистых влагалищ или колониальной слизи. Пигменты встречаются и в слизи и придают нитям или колониям желтый, коричневый, красноватый, фиолетовый или синий оттенок. Цвет слизи, в свою очередь, зависит от экологических условий — от света, химизма и рН среды, от количества влаги в воздухе (у аэрофитов).Строение нитей. Немногие сине-зеленые водоросли растут в виде отдельных клеток, большинству свойственно образование колоний или многоклеточных нитей. В свою очередь, нити могут или образовывать ложнопаренхимпые колонии, в которых они тесно сомкнуты, а клетки сохраняют физиологическую самостоятельность, или иметь гормогониальное строение, при котором клетки соединены в ряд, слагая так называемый трихом. В трихоме протопласты соседних клеток соединены плазмодесмами. Трихом, окруженный слизистым влагалищем, получил название нити.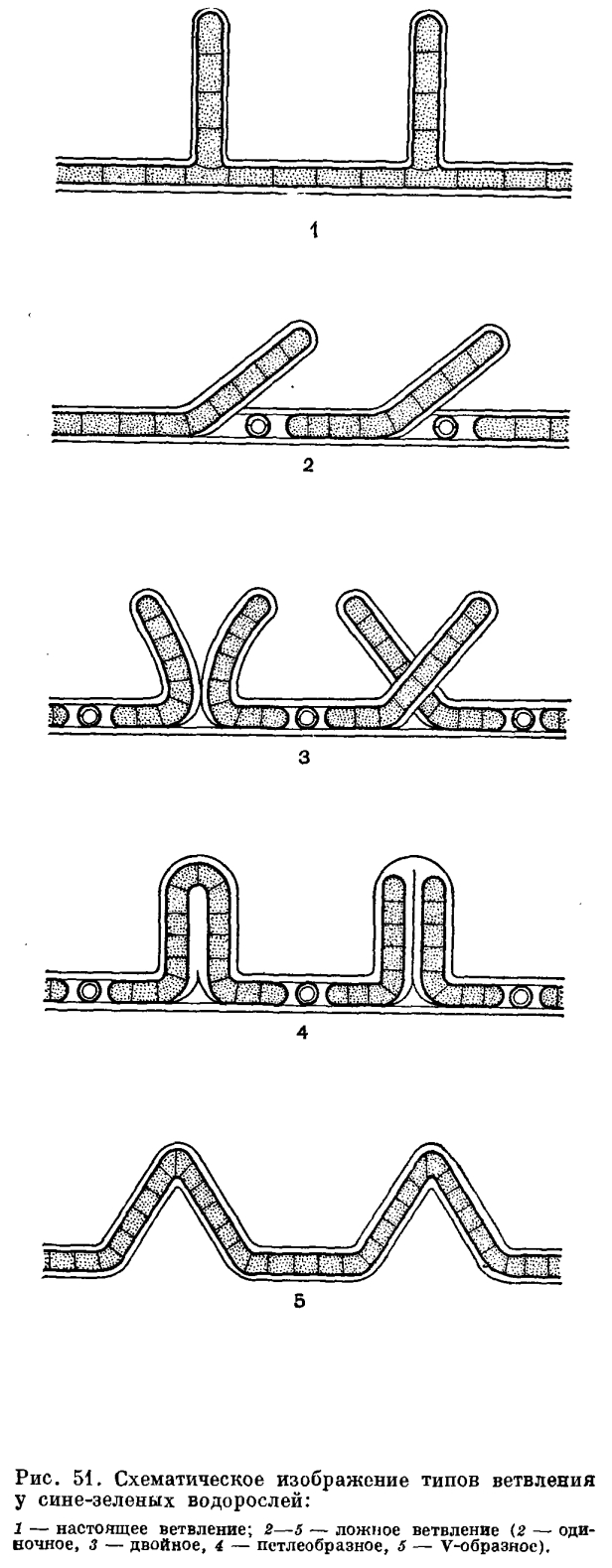 Нитчатые формы могут быть простые и разветвленные. Ветвление у сине-зеленых водорослей бывает двояким — настоящим и ложным (рис. 51). Настоящим называют такое ветвление, когда боковая ветвь возникает в результате деления одной клетки перпендикулярно к основной нити (порядок Stigonematales). Ложным ветвлением называют образование боковой ветви путем разрыва трихома и прорыва его через влагалище в сторону одним или обоими концами. В первом случае говорят об одиночном, во втором — о двойном (или парном) ложном ветвлении. Ложным ветвлением можно считать и петлеобразное ветвление, характерное для семейства Scytonemataceae, и редко встречающееся V-образное ветвление — результат повторного деления и роста двух соседних клеток трихома в двух взаимно противоположных направлениях по отношению к длинной оси нити.У очень многих нитчатых сине-зеленых водорослей имеются своеобразные клетки, получившие название гетероцист. У них хорошо выражена двухслойная оболочка, а содержимое всегда лишено ассимиляционных пигментов (оно бесцветное, голубоватое или желтоватое), газовых вакуолей и зерен запасных веществ. Они образуются из вегетативных клеток в разных местах трихома, в зависимости от систематического положения водоросли: на одном (Rivularia, Calothrix, Gloeotrichia) и обоих (Anabaenopsis, Cylindrospermum) концах трихома — базально и терминально; в трихоме между вегетативными клетками, т. е. интеркалярно (Nostoc, Anabaena, Nodularia) или сбоку трихома — латерально (у некоторых Stigonematales). Гетероцисты встречаются поодиночке или по нескольку (2—10) в ряд. В зависимости от расположения в каждой гетероцисте возникают одна (у терминальных и латеральных гетероцист) или две, изредка даже три (у интеркалярных) пробки, которые с внутренней стороны закупоривают поры между гетероцистой и соседними вегетативными клетками (рис. 5, 2).
Нитчатые формы могут быть простые и разветвленные. Ветвление у сине-зеленых водорослей бывает двояким — настоящим и ложным (рис. 51). Настоящим называют такое ветвление, когда боковая ветвь возникает в результате деления одной клетки перпендикулярно к основной нити (порядок Stigonematales). Ложным ветвлением называют образование боковой ветви путем разрыва трихома и прорыва его через влагалище в сторону одним или обоими концами. В первом случае говорят об одиночном, во втором — о двойном (или парном) ложном ветвлении. Ложным ветвлением можно считать и петлеобразное ветвление, характерное для семейства Scytonemataceae, и редко встречающееся V-образное ветвление — результат повторного деления и роста двух соседних клеток трихома в двух взаимно противоположных направлениях по отношению к длинной оси нити.У очень многих нитчатых сине-зеленых водорослей имеются своеобразные клетки, получившие название гетероцист. У них хорошо выражена двухслойная оболочка, а содержимое всегда лишено ассимиляционных пигментов (оно бесцветное, голубоватое или желтоватое), газовых вакуолей и зерен запасных веществ. Они образуются из вегетативных клеток в разных местах трихома, в зависимости от систематического положения водоросли: на одном (Rivularia, Calothrix, Gloeotrichia) и обоих (Anabaenopsis, Cylindrospermum) концах трихома — базально и терминально; в трихоме между вегетативными клетками, т. е. интеркалярно (Nostoc, Anabaena, Nodularia) или сбоку трихома — латерально (у некоторых Stigonematales). Гетероцисты встречаются поодиночке или по нескольку (2—10) в ряд. В зависимости от расположения в каждой гетероцисте возникают одна (у терминальных и латеральных гетероцист) или две, изредка даже три (у интеркалярных) пробки, которые с внутренней стороны закупоривают поры между гетероцистой и соседними вегетативными клетками (рис. 5, 2).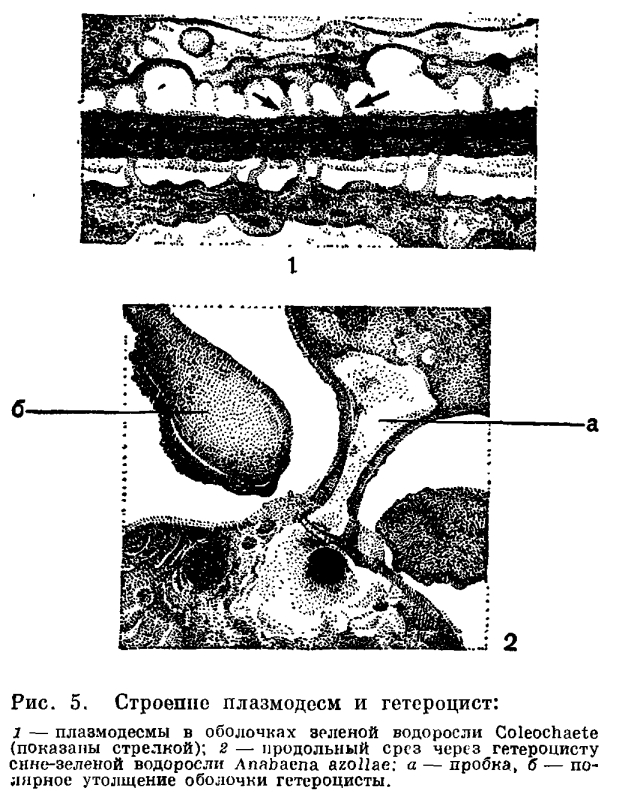 Гетероцисты называют ботанической загадкой. В световом микроскопе они выглядят как будто пустыми, но иногда, к большому удивлению исследователей, они вдруг прорастали, давая начало новым трихомам. При ложном ветвлении и во время разделения нитей трихомы чаще всего разрываются возле гетероцист, как будто ими ограничивается рост трихомов. Благодаря этому их раньше называли пограничными клетками. Нити с базальными и терминальными гетероцистами прикрепляются к субстрату при помощи гетероцист. У некоторых видов с гетероцистами связано образование покоящихся клеток — спор: они располагаются рядом с гетероцистой по одну (у Суlindrospermum, Gloeotrichia, Anabaenopsis raciborskii) или по обе ее стороны (у некоторых Anabaena). Не исключено, что гетероцисты являются хранилищами каких-то запасных веществ или энзимов. Любопытно отметить, что все виды сине-зеленых водорослей, способные фиксировать атмосферный азот, имеют гетероцисты.Размножение. Самым обычным типом размножения у сине-зеленых водорослей является деление клеток надвое. Для одноклеточных форм этот способ единственный; в колониях и нитях он приводит к росту нити или колонии.Трихом образуется тогда, когда делящиеся в одном направлении клетки не отходят друг от друга. При нарушении линейного расположения возникает колония с беспорядочно расположенными клетками. При делении в двух перпендикулярных направлениях в одной плоскости образуется пластинчатая колония с правильным расположением клеток в виде тетрад (Merismopedia). Объемные скопления в виде пакетов возникают в том случае, когда клетки делятся в трех плоскостях (Eucapsis).Представителям некоторых родов (Gloeocapsa, Microcystis) свойственно также быстрое деление с образованием в материнской клетке множества мелких клеток — нанноцитов.Сине-зеленые водоросли размножаются и другими способами — образованием спор (покоящихся клеток), экзо- и эндоспор, гормогониев, гормоспор, гонидиев, кокков и планококков. Одним из самых распространенных видов размножения нитчатых форм является образование гормогониев. Этот способ размножения столь характерен для части синезеленых водорослей, что послужил названием целому классу гормогониевых (Hormogoniophyсеае). Гормогониями принято называть фрагменты трихома, на которые последний распадается. Образование гормогониев — не просто механическое отделение группы из двух, трех или большего числа клеток. Гормогонии обособляются благодаря отмиранию некоторых некроидальных клеток, затем с помощью выделения слизи они выскальзывают из влагалища (если оно имеется) и, совершая колебательные движения, перемещаются в воде или по субстрату. Каждый гормогонии может дать начало новой особи. Если группа клеток, похожая на гормогонии, одета толстой оболочкой, ее называют гормоспорой (гормоцистой), которая одновременно выполняет функции и размножения, и перенесения неблагоприятных условий.У некоторых видов от таллома отделяются одноклеточные фрагменты, которые названы гонидиями, кокками или планококками. Гонидии сохраняют слизистую оболочку; кокки лишены ясно выраженных оболочек; планококки тоже голые, но, подобно гормогониям, обладают способностью к активному движению.Причины движения гормогониев, планококков и целых трихомов (у Oscillatoriaceae) далеко еще не выяснены. Они скользят вдоль продольной оси, колеблясь из стороны в сторону, или вращаются вокруг нее. Движущей силой считают выделение слизи, сокращение трихомов по направлению продольной оси, сокращения внешней волнистой мембраны, а также злектрокинетические явления.Довольно распространенными органами размножения являются споры, особенно у водорослей из порядка Nostocales. Они одноклеточные, обычно крупнее вегетативных клеток и возникают из них, чаще из одной. Однако у представителей некоторых родов (Gloeotrichia, Anabaena) они образуются в результате слияния нескольких вегетативных клеток, и длина таких спор может достигать 0,5 мм. Не исключено, что в процессе такого слияния происходит и рекомбинация, но пока точных данных об этом нет.Споры покрыты толстой, двухслойной оболочкой, внутренний слой которой называют эндоспорием, а наружный — экзоспорием. Оболочки гладкие или усеяны сосочками, бесцветные, желтые или коричневатые. Благодаря толстым оболочкам и физиологическим изменениям в протопласте (накопление запасных веществ, исчезновение ассимиляционных пигментов, иногда увеличение количества цианофициновых зерен) споры могут длительное время сохранять жизнеспособность в неблагоприятных условиях и при разнообразных сильных воздействиях (при низких и высоких температурах, при высыхании и сильном облучении). В благоприятных условиях спора прорастает, ее содержимое делится на клетки — образуются спорогормогонии, оболочка ослизняется, разрывается или открывается крышкой и гормогонии выходит.Эндо- и экзоспоры встречаются главным образом у представителей класса хамесифоновых (Chamaesiphonophyceae). Эндоспоры образуются в увеличенных материнских клетках в большом количестве (свыше ста). Образование их происходит сукцеданно (в результате ряда последовательных делений протопласта материнской клетки) или симультанно (путем одновременного распадения материнской клетки на многие мелкие клетки). Экзоспоры по мере своего образовапия отчленяются от протопласта материнской клетки и выходят наружу. Иногда они не отделяются от материнской клетки, а образуют на ней цепочки (например, у некоторых видов Chamaesiphon).Половое размножение у сине-зеленых водорослей полностью отсутствует.Способы питания и экология. Известно, что большинство сине-зеленых водорослей способно синтезировать все вещества своей клетки за счет энергии света. Фотосинтетические процессы, происходящие в клетках сине-зеленых водорослей, в своей принципиальной схеме близки процессам, которые совершаются в других хлорофиллсодержащих организмах.Фотоавтотрофный тип питания является для них основным, но не единственным. Кроме настоящего фотосинтеза, сине-зеленые водоросли способны к фоторедукции, фотогетеротрофии, автогетеротрофии, гетероавтотрофии и даже полной гетеротрофии. При наличии в среде органических веществ они используют и их в качестве дополнительных источников энергии. Благодаря способности к смешанному (миксотрофному) питанию они могут быть активными и в крайних для фотоавтотрофной жизни условиях. В подобных местообитаниях почти полностью отсутствует конкуренция, и сине-зеленые водоросли занимают доминирующее место.В условиях плохой освещенности (в пещерах, в глубинных горизонтах водоемов) в клетках сине-зеленых водорослей изменяется пигментный состав. Это явление, получившее название хроматической адаптации, представляет собой приспособительное изменение окраски водорослей под влиянием изменения спектрального состава света за счет увеличения количества пигментов, имеющих окраску, дополнительную к цвету падающих лучей. Изменения окраски клеток (хлорозы) происходят и в случае недостатка в среде некоторых компонентов, в присутствии токсических веществ, а также при переходе к гетеротрофному типу питания.Есть среди сине-зеленых водорослей и такая группа видов, подобной которой среди других организмов вообще мало. Эти водоросли способны фиксировать атмосферный азот, и это свойство сочетается у них с фотосинтезом. Сейчас известно уже около ста таких видов. Как уже указывалось, зта способность свойственна только водорослям, имеющим гетероцисты, да и им не всем.Большинство сине-зеленых водорослей-азотфиксаторов приурочено к наземным местообитаниям. Не исключено, что именно их относительная пищевая независимость как фиксаторов атмосферного азота позволяет им заселять необитаемые, без малейших следов почвы, скалы, как это наблюдалось на острове Кракатау в 1883 г.: через три года после извержения вулкана на пепле и туфах были найдены слизистые скопления, состоящие из представителей родов Anabaena, Gloeocapsa, Nostoc, Calothrix, Phormidium и др. Первыми поселенцами острова Сурцей, возникшего в результате извержения подводного вулкана в 1963 г. около южного берега Исландии, были тоже азотфиксаторы. Среди них оказались некоторые широко распространенные планктонные виды, вызывающие «цветение» воды (Anabaena circinalis, А. cylindrica, A. flos-aquae, A. lemmermannii, А. scheremetievii, A. spiroides, Anabaenopsis circularis, Gloeotrichia echinulata).Максимальной температурой для существования живой и ассимилирующей клетки считают +65°С, но это не предел для сине-зеленых водорослей (см. очерк о водорослях горячих источников). Такую высокую температуру термофильные сине-зеленые водоросли переносят благодаря своеобразному коллоидному состоянию протоплазмы, которая при высокой температуре очень медленно коагулирует. Самыми распространенными термофилами являются космополиты Mastigocladus laminosus, Phormidium laminosum. Сине-зеленые водоросли способны выдерживать и низкую температуру. Некоторые виды без повреждения хранились в течение недели при температуре жидкого воздуха (—190°С). В природе такой температуры нет, но в Антарктиде при температуре —83°С были в большом количестве найдены сине-зеленые водоросли (ностоки).В Антарктиде и в высокогорьях, кроме низкой температуры, на водоросли влияет еще и высокая солнечная радиация. Для снижения вредного влияния коротковолнового радиационного излучения сине-зеленые водоросли в ходе эволюции приобрели ряд приспособлений. Важнейшим из них является выделение слизи вокруг клеток. Слизь колоний и слизистые влагалища нитчатых форм являются хорошей защитной обверткой, предохраняющей клетки от высыхания и одновременно действующей как фильтр, устраняющий вредное влияние радиации. В зависимости от интенсивности света в слизи отлагается больше или меньше пигмента, и она окрашивается по всей толще или по слоям.Способность слизи быстро поглощать и длительно удерживать воду позволяет сине-зеленым водорослям нормально вегетировать и в пустынных районах. Слизь поглощает максимальное количество ночной или утренней влаги, колонии набухают, и в клетках начинается ассимиляция. К полудню студенистые колонии или скопления клеток высыхают и превращаются в черные хрустящие корочки. В таком состоянии они держатся до следующей ночи, когда снова начинается поглощение влаги.Для активной жизни им вполне достаточно парообразной воды.Сине-зеленые водоросли весьма обычны в почве и в напочвенных сообществах, встречаются они и в сырых местообитаниях, а также на коре деревьев, на камнях и т. п. Все эти местообитания часто не постоянно обеспечены влагой и неравномерно освещены (подробнее см. в очерках о наземных и почвенных водорослях).Сине-зеленые водоросли встречаются также в криофильных сообществах — на льдах и на снегу. Фотосинтез возможен, конечно, только в том случае, когда клетки окружены прослойкой жидкой воды, что и происходит здесь при ярком солнечном освещении снега и льда.Солнечная радиация на ледниках и снежниках очень интенсивна, значительную часть ее составляет коротковолновое излучение, что вызывает у водорослей защитные приспособления. В группу криобионтов входит ряд видов сине-зеленых водорослей, но все же в целом представители этого отдела предпочитают местообитания с повышенной температурой (подробнее см. в очерке о водорослях снега и льда).
Гетероцисты называют ботанической загадкой. В световом микроскопе они выглядят как будто пустыми, но иногда, к большому удивлению исследователей, они вдруг прорастали, давая начало новым трихомам. При ложном ветвлении и во время разделения нитей трихомы чаще всего разрываются возле гетероцист, как будто ими ограничивается рост трихомов. Благодаря этому их раньше называли пограничными клетками. Нити с базальными и терминальными гетероцистами прикрепляются к субстрату при помощи гетероцист. У некоторых видов с гетероцистами связано образование покоящихся клеток — спор: они располагаются рядом с гетероцистой по одну (у Суlindrospermum, Gloeotrichia, Anabaenopsis raciborskii) или по обе ее стороны (у некоторых Anabaena). Не исключено, что гетероцисты являются хранилищами каких-то запасных веществ или энзимов. Любопытно отметить, что все виды сине-зеленых водорослей, способные фиксировать атмосферный азот, имеют гетероцисты.Размножение. Самым обычным типом размножения у сине-зеленых водорослей является деление клеток надвое. Для одноклеточных форм этот способ единственный; в колониях и нитях он приводит к росту нити или колонии.Трихом образуется тогда, когда делящиеся в одном направлении клетки не отходят друг от друга. При нарушении линейного расположения возникает колония с беспорядочно расположенными клетками. При делении в двух перпендикулярных направлениях в одной плоскости образуется пластинчатая колония с правильным расположением клеток в виде тетрад (Merismopedia). Объемные скопления в виде пакетов возникают в том случае, когда клетки делятся в трех плоскостях (Eucapsis).Представителям некоторых родов (Gloeocapsa, Microcystis) свойственно также быстрое деление с образованием в материнской клетке множества мелких клеток — нанноцитов.Сине-зеленые водоросли размножаются и другими способами — образованием спор (покоящихся клеток), экзо- и эндоспор, гормогониев, гормоспор, гонидиев, кокков и планококков. Одним из самых распространенных видов размножения нитчатых форм является образование гормогониев. Этот способ размножения столь характерен для части синезеленых водорослей, что послужил названием целому классу гормогониевых (Hormogoniophyсеае). Гормогониями принято называть фрагменты трихома, на которые последний распадается. Образование гормогониев — не просто механическое отделение группы из двух, трех или большего числа клеток. Гормогонии обособляются благодаря отмиранию некоторых некроидальных клеток, затем с помощью выделения слизи они выскальзывают из влагалища (если оно имеется) и, совершая колебательные движения, перемещаются в воде или по субстрату. Каждый гормогонии может дать начало новой особи. Если группа клеток, похожая на гормогонии, одета толстой оболочкой, ее называют гормоспорой (гормоцистой), которая одновременно выполняет функции и размножения, и перенесения неблагоприятных условий.У некоторых видов от таллома отделяются одноклеточные фрагменты, которые названы гонидиями, кокками или планококками. Гонидии сохраняют слизистую оболочку; кокки лишены ясно выраженных оболочек; планококки тоже голые, но, подобно гормогониям, обладают способностью к активному движению.Причины движения гормогониев, планококков и целых трихомов (у Oscillatoriaceae) далеко еще не выяснены. Они скользят вдоль продольной оси, колеблясь из стороны в сторону, или вращаются вокруг нее. Движущей силой считают выделение слизи, сокращение трихомов по направлению продольной оси, сокращения внешней волнистой мембраны, а также злектрокинетические явления.Довольно распространенными органами размножения являются споры, особенно у водорослей из порядка Nostocales. Они одноклеточные, обычно крупнее вегетативных клеток и возникают из них, чаще из одной. Однако у представителей некоторых родов (Gloeotrichia, Anabaena) они образуются в результате слияния нескольких вегетативных клеток, и длина таких спор может достигать 0,5 мм. Не исключено, что в процессе такого слияния происходит и рекомбинация, но пока точных данных об этом нет.Споры покрыты толстой, двухслойной оболочкой, внутренний слой которой называют эндоспорием, а наружный — экзоспорием. Оболочки гладкие или усеяны сосочками, бесцветные, желтые или коричневатые. Благодаря толстым оболочкам и физиологическим изменениям в протопласте (накопление запасных веществ, исчезновение ассимиляционных пигментов, иногда увеличение количества цианофициновых зерен) споры могут длительное время сохранять жизнеспособность в неблагоприятных условиях и при разнообразных сильных воздействиях (при низких и высоких температурах, при высыхании и сильном облучении). В благоприятных условиях спора прорастает, ее содержимое делится на клетки — образуются спорогормогонии, оболочка ослизняется, разрывается или открывается крышкой и гормогонии выходит.Эндо- и экзоспоры встречаются главным образом у представителей класса хамесифоновых (Chamaesiphonophyceae). Эндоспоры образуются в увеличенных материнских клетках в большом количестве (свыше ста). Образование их происходит сукцеданно (в результате ряда последовательных делений протопласта материнской клетки) или симультанно (путем одновременного распадения материнской клетки на многие мелкие клетки). Экзоспоры по мере своего образовапия отчленяются от протопласта материнской клетки и выходят наружу. Иногда они не отделяются от материнской клетки, а образуют на ней цепочки (например, у некоторых видов Chamaesiphon).Половое размножение у сине-зеленых водорослей полностью отсутствует.Способы питания и экология. Известно, что большинство сине-зеленых водорослей способно синтезировать все вещества своей клетки за счет энергии света. Фотосинтетические процессы, происходящие в клетках сине-зеленых водорослей, в своей принципиальной схеме близки процессам, которые совершаются в других хлорофиллсодержащих организмах.Фотоавтотрофный тип питания является для них основным, но не единственным. Кроме настоящего фотосинтеза, сине-зеленые водоросли способны к фоторедукции, фотогетеротрофии, автогетеротрофии, гетероавтотрофии и даже полной гетеротрофии. При наличии в среде органических веществ они используют и их в качестве дополнительных источников энергии. Благодаря способности к смешанному (миксотрофному) питанию они могут быть активными и в крайних для фотоавтотрофной жизни условиях. В подобных местообитаниях почти полностью отсутствует конкуренция, и сине-зеленые водоросли занимают доминирующее место.В условиях плохой освещенности (в пещерах, в глубинных горизонтах водоемов) в клетках сине-зеленых водорослей изменяется пигментный состав. Это явление, получившее название хроматической адаптации, представляет собой приспособительное изменение окраски водорослей под влиянием изменения спектрального состава света за счет увеличения количества пигментов, имеющих окраску, дополнительную к цвету падающих лучей. Изменения окраски клеток (хлорозы) происходят и в случае недостатка в среде некоторых компонентов, в присутствии токсических веществ, а также при переходе к гетеротрофному типу питания.Есть среди сине-зеленых водорослей и такая группа видов, подобной которой среди других организмов вообще мало. Эти водоросли способны фиксировать атмосферный азот, и это свойство сочетается у них с фотосинтезом. Сейчас известно уже около ста таких видов. Как уже указывалось, зта способность свойственна только водорослям, имеющим гетероцисты, да и им не всем.Большинство сине-зеленых водорослей-азотфиксаторов приурочено к наземным местообитаниям. Не исключено, что именно их относительная пищевая независимость как фиксаторов атмосферного азота позволяет им заселять необитаемые, без малейших следов почвы, скалы, как это наблюдалось на острове Кракатау в 1883 г.: через три года после извержения вулкана на пепле и туфах были найдены слизистые скопления, состоящие из представителей родов Anabaena, Gloeocapsa, Nostoc, Calothrix, Phormidium и др. Первыми поселенцами острова Сурцей, возникшего в результате извержения подводного вулкана в 1963 г. около южного берега Исландии, были тоже азотфиксаторы. Среди них оказались некоторые широко распространенные планктонные виды, вызывающие «цветение» воды (Anabaena circinalis, А. cylindrica, A. flos-aquae, A. lemmermannii, А. scheremetievii, A. spiroides, Anabaenopsis circularis, Gloeotrichia echinulata).Максимальной температурой для существования живой и ассимилирующей клетки считают +65°С, но это не предел для сине-зеленых водорослей (см. очерк о водорослях горячих источников). Такую высокую температуру термофильные сине-зеленые водоросли переносят благодаря своеобразному коллоидному состоянию протоплазмы, которая при высокой температуре очень медленно коагулирует. Самыми распространенными термофилами являются космополиты Mastigocladus laminosus, Phormidium laminosum. Сине-зеленые водоросли способны выдерживать и низкую температуру. Некоторые виды без повреждения хранились в течение недели при температуре жидкого воздуха (—190°С). В природе такой температуры нет, но в Антарктиде при температуре —83°С были в большом количестве найдены сине-зеленые водоросли (ностоки).В Антарктиде и в высокогорьях, кроме низкой температуры, на водоросли влияет еще и высокая солнечная радиация. Для снижения вредного влияния коротковолнового радиационного излучения сине-зеленые водоросли в ходе эволюции приобрели ряд приспособлений. Важнейшим из них является выделение слизи вокруг клеток. Слизь колоний и слизистые влагалища нитчатых форм являются хорошей защитной обверткой, предохраняющей клетки от высыхания и одновременно действующей как фильтр, устраняющий вредное влияние радиации. В зависимости от интенсивности света в слизи отлагается больше или меньше пигмента, и она окрашивается по всей толще или по слоям.Способность слизи быстро поглощать и длительно удерживать воду позволяет сине-зеленым водорослям нормально вегетировать и в пустынных районах. Слизь поглощает максимальное количество ночной или утренней влаги, колонии набухают, и в клетках начинается ассимиляция. К полудню студенистые колонии или скопления клеток высыхают и превращаются в черные хрустящие корочки. В таком состоянии они держатся до следующей ночи, когда снова начинается поглощение влаги.Для активной жизни им вполне достаточно парообразной воды.Сине-зеленые водоросли весьма обычны в почве и в напочвенных сообществах, встречаются они и в сырых местообитаниях, а также на коре деревьев, на камнях и т. п. Все эти местообитания часто не постоянно обеспечены влагой и неравномерно освещены (подробнее см. в очерках о наземных и почвенных водорослях).Сине-зеленые водоросли встречаются также в криофильных сообществах — на льдах и на снегу. Фотосинтез возможен, конечно, только в том случае, когда клетки окружены прослойкой жидкой воды, что и происходит здесь при ярком солнечном освещении снега и льда.Солнечная радиация на ледниках и снежниках очень интенсивна, значительную часть ее составляет коротковолновое излучение, что вызывает у водорослей защитные приспособления. В группу криобионтов входит ряд видов сине-зеленых водорослей, но все же в целом представители этого отдела предпочитают местообитания с повышенной температурой (подробнее см. в очерке о водорослях снега и льда).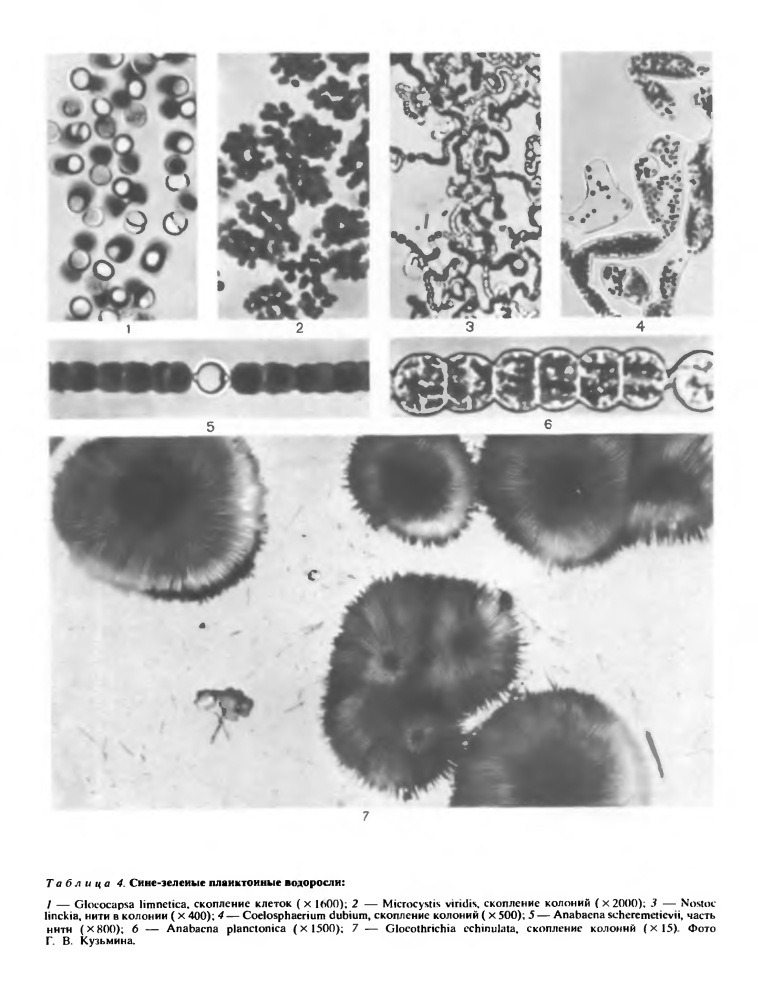 Сине-зеленые водоросли преобладают в планктоне эвтрофных (богатых питательными веществами) водоемов, где их массовое развитие часто вызывает «цветение» воды. Планктонному образу жизни этих водорослей способствуют газовые вакуоли в клетках, хотя они имеются и не у всех возбудителей «цветения» (табл. 4). Прижизненные выделения и продукты посмертного разложения у некоторых из этих синезеленых водорослей ядовиты. Массовое развитие большинства планктонных сине-зеленых водорослей начинается при высокой температуре, т. е. во второй половине весны, летом и в начале осени. Установлено, что для большинства пресноводных сине-зеленых водорослей температурный оптимум находится около+30°С. Есть и исключения. Некоторые виды осциллатории вызывают «цветение» воды подо льдом, т. е. при температуре около 0°С. Бесцветные и сероводородолюбивые виды развиваются в массовом количестве в глубинных слоях озер. Некоторые возбудители «цветения» явно выходят за границы своего ареала благодаря человеческой деятельности. Так, виды рода Anabaenopsis за пределами тропических и субтропических областей долгое время совсем не встречались, но потом были найдены в южных районах умеренного пояса, а несколько лет назад развились уже в Хельсинкской бухте. Подходящая температура и повышенная эвтрофизация (органическое загрязнение) позволили этому организму развиваться в больших количествах и севернее 60-й параллели.«Цветение» воды вообще, а вызванное синезелеными водорослями особенно считается стихийным бедствием, так как вода становится почти ни к чему уже не пригодной. При этом значительно увеличиваются вторичное загрязнение и заиление водоема, так как биомасса водорослей в «цветущем» водоеме достигает значительных величин (средняя биомасса— до 200 г/м3, максимальная — до 450—500 г/м3), а среди сине-зеленых очень мало таких видов, которые употреблялись бы другими организмами в пищу.Многосторонни отношения между сине-зелеными водорослями и другими организмами. Виды из родов Gloeocapsa, Nostoc, Scytonema, Stigonema, Rivularia и Calothrix являются фикобионтами в лишайниках. Некоторые синезеленые водоросли живут в других организмах в качестве ассимиляторов. В воздушных камерах мхов Anthoceros, Blasia живут виды Anabaena и Nostoc. В листьях водяного папоротника Azolla americana обитает Anabaena azollae, в межклетниках Cycas и Zamia—Nostoc punctiforme (подробнее см. в очерке о симбиозе водорослей с другими организмами).Таким образом, сине-зеленые водоросли встречаются на всех континентах и во всевозможных местообитаниях — в воде и на суше, в пресных и соленых водах, везде и всюду.Многие авторы придерживаются мнения, что все сине-зеленые водоросли убиквисты и космополиты, но это далеко не так. Выше уже говорилось о географическом распространении рода Anabaenopsis. Подробными исследованиями доказано, что даже такой распространенный вид, как Nostoc pruniforme, не космополит. Некоторые роды (например, Nostochopsis, Camptylonemopsis, Raphidiopsis) целиком приурочены к поясам жаркого или теплого климата, Nostoc flagelliforme — к аридным районам, многие виды рода Chamaesiphon — к холодным и чистоводным рекам и ручьям горных стран.Отдел сине-зеленых водорослей считают древнейшей группой автотрофных растений на Земле. Примитивное строение клетки, отсутствие полового размножения и жгутиковых стадий— все это серьезные доказательства их древности. По цитологии сине-зеленые сходны с бактериями, а некоторые их пигменты (билипротеины) встречаются и у красных водорослей. Однако, учитывая весь комплекс характерных для отдела признаков, можно предполагать, что сине-зеленые водоросли являются самостоятельной ветвью эволюции. Свыше трех миллиардов лет назад они отошли от основного ствола растительной эволюции и образовали тупиковую ветвь.Говоря о хозяйственном значении сине-зеленых, на первое место нужно поставить их роль в качестве возбудителей «цветения» воды. Это, к сожалению, отрицательная роль. Положительное значение их заключается прежде всего в способности усваивать свободный азот. В восточных странах сине-зеленые водоросли используют даже в пищу, а в последние годы некоторые из них нашли дорогу в бассейны массовых культур для индустриального производства органического вещества.Систематика сине-зеленых водорослей еще далека от совершенства. Сравнительная простота морфологии, относительно малое количество ценных с точки зрения систематики признаков и широкая изменчивость некоторых из них, а также разная трактовка одних и тех же признаков привели к тому, что почти все имеющиеся системы в той или иной мере субъективны и далеки от естественной. Нет хорошего, обоснованного разграничения вида как целого и объем вида в разных системах понимается по-разному. Общее количество видов в отделе определяется в 1500—2000. По принятой нами системе отдел сине-зеленых водорослей делится на 3 класса, несколько порядков и много семейств.
Сине-зеленые водоросли преобладают в планктоне эвтрофных (богатых питательными веществами) водоемов, где их массовое развитие часто вызывает «цветение» воды. Планктонному образу жизни этих водорослей способствуют газовые вакуоли в клетках, хотя они имеются и не у всех возбудителей «цветения» (табл. 4). Прижизненные выделения и продукты посмертного разложения у некоторых из этих синезеленых водорослей ядовиты. Массовое развитие большинства планктонных сине-зеленых водорослей начинается при высокой температуре, т. е. во второй половине весны, летом и в начале осени. Установлено, что для большинства пресноводных сине-зеленых водорослей температурный оптимум находится около+30°С. Есть и исключения. Некоторые виды осциллатории вызывают «цветение» воды подо льдом, т. е. при температуре около 0°С. Бесцветные и сероводородолюбивые виды развиваются в массовом количестве в глубинных слоях озер. Некоторые возбудители «цветения» явно выходят за границы своего ареала благодаря человеческой деятельности. Так, виды рода Anabaenopsis за пределами тропических и субтропических областей долгое время совсем не встречались, но потом были найдены в южных районах умеренного пояса, а несколько лет назад развились уже в Хельсинкской бухте. Подходящая температура и повышенная эвтрофизация (органическое загрязнение) позволили этому организму развиваться в больших количествах и севернее 60-й параллели.«Цветение» воды вообще, а вызванное синезелеными водорослями особенно считается стихийным бедствием, так как вода становится почти ни к чему уже не пригодной. При этом значительно увеличиваются вторичное загрязнение и заиление водоема, так как биомасса водорослей в «цветущем» водоеме достигает значительных величин (средняя биомасса— до 200 г/м3, максимальная — до 450—500 г/м3), а среди сине-зеленых очень мало таких видов, которые употреблялись бы другими организмами в пищу.Многосторонни отношения между сине-зелеными водорослями и другими организмами. Виды из родов Gloeocapsa, Nostoc, Scytonema, Stigonema, Rivularia и Calothrix являются фикобионтами в лишайниках. Некоторые синезеленые водоросли живут в других организмах в качестве ассимиляторов. В воздушных камерах мхов Anthoceros, Blasia живут виды Anabaena и Nostoc. В листьях водяного папоротника Azolla americana обитает Anabaena azollae, в межклетниках Cycas и Zamia—Nostoc punctiforme (подробнее см. в очерке о симбиозе водорослей с другими организмами).Таким образом, сине-зеленые водоросли встречаются на всех континентах и во всевозможных местообитаниях — в воде и на суше, в пресных и соленых водах, везде и всюду.Многие авторы придерживаются мнения, что все сине-зеленые водоросли убиквисты и космополиты, но это далеко не так. Выше уже говорилось о географическом распространении рода Anabaenopsis. Подробными исследованиями доказано, что даже такой распространенный вид, как Nostoc pruniforme, не космополит. Некоторые роды (например, Nostochopsis, Camptylonemopsis, Raphidiopsis) целиком приурочены к поясам жаркого или теплого климата, Nostoc flagelliforme — к аридным районам, многие виды рода Chamaesiphon — к холодным и чистоводным рекам и ручьям горных стран.Отдел сине-зеленых водорослей считают древнейшей группой автотрофных растений на Земле. Примитивное строение клетки, отсутствие полового размножения и жгутиковых стадий— все это серьезные доказательства их древности. По цитологии сине-зеленые сходны с бактериями, а некоторые их пигменты (билипротеины) встречаются и у красных водорослей. Однако, учитывая весь комплекс характерных для отдела признаков, можно предполагать, что сине-зеленые водоросли являются самостоятельной ветвью эволюции. Свыше трех миллиардов лет назад они отошли от основного ствола растительной эволюции и образовали тупиковую ветвь.Говоря о хозяйственном значении сине-зеленых, на первое место нужно поставить их роль в качестве возбудителей «цветения» воды. Это, к сожалению, отрицательная роль. Положительное значение их заключается прежде всего в способности усваивать свободный азот. В восточных странах сине-зеленые водоросли используют даже в пищу, а в последние годы некоторые из них нашли дорогу в бассейны массовых культур для индустриального производства органического вещества.Систематика сине-зеленых водорослей еще далека от совершенства. Сравнительная простота морфологии, относительно малое количество ценных с точки зрения систематики признаков и широкая изменчивость некоторых из них, а также разная трактовка одних и тех же признаков привели к тому, что почти все имеющиеся системы в той или иной мере субъективны и далеки от естественной. Нет хорошего, обоснованного разграничения вида как целого и объем вида в разных системах понимается по-разному. Общее количество видов в отделе определяется в 1500—2000. По принятой нами системе отдел сине-зеленых водорослей делится на 3 класса, несколько порядков и много семейств.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.
.