- Опыление
- Опыление
-
Со времен Ч.Дарвина до наших дней опыление орхидных не перестает привлекать внимание исследователей. И. И. Мечников в своей книге «Этюды о природе человека» (1903) называет механизм опыления орхидей одним из наиболее удивительных примеров среди гармонических явлений природы. Совершенные, иногда почти фантастические приспособления орхидей к специфическим опылителям поражают воображение. Словно изобретательный художник-конструктор забавлялся, выдумывая все новые и новые, иногда очень сложные механизмы, ярко и красочно исполненные и точно подогнанные к потребностям насекомых. Остроумные, часто обманные приспособления растений к опылению заставляют некоторых авторов говорить о «цветковой дипломатии», «сообразительности» и «безнравственности» орхидей. Опылению орхидных посвящена необозримая литература, основу которой заложил Ч. Дарвин (1862) замечательной книгой «Приспособления орхидных к оплодотворению насекомыми».
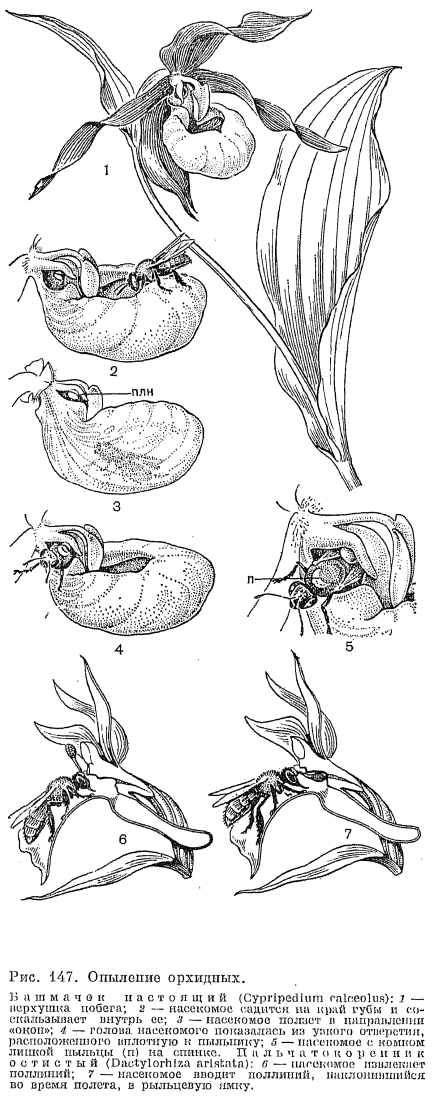
, Опылителями орхидей подсемейства циприпедиевых являются двукрылые и пчелиные. Цветки одной из наиболее декоративных за пределами тропиков орхидей венерина башмачка (Cypripedium calceolus, рис. 147, табл. 28, 3) представляют собой как бы заранее расставленную и тонко продуманную ловушку, заставляющую насекомое произвести опыление. Крупная желтая губа, имеющая форму туфельки с завернутыми внутрь краями, сверху закрыта щитковидным стаминодием. Под ним лежит рыльце с обращенной вниз воспринимающей поверхностью, а по обе стороны от рыльца — по одной фертильной тычинке. Одиночные пчелы, особенно самки из рода андрена (Andraena), вползают и соскальзывают в губу цветка и, обследовав ее, начинают искать из нее выход. Ограниченное пространство не позволяет им взлететь, а завернутые края губы, ее гладкая внутренняя поверхность и заслон из стаминодия не позволяют не только выползти кратчайшим путем, но и рассмотреть прямой выход со дна губы. В задней стенке губы в направлении к гиностемию имеются, однако, «окна» — участки ткани, образованные клетками без пигментации, пропускающими свет. Принимая их за выходы, насекомое ползет в указанном ему направлении и отсюда уже видит настоящие выходы наружу: два узких отверстия слева и справа от колонки вплотную к пыльникам. Выбираясь из цветка, пчела сначала встречает на своем пути первую преграду — выпуклое рыльце. Грудь насекомого проходит над одним из выпуклых изгибов рыльца, которые счищают с нее принесенную пыльцу. Когда пчела. пробирается все выше и ближе к свету, вторая помеха — пыльник суживает проход. А когда несекомое протискивается мимо пыльника, большая порция липкой пыльцы, как подушечка, приклеивается к груди пчелы (рис. 147).Мелкие и ослабленные насекомые иногда так приклеиваются к пыльнику, что остаются там навсегда, закупоривая выход. А все, кому удается выбраться, уставшие и нередко мокрые после вынужденного купания в дождевой воде, которая скапливается на дне губы, на некоторое время даже теряют способность к координированному движению, но, отдохнув, снова летят на другие цветки. Что привлекает их в этих цветках? Дарвин считал, что насекомых может интересовать жидкость, выделяемая волосками, расположенными полосками в передней части губы. Е. Дауман (1969) показал, что, вопреки предположениям разных авторов, здесь не происходит отделения нектара или каких-либо иных питательных веществ, а маслянистая жидкость, содержащаяся в волосках, не служит питательным веществом для насекомых. Весь механизм опыления здесь построен на обмане: насекомых привлекает, по-видимому, запах и цвет, а яркий стаминодий служит для них указателем несуществующего нектара.
Опылителями орхидей подсемейства циприпедиевых являются двукрылые и пчелиные. Цветки одной из наиболее декоративных за пределами тропиков орхидей венерина башмачка (Cypripedium calceolus, рис. 147, табл. 28, 3) представляют собой как бы заранее расставленную и тонко продуманную ловушку, заставляющую насекомое произвести опыление. Крупная желтая губа, имеющая форму туфельки с завернутыми внутрь краями, сверху закрыта щитковидным стаминодием. Под ним лежит рыльце с обращенной вниз воспринимающей поверхностью, а по обе стороны от рыльца — по одной фертильной тычинке. Одиночные пчелы, особенно самки из рода андрена (Andraena), вползают и соскальзывают в губу цветка и, обследовав ее, начинают искать из нее выход. Ограниченное пространство не позволяет им взлететь, а завернутые края губы, ее гладкая внутренняя поверхность и заслон из стаминодия не позволяют не только выползти кратчайшим путем, но и рассмотреть прямой выход со дна губы. В задней стенке губы в направлении к гиностемию имеются, однако, «окна» — участки ткани, образованные клетками без пигментации, пропускающими свет. Принимая их за выходы, насекомое ползет в указанном ему направлении и отсюда уже видит настоящие выходы наружу: два узких отверстия слева и справа от колонки вплотную к пыльникам. Выбираясь из цветка, пчела сначала встречает на своем пути первую преграду — выпуклое рыльце. Грудь насекомого проходит над одним из выпуклых изгибов рыльца, которые счищают с нее принесенную пыльцу. Когда пчела. пробирается все выше и ближе к свету, вторая помеха — пыльник суживает проход. А когда несекомое протискивается мимо пыльника, большая порция липкой пыльцы, как подушечка, приклеивается к груди пчелы (рис. 147).Мелкие и ослабленные насекомые иногда так приклеиваются к пыльнику, что остаются там навсегда, закупоривая выход. А все, кому удается выбраться, уставшие и нередко мокрые после вынужденного купания в дождевой воде, которая скапливается на дне губы, на некоторое время даже теряют способность к координированному движению, но, отдохнув, снова летят на другие цветки. Что привлекает их в этих цветках? Дарвин считал, что насекомых может интересовать жидкость, выделяемая волосками, расположенными полосками в передней части губы. Е. Дауман (1969) показал, что, вопреки предположениям разных авторов, здесь не происходит отделения нектара или каких-либо иных питательных веществ, а маслянистая жидкость, содержащаяся в волосках, не служит питательным веществом для насекомых. Весь механизм опыления здесь построен на обмане: насекомых привлекает, по-видимому, запах и цвет, а яркий стаминодий служит для них указателем несуществующего нектара.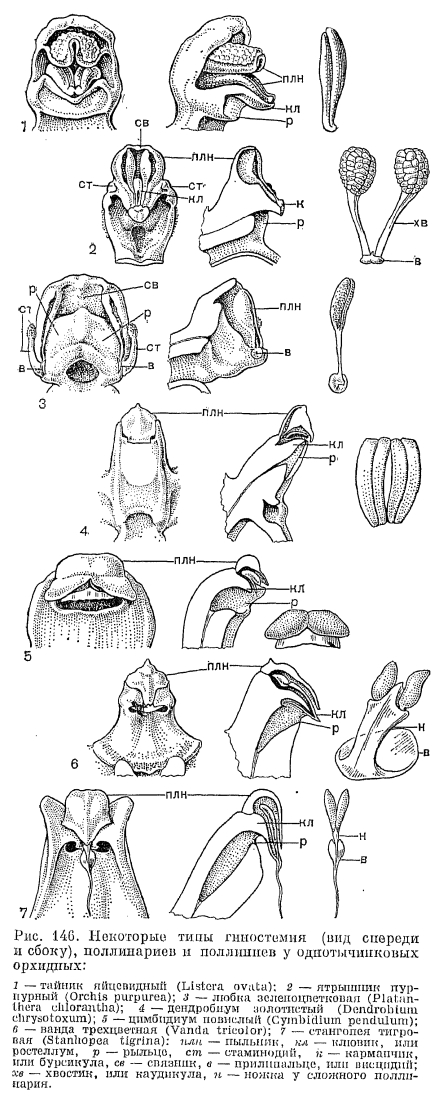
,
,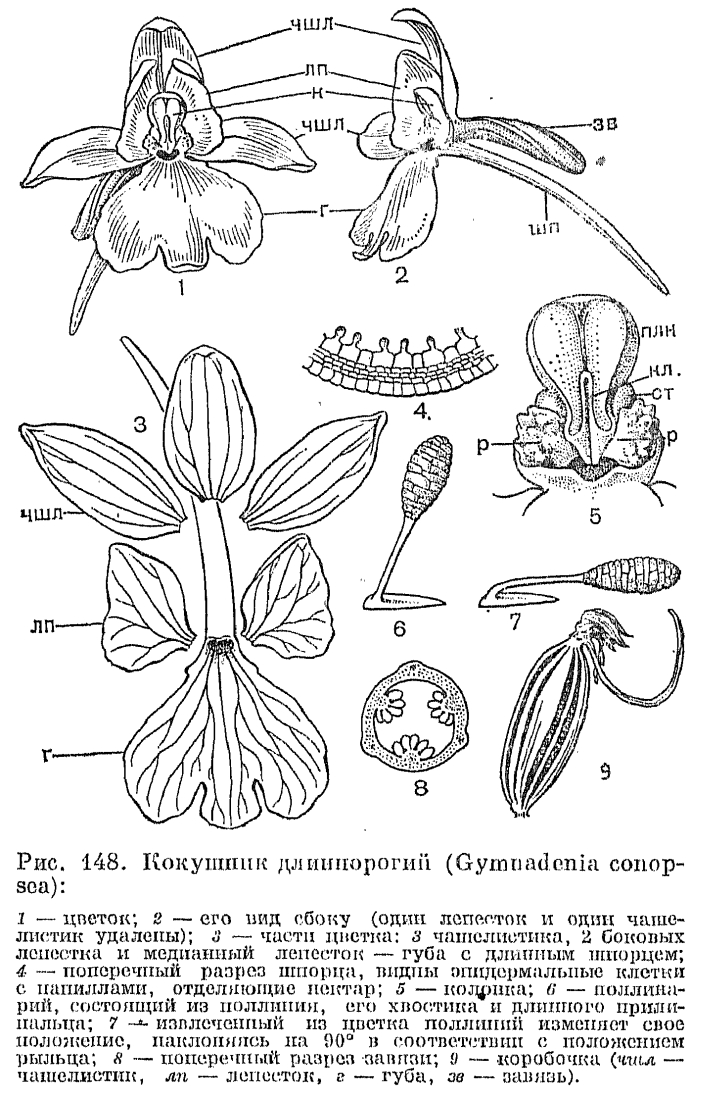 У подсемейства собственно орхидных бесчисленные вариации колонки определяют специфичность агентов опыления. На примере ятрышника мужского (Orchis mascula) — довольно обычной в Европе орхидеи — рассмотрим основную схему опыления у орхидных с базитонными пыльниками. Завязь этого вида ресупинирована, и губа занимает удобное для насекомых нижнее положение, имеет короткий шпорец. Два верхних лепестка и верхний чашелистик образуют колпачок, защищающий колонку от непогоды. В каждом пыльцевом мешке, открывающемся продольной щелью, располагается по одному поллинию с каудикулами и прилипальцами, скрытыми в карманчике на краю клювика (строение колонки ятрышника показано на рис. 146). Когда насекомое, севшее на губу, просовывает голову в полость, ведущую в шпорец, оно неизбежно натыкается на выступающий в этот проход клювик, нависающий над рыльцем. Конец клювика от прикосновения мгновенно расщепляется, и эластичный карманчик отгибается, обнажая 2 прилипальца, которые касаются головы насекомого, и быстро твердея, накрепко приклеиваются к нему. Если насекомое извлекает голову из цветка, оно вытаскивает за хвостик поллиний (один или оба), эластичный карманчик при этом захлопывается снова, защищая прилипальце оставшегося поллиния. Прикрепившиеся поллинарии сначала стоят вертикально на голове насекомого, как 2 рожка (рис. 147, 6 и рис. 148, 6). Но в таком положении при посещении следующего цветка они попали бы снова на гнезда пыльника. И здесь вступает в действие замечательная способность поллинария к движению. Во время полета (примерно через 30 с) поллинарий благодаря несимметричному высыханию хвостика и сокращению прилипальца изгибается вперед примерно под углом 90°, и при посещении пчелой другого цветка поллиний точно попадает на его рыльце. Поверхность рыльца липкая, и весь поллиний или отдельные пакетики пыльцы остаются на нем, а пчела улетает с остатками хвостиков на голове, как свидетельством успешно выполненной работы.Большинство ятрышников не имеет нектара в шпорце, и чем они привлекают насекомых, остается неизвестным. Ч. Дарвин полагал, что насекомые способны пробивать своим хоботком внутреннюю поверхность шпорца и высасывать обильную жидкость из его ткани, но более поздними наблюдениями это не было подтверждено. Ятрышник опыляют преимущественно короткохоботковые пчелиные, а также бабочки и мухи.У других орхидных с базитонными. пыльниками процесс опыления отличается только небольшими деталями. Например, у кокушника (рис. 148) удлиненные прилипальца образуют сводчатую кровлю над входом в нектарник — длинный шпорец, а лопасти рыльца занимают боковое положение с двух сторон от вхнда в нектарник. Когда бабочки просовывают хоботок в шпорец, сводчатые прилипальца приклеивается к хоботку по бокам его и, как и у ятрышника, сначала торчат вертикально, а затем наклоняются вперед по обе стороны хоботка и, точно соответствуя боковому расположению рылец, попадают на них при последующем посещении цветка. Цветки кокушника привлекают насекомых запахом, усиливающимся к вечеру, и предлагают им нектар, обильно выделяемый эпидермальными клетками с папиллами в длинном изогнутом шпорце.
У подсемейства собственно орхидных бесчисленные вариации колонки определяют специфичность агентов опыления. На примере ятрышника мужского (Orchis mascula) — довольно обычной в Европе орхидеи — рассмотрим основную схему опыления у орхидных с базитонными пыльниками. Завязь этого вида ресупинирована, и губа занимает удобное для насекомых нижнее положение, имеет короткий шпорец. Два верхних лепестка и верхний чашелистик образуют колпачок, защищающий колонку от непогоды. В каждом пыльцевом мешке, открывающемся продольной щелью, располагается по одному поллинию с каудикулами и прилипальцами, скрытыми в карманчике на краю клювика (строение колонки ятрышника показано на рис. 146). Когда насекомое, севшее на губу, просовывает голову в полость, ведущую в шпорец, оно неизбежно натыкается на выступающий в этот проход клювик, нависающий над рыльцем. Конец клювика от прикосновения мгновенно расщепляется, и эластичный карманчик отгибается, обнажая 2 прилипальца, которые касаются головы насекомого, и быстро твердея, накрепко приклеиваются к нему. Если насекомое извлекает голову из цветка, оно вытаскивает за хвостик поллиний (один или оба), эластичный карманчик при этом захлопывается снова, защищая прилипальце оставшегося поллиния. Прикрепившиеся поллинарии сначала стоят вертикально на голове насекомого, как 2 рожка (рис. 147, 6 и рис. 148, 6). Но в таком положении при посещении следующего цветка они попали бы снова на гнезда пыльника. И здесь вступает в действие замечательная способность поллинария к движению. Во время полета (примерно через 30 с) поллинарий благодаря несимметричному высыханию хвостика и сокращению прилипальца изгибается вперед примерно под углом 90°, и при посещении пчелой другого цветка поллиний точно попадает на его рыльце. Поверхность рыльца липкая, и весь поллиний или отдельные пакетики пыльцы остаются на нем, а пчела улетает с остатками хвостиков на голове, как свидетельством успешно выполненной работы.Большинство ятрышников не имеет нектара в шпорце, и чем они привлекают насекомых, остается неизвестным. Ч. Дарвин полагал, что насекомые способны пробивать своим хоботком внутреннюю поверхность шпорца и высасывать обильную жидкость из его ткани, но более поздними наблюдениями это не было подтверждено. Ятрышник опыляют преимущественно короткохоботковые пчелиные, а также бабочки и мухи.У других орхидных с базитонными. пыльниками процесс опыления отличается только небольшими деталями. Например, у кокушника (рис. 148) удлиненные прилипальца образуют сводчатую кровлю над входом в нектарник — длинный шпорец, а лопасти рыльца занимают боковое положение с двух сторон от вхнда в нектарник. Когда бабочки просовывают хоботок в шпорец, сводчатые прилипальца приклеивается к хоботку по бокам его и, как и у ятрышника, сначала торчат вертикально, а затем наклоняются вперед по обе стороны хоботка и, точно соответствуя боковому расположению рылец, попадают на них при последующем посещении цветка. Цветки кокушника привлекают насекомых запахом, усиливающимся к вечеру, и предлагают им нектар, обильно выделяемый эпидермальными клетками с папиллами в длинном изогнутом шпорце. Опыление любки двулистной производят ночные бабочки, особенно бражники, летящие на сильный запах, который испускают к вечеру белые цветки. Отделенные друг от друга и расположенные по бокам клювика прилипальца (рис. 140) приклеиваются сбоку головки бабочки или к основанию хоботка либо даже на один из ее глаз, и во время полета их хвостик совершает движение вниз к центру так, чтобы поллиний мог попасть затем на поверхность рыльца, расположенную в середине колонки. А у близкого к ятрышнику рода анакамптис (Аnаcamptis) прилипальце имеет седловидную форму, и во время полета бабочки края этого «седла» мгновенно закручиваются внутрь, охватывая хоботок с двух сторон, что приводит к расхождению поллиниев в стороны, затем они изгибаются, как у кокушника, параллельно хоботку, и такое их положение соответствует боковому положению лопастей рыльца. Разнообразные выросты, бороздки на губе ориентируют опылителя в нужном направлении. У анакамптиса, например, на губе имеются суживающие валики, направляющие движение хоботка насекомого к клювику. Иногда и сам клювик определяет действие насекомых. У хабенарии прямо вперед торчащий клювик заставляет пчел просовывать хоботок в нектарник со стороны так, чтобы хоботок коснулся рыльца. А у спирантеса (Spiranthes) клювик в молодых цветках закрывает вход в нектарник и одновременно доступ к рыльцу. Насекомое может только удалить поллинии, но не оставить их на рыльце, но у более старых цветков клювик сморщивается и отодвигается. Тогда освобождается проход к рыльцу. Этим обесценивается перекрестное опыление.Среди орхидных трибы ятрышниковых наиболее замечательный способ опыления наблюдается у офрисов. Цветки этих красивых и редких растений, растущих в Европе, Западной Азии и Северной Африке, часто напоминают насекомых (мух, пчел, шмелей) или пауков. От этого сходства и происходят их видовые названия — офрис насекомоносный (Ophrys insectifera), офрис пауковидный (О. spfegodes), офрис шмелецветковый (О. bombyliflora) и т. д.
Опыление любки двулистной производят ночные бабочки, особенно бражники, летящие на сильный запах, который испускают к вечеру белые цветки. Отделенные друг от друга и расположенные по бокам клювика прилипальца (рис. 140) приклеиваются сбоку головки бабочки или к основанию хоботка либо даже на один из ее глаз, и во время полета их хвостик совершает движение вниз к центру так, чтобы поллиний мог попасть затем на поверхность рыльца, расположенную в середине колонки. А у близкого к ятрышнику рода анакамптис (Аnаcamptis) прилипальце имеет седловидную форму, и во время полета бабочки края этого «седла» мгновенно закручиваются внутрь, охватывая хоботок с двух сторон, что приводит к расхождению поллиниев в стороны, затем они изгибаются, как у кокушника, параллельно хоботку, и такое их положение соответствует боковому положению лопастей рыльца. Разнообразные выросты, бороздки на губе ориентируют опылителя в нужном направлении. У анакамптиса, например, на губе имеются суживающие валики, направляющие движение хоботка насекомого к клювику. Иногда и сам клювик определяет действие насекомых. У хабенарии прямо вперед торчащий клювик заставляет пчел просовывать хоботок в нектарник со стороны так, чтобы хоботок коснулся рыльца. А у спирантеса (Spiranthes) клювик в молодых цветках закрывает вход в нектарник и одновременно доступ к рыльцу. Насекомое может только удалить поллинии, но не оставить их на рыльце, но у более старых цветков клювик сморщивается и отодвигается. Тогда освобождается проход к рыльцу. Этим обесценивается перекрестное опыление.Среди орхидных трибы ятрышниковых наиболее замечательный способ опыления наблюдается у офрисов. Цветки этих красивых и редких растений, растущих в Европе, Западной Азии и Северной Африке, часто напоминают насекомых (мух, пчел, шмелей) или пауков. От этого сходства и происходят их видовые названия — офрис насекомоносный (Ophrys insectifera), офрис пауковидный (О. spfegodes), офрис шмелецветковый (О. bombyliflora) и т. д.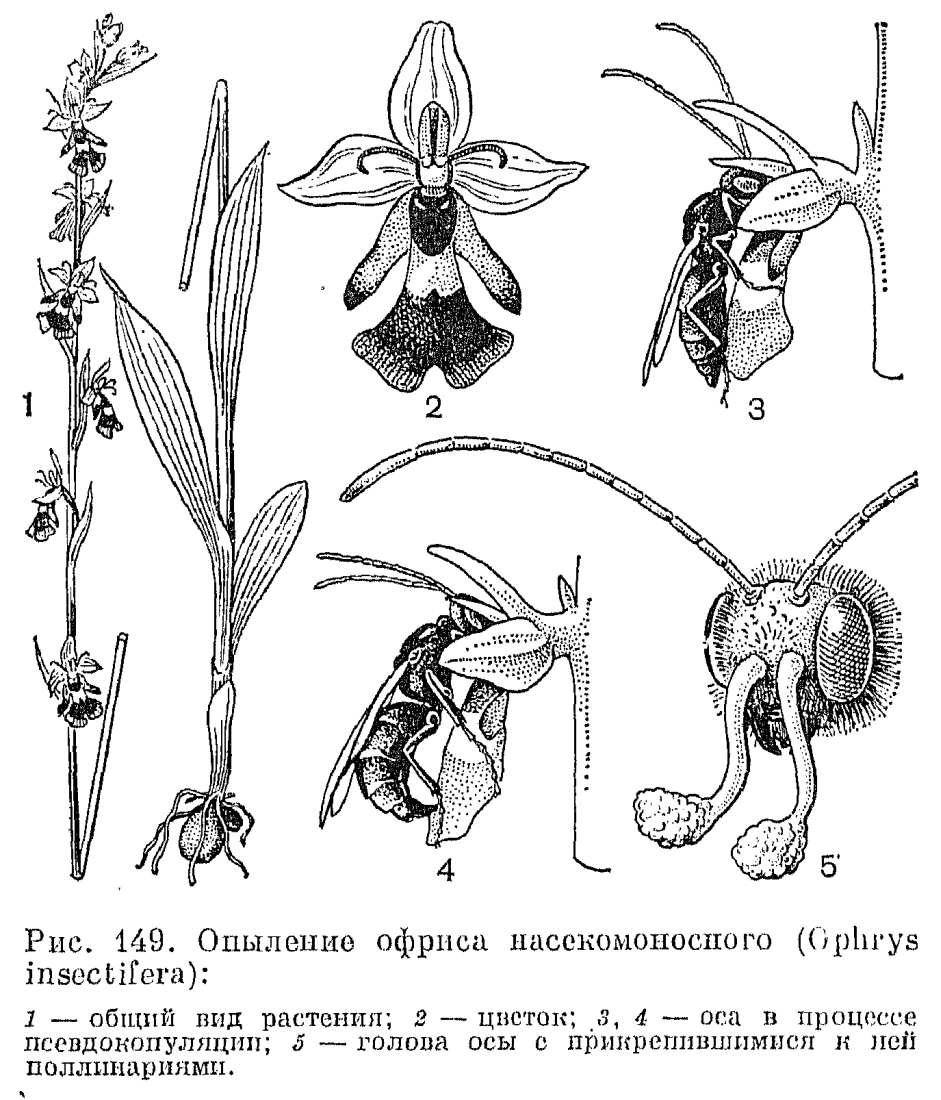 У цветков офриса насекомоносного (рис. 149), произрастающего на карбонатной почве на лугах и лесных опушках от Средиземноморья и Малой Азии до Скандинавии, губа похожа на тело насекомого со сложенными крыльями, блеск которых имитируют серо-голубые полоски на губе, боковые лепестки имеют вид антенн, а расположенные в основании губы ложные нектарники напоминают глаза насекомого. Это загадочное сходство привлекало внимание многих ученых. Каким образом оно возникло и зачем оно растениям? Какие только догадки не высказывались по этому доводу! Не «сфотографировались» ли насекомые, кружащиеся над офрисами, на их цветках? Не существует ли это сходство только в восприятии человека, а насекомые его не замечают? Загадка разрешилась после того, как в 1916 г. французский ученый М. Пуйян высказал сенсационное предположение: цветки офриса воздействуют на половые инстинкты насекомых-самцов, имитируя их самок. Гипотеза Пуйяна была принята многими учеными. В России она пропагандировалась Б. М. Козо-Полянским (1938, 1939) в его блестящих работах о мимикрии у растений, в которых он также высказал предположение, что в привлечении самцов насекомых имеют значение какие-то неуловимые для нас запахи. Все это позднее было подтверждено экспериментально, а совсем недавно также наблюдениями в природе шведских ученых Б. Кулленберга и Г. Бергстрема (1976). Б. Кулленберг снял документальный фильм об опылении этих удивительных цветков, который был показан на XII Международном ботаническом конгрессе в Ленинграде в 1975 г.Цветки офрисов не «предлагают» насекомым нектара, пыльцы или иных питательных веществ. Но губа их выделяет летучие вещества из группы феромонов, главным образом производные жирных кислот и циклические сесквитерпены, набор которых специфичен для каждого вида или группы видов. Некоторые из этих соединений идентичны секрету желёз насекомых и воздействуют на них как ключевой раздражитель, стимулирующий цепь поведенческих реакций, характерных для спаривания. Внешний облик губы действует как визуальный стимул, а различные эпидермальные выросты на ней и определенным образом расположенные волоски воздействуют как тактильный раздражитель при попытках спаривания. Самцы жалящих перепончатокрылых из родов андрена (Andrena), эвцера (Еuсеrа), кампсосколия (Campsoscolia), антофора (Anthophora) и др. появляются после перезимовки несколько раньше самок и, побуждаемые половым инстинктом, летят на цветки офрисов. На разных их видах насекомые, в зависимости от структуры губы, ориентируются либо головой, либо брюшком к гиностемию и ведут себя так, как при копуляции с самками (рис. 149). В результате их импульсивных движений поллинии прилипают либо к их голове, либо к брюшку. Сексуальный инстинкт насекомых не удовлетворяется на одном цветке, и в результате нескольких посещений они производят перекрестное опыление.Такая приспособленность офрисов к специфическим опылителям могла возникнуть только в результате длительной синхронной эволюции насекомых и растений. В этой цепи развития приспособлений наиболее специализирован офрис зеркальный (Ophrys speculum), опыляемый только осой из семейства сколиидовых кампсосколией реснитчатой (Campsoscolia. ciliata), а наименее — офрис пчелоносный (О. apifera), который частично опыляется пчелами рода эвцера, а частично самоопыляется. При этом в строении цветка его имеются и совершенные приспособления к самоопылению. Они состоят в том, что гнезда пыльников самопроизвольно вскрываются, и поллинии под влиянием собственной тяжести выпадают из них, опускаются в направлении рыльцевой ямки, оставаясь прикрепленными к фиксированным хвостикам, имеющим как раз такую длину, которая необходима, чтобы при колебаниях воздуха они коснулись рыльца.Процент завязывания семян у опыляющихся описанным способом офрисов чрезвычайно низок вследствие их очень узкой специализации по отношению к определенным видам насекомых. Особенно это заметно по контрасту с самоопыляющимся офрисом пчелоносным, у которого, как отмечено Ч. Дарвином, каждый цветок дает коробочку с семенами.Привлечение опылителей запахами, воздействующими на их сексуальные инстинкты, характерно и для ряда орхидных из других триб, в частности для видов спикулеи (Spiculaea), дракеи (Drakaea), криптостилиса (Cryptostylus). Губа этих растений, с точки зрения человека, не всегда имеет сходство с самкой насекомого, тем не менее самцы пытаются спариваться с ней и уносят поллинии, которые, изменив во время полета свое положение на наклонное, при следующем «спаривании» точно попадают на рыльце.Рассмотрим теперь опыление нескольких представителей трибы неоттиевых — листеры, или тайника, неоттии, или гнездовки, и эпипактиса, или дремлика. Принципиальное отличие их от представителей трибы орхидных состоит в том, что поллинии их лишены каудикул и контактируют с клювиком своей верхушкой (рис. 140, 7). У листеры в начале цветения клювик, как ширма нависающий над рыльцем, не имеет связи с поллиниями. Внутри клювик подразделен на ряд камер, содержащих липкую жидкость. Пыльник вскрывается еще в бутоне, и поллинии свободно лежат на клювике, защищенные вытянутой верхушкой тычиночной нити. Удлиненная раздвоенная на конце губа имеет по центру продольную бороздку, в которой скапливается нектар. Мелкие двукрылые и перепончатокрылые садятся на губу и, слизывая нектар, постепенно двигаются снизу вверх. Заканчивая трапезу, они поднимают головку вверх и касаются клювика. Из него тотчас выступает капля клейкой жидкости, которая приклеивается одновременно и к головке насекомого, и к поллиниям. Через несколько секунд эта жидкость затвердевает, прочно цементируя поллиний на голове насекомого. Поскольку поллинии из-за отсутствия хвостиков не способны здесь к движению, это компенсируется движением клювика и явлением протандрии. У более старых цветков, из которых извлечены поллинии, клювик поднимается кверху, освобождая широкий доступ к рыльцу. И множество разнообразных насекомых, для которых невзрачные цветки листеры с открытым нектаром очень привлекательны, успешно совершают перекрестное опыление.Сходный механизм перекрестного опыления характерен для гнездовки, но пыльцевые массы здесь гораздо более рассыпчаты, и пыльца часто самопроизвольно падает на рыльце или переносится мелкими ползающими насекомыми в пределах одного цветка. Это вызывает самоопыление, которое является здесь, по-видимому, нормой. У неоттии бывают и клейстогамные цветки, развивающиеся под землей или в моховом покрове и образующие нормальные семена. Есть сообщения о клейстогамии и у других неоттиевых.
У цветков офриса насекомоносного (рис. 149), произрастающего на карбонатной почве на лугах и лесных опушках от Средиземноморья и Малой Азии до Скандинавии, губа похожа на тело насекомого со сложенными крыльями, блеск которых имитируют серо-голубые полоски на губе, боковые лепестки имеют вид антенн, а расположенные в основании губы ложные нектарники напоминают глаза насекомого. Это загадочное сходство привлекало внимание многих ученых. Каким образом оно возникло и зачем оно растениям? Какие только догадки не высказывались по этому доводу! Не «сфотографировались» ли насекомые, кружащиеся над офрисами, на их цветках? Не существует ли это сходство только в восприятии человека, а насекомые его не замечают? Загадка разрешилась после того, как в 1916 г. французский ученый М. Пуйян высказал сенсационное предположение: цветки офриса воздействуют на половые инстинкты насекомых-самцов, имитируя их самок. Гипотеза Пуйяна была принята многими учеными. В России она пропагандировалась Б. М. Козо-Полянским (1938, 1939) в его блестящих работах о мимикрии у растений, в которых он также высказал предположение, что в привлечении самцов насекомых имеют значение какие-то неуловимые для нас запахи. Все это позднее было подтверждено экспериментально, а совсем недавно также наблюдениями в природе шведских ученых Б. Кулленберга и Г. Бергстрема (1976). Б. Кулленберг снял документальный фильм об опылении этих удивительных цветков, который был показан на XII Международном ботаническом конгрессе в Ленинграде в 1975 г.Цветки офрисов не «предлагают» насекомым нектара, пыльцы или иных питательных веществ. Но губа их выделяет летучие вещества из группы феромонов, главным образом производные жирных кислот и циклические сесквитерпены, набор которых специфичен для каждого вида или группы видов. Некоторые из этих соединений идентичны секрету желёз насекомых и воздействуют на них как ключевой раздражитель, стимулирующий цепь поведенческих реакций, характерных для спаривания. Внешний облик губы действует как визуальный стимул, а различные эпидермальные выросты на ней и определенным образом расположенные волоски воздействуют как тактильный раздражитель при попытках спаривания. Самцы жалящих перепончатокрылых из родов андрена (Andrena), эвцера (Еuсеrа), кампсосколия (Campsoscolia), антофора (Anthophora) и др. появляются после перезимовки несколько раньше самок и, побуждаемые половым инстинктом, летят на цветки офрисов. На разных их видах насекомые, в зависимости от структуры губы, ориентируются либо головой, либо брюшком к гиностемию и ведут себя так, как при копуляции с самками (рис. 149). В результате их импульсивных движений поллинии прилипают либо к их голове, либо к брюшку. Сексуальный инстинкт насекомых не удовлетворяется на одном цветке, и в результате нескольких посещений они производят перекрестное опыление.Такая приспособленность офрисов к специфическим опылителям могла возникнуть только в результате длительной синхронной эволюции насекомых и растений. В этой цепи развития приспособлений наиболее специализирован офрис зеркальный (Ophrys speculum), опыляемый только осой из семейства сколиидовых кампсосколией реснитчатой (Campsoscolia. ciliata), а наименее — офрис пчелоносный (О. apifera), который частично опыляется пчелами рода эвцера, а частично самоопыляется. При этом в строении цветка его имеются и совершенные приспособления к самоопылению. Они состоят в том, что гнезда пыльников самопроизвольно вскрываются, и поллинии под влиянием собственной тяжести выпадают из них, опускаются в направлении рыльцевой ямки, оставаясь прикрепленными к фиксированным хвостикам, имеющим как раз такую длину, которая необходима, чтобы при колебаниях воздуха они коснулись рыльца.Процент завязывания семян у опыляющихся описанным способом офрисов чрезвычайно низок вследствие их очень узкой специализации по отношению к определенным видам насекомых. Особенно это заметно по контрасту с самоопыляющимся офрисом пчелоносным, у которого, как отмечено Ч. Дарвином, каждый цветок дает коробочку с семенами.Привлечение опылителей запахами, воздействующими на их сексуальные инстинкты, характерно и для ряда орхидных из других триб, в частности для видов спикулеи (Spiculaea), дракеи (Drakaea), криптостилиса (Cryptostylus). Губа этих растений, с точки зрения человека, не всегда имеет сходство с самкой насекомого, тем не менее самцы пытаются спариваться с ней и уносят поллинии, которые, изменив во время полета свое положение на наклонное, при следующем «спаривании» точно попадают на рыльце.Рассмотрим теперь опыление нескольких представителей трибы неоттиевых — листеры, или тайника, неоттии, или гнездовки, и эпипактиса, или дремлика. Принципиальное отличие их от представителей трибы орхидных состоит в том, что поллинии их лишены каудикул и контактируют с клювиком своей верхушкой (рис. 140, 7). У листеры в начале цветения клювик, как ширма нависающий над рыльцем, не имеет связи с поллиниями. Внутри клювик подразделен на ряд камер, содержащих липкую жидкость. Пыльник вскрывается еще в бутоне, и поллинии свободно лежат на клювике, защищенные вытянутой верхушкой тычиночной нити. Удлиненная раздвоенная на конце губа имеет по центру продольную бороздку, в которой скапливается нектар. Мелкие двукрылые и перепончатокрылые садятся на губу и, слизывая нектар, постепенно двигаются снизу вверх. Заканчивая трапезу, они поднимают головку вверх и касаются клювика. Из него тотчас выступает капля клейкой жидкости, которая приклеивается одновременно и к головке насекомого, и к поллиниям. Через несколько секунд эта жидкость затвердевает, прочно цементируя поллиний на голове насекомого. Поскольку поллинии из-за отсутствия хвостиков не способны здесь к движению, это компенсируется движением клювика и явлением протандрии. У более старых цветков, из которых извлечены поллинии, клювик поднимается кверху, освобождая широкий доступ к рыльцу. И множество разнообразных насекомых, для которых невзрачные цветки листеры с открытым нектаром очень привлекательны, успешно совершают перекрестное опыление.Сходный механизм перекрестного опыления характерен для гнездовки, но пыльцевые массы здесь гораздо более рассыпчаты, и пыльца часто самопроизвольно падает на рыльце или переносится мелкими ползающими насекомыми в пределах одного цветка. Это вызывает самоопыление, которое является здесь, по-видимому, нормой. У неоттии бывают и клейстогамные цветки, развивающиеся под землей или в моховом покрове и образующие нормальные семена. Есть сообщения о клейстогамии и у других неоттиевых.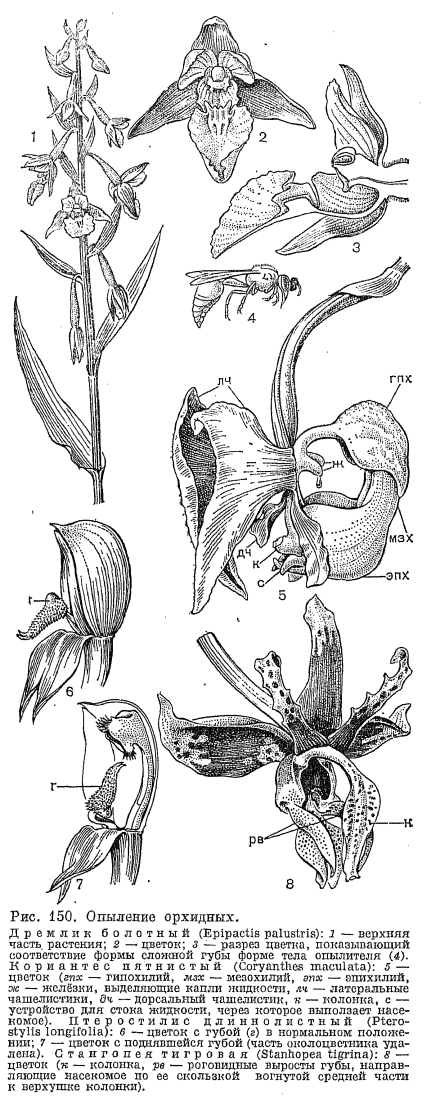 Опыление дремлика болотного (Epipactis раlustris, рис. 150) производят главным образом одиночные осы и, как выяснено Л. Нильсоном (1978), строение и размеры частей губы являются идеальной адаптацией к строению и размерам их тела. Губа дремлика разделена на 2 подвижно сочлененные половины: дистальную (эпихилий) и базальную (гипохилий). Эпихилий у своего основания утолщен, снабжен посредине глубокой бороздкой и краями обращен вверх, причем расстояние между краями очень мало (1,7 мм), так что там может разместиться только мелкое насекомое, у которого верхний сегмент брюшка меньше этой величины. Из жалящих перепончатокрылых со стебельчатым брюшком наиболее подходящие размеры имеют осы рода эвменес (Eumenes), которые и являются самыми эффективными опылителями этих растений (по наблюдениям Л. Нильсона в Швеции). Первый сегмент их брюшка имеет своеобразную колокольчатую форму, идеально соответствующую величине и форме эпихилия. Когда оса (рис. 150) садится на эпихилий, он, будучи подвижно сочленен с гипохилием, прогибается под ее тяжестью вниз и при кратковременной потере равновесия оса делает инстинктивное балансирующее движение своим брюшком, приводящее ее в контакт с клювиком и поллиниями. Нектар дремлика, отделяемый в чашевидной части губы, обладает наркотическим действием, что, возможно, зависит от сбраживающих процессов, вызываемых в нем дрожжевыми грибками, которые там обнаружены. Оса после посещения одного цветка еще способна сбросить с себя поллинии своими сильными ножками, но после посещения нескольких цветков она, отравленная нектаром, падает на землю и уже не может лететь и переползает от цветка к цветку, от соцветия, к соцветию, нагруженная гроздьями поллиниев. А цветки дремлика долго не увядают и после опыления, продолжая выделять дурманящий нектар, неудержимо влекущий к ним насекомых во все время цветения. Кроме эвменеса, цветки дремлика опыляют и другие насекомые, другие одиночные осы, шмели, муравьи.
Опыление дремлика болотного (Epipactis раlustris, рис. 150) производят главным образом одиночные осы и, как выяснено Л. Нильсоном (1978), строение и размеры частей губы являются идеальной адаптацией к строению и размерам их тела. Губа дремлика разделена на 2 подвижно сочлененные половины: дистальную (эпихилий) и базальную (гипохилий). Эпихилий у своего основания утолщен, снабжен посредине глубокой бороздкой и краями обращен вверх, причем расстояние между краями очень мало (1,7 мм), так что там может разместиться только мелкое насекомое, у которого верхний сегмент брюшка меньше этой величины. Из жалящих перепончатокрылых со стебельчатым брюшком наиболее подходящие размеры имеют осы рода эвменес (Eumenes), которые и являются самыми эффективными опылителями этих растений (по наблюдениям Л. Нильсона в Швеции). Первый сегмент их брюшка имеет своеобразную колокольчатую форму, идеально соответствующую величине и форме эпихилия. Когда оса (рис. 150) садится на эпихилий, он, будучи подвижно сочленен с гипохилием, прогибается под ее тяжестью вниз и при кратковременной потере равновесия оса делает инстинктивное балансирующее движение своим брюшком, приводящее ее в контакт с клювиком и поллиниями. Нектар дремлика, отделяемый в чашевидной части губы, обладает наркотическим действием, что, возможно, зависит от сбраживающих процессов, вызываемых в нем дрожжевыми грибками, которые там обнаружены. Оса после посещения одного цветка еще способна сбросить с себя поллинии своими сильными ножками, но после посещения нескольких цветков она, отравленная нектаром, падает на землю и уже не может лететь и переползает от цветка к цветку, от соцветия, к соцветию, нагруженная гроздьями поллиниев. А цветки дремлика долго не увядают и после опыления, продолжая выделять дурманящий нектар, неудержимо влекущий к ним насекомых во все время цветения. Кроме эвменеса, цветки дремлика опыляют и другие насекомые, другие одиночные осы, шмели, муравьи.
,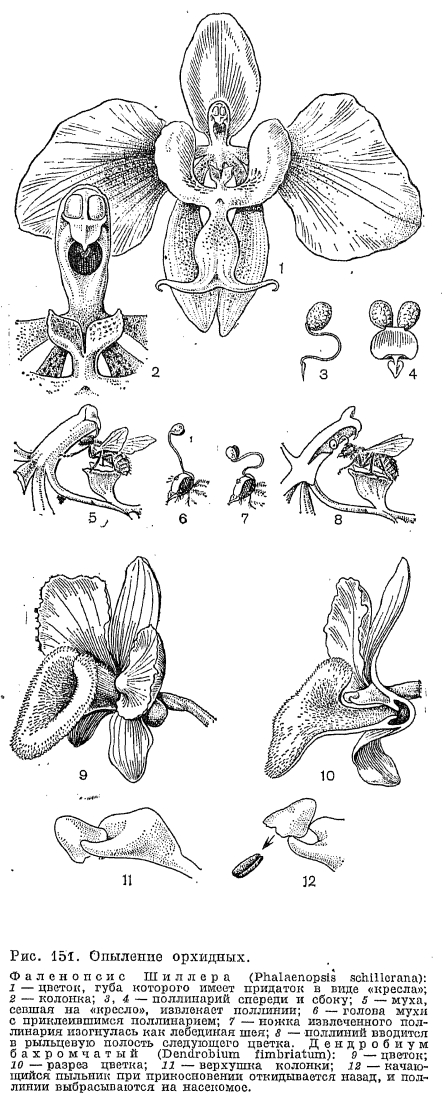 Характерное строение колонки эпидендровых и вандовых (рис. 146) с пыльником, расположенным на ней сверху, как шапочка, обусловливает особенности опыления этих растений. Поллинии приклеиваются к насекомым в момент, когда опылители покидают цветок и при этом приподнимают крышку пыльника. Пыльник, если он опадающий, приклеивается к насекомому вместе с поллиниями, а если он качающийся (сидит на тонкой нити с подвижным сочленением), то при прикосновении он отскакивает назад, выбрасывая поллинии (рис. 151). Сохранение крышки пыльника, снимающейся вместе с поллиниями, препятствует опылению того же цветка и, кроме того, действует как раздражающий фактор для насекомого, заставляющий его взлететь, чтобы избавиться от помехи. Сбросив с себя пыльник, насекомое с прикрепившимися поллиниями летит на следующий цветок, производя перекрестное опыление. У дендробиума золотистоцветкового (Dendrobium chrysanthum), по наблюдению Ч. Дарвина, после посещения насекомого включается еще дополнительный механизм, гарантирующий самоопыление. Поллинии здесь, если не успели приклеиться к опылителю после скачкообразного движения пыльника, провисают на эластичной нити до самой губы венчика. Губа под действием толчка от взлетающего насекомого подбрасывает поллиний так, что он попадает на рыльце собственного цветка, где и прикрепляется.В опылении вандовых большое значение имеет характерная для их поллиниев ножка, которая выполняет здесь ту же роль, что и каудикула в трибе ятрышниковых. Ножка вследствие высыхания на открытом воздухе и сокращения способна к движению, может свертываться, наклоняться, складываться, благодаря чему поллинии занимают положение, необходимое для точного попадания на рыльце; длина ее в сложенном виде обычно соответствует глубине рыльцевой полости. От движения ножки происходит также сбрасывание с поллиниев крышечки пыльника. Ножки вандовых бывают очень разной формы. Каудикулы, как правило, скрыты в поллиниях, тем не менее они прочно прикрепляют их к ножке и благодаря их прочности поллинии могут быть удержаны и оторваны только очень липким рыльцем, что соответствует состоянию его зрелости. Тем самым также обеспечивается перекрестное опыление. Насекомых привлекает нектар, выделяемый в глубоком нектарнике, расположенном на внутренней стенке рыльцевой полости ниже рыльца или в шпорце губы. Опыление производят крупные насекомые, нередко пчелиные и бабочки, вынужденные глубоко просовывать в узкую рыльцевую щель свои хоботки, чтобы добыть нектар. Во многих случаях эта щель так узка, что способ введения в нее поллиниев остается не вполне ясным.Успеху перекрестного опыления нередко способствует особая структура губы. У прекрасной орхидеи азиатских тропиков фаленопсиса (Phalaenopsis) губа снабжена у своего основания разной формы придатками. У фаленопсиса Шиллера (P. schillerana) этот придаток имеет форму кресла (рис. 151, 1 — 5), на котором удобно устраивается муха, просовывающая свою голову в круглое отверстие рыльцевой полости, где имеется нектар. Прямо над этим отверстием нависает клювик, действительно имеющий здесь форму птичьего клюва, верхушка которого легко прилипает к голове мухи. Когда муха закончит свой обед, она вытягивает из пыльника оба поллиния, соединенные с прилипальцем клювика, и летит с ними на другой цветок. За краткое время перелета ножка поллиниев изгибается, по выражению А. Кернера фон Марилауна (1891), подобно лебединой шее, и в другом цветке направленные вперед поллинии входят в рыльцевую полость и приклеиваются к рыльцевой поверхности. У некоторых других видов фаленопсиса придатки губы захлопываются как щипцы вокруг насекомого, заставляя его извлечь поллинии вместе с пыльпиками.
Характерное строение колонки эпидендровых и вандовых (рис. 146) с пыльником, расположенным на ней сверху, как шапочка, обусловливает особенности опыления этих растений. Поллинии приклеиваются к насекомым в момент, когда опылители покидают цветок и при этом приподнимают крышку пыльника. Пыльник, если он опадающий, приклеивается к насекомому вместе с поллиниями, а если он качающийся (сидит на тонкой нити с подвижным сочленением), то при прикосновении он отскакивает назад, выбрасывая поллинии (рис. 151). Сохранение крышки пыльника, снимающейся вместе с поллиниями, препятствует опылению того же цветка и, кроме того, действует как раздражающий фактор для насекомого, заставляющий его взлететь, чтобы избавиться от помехи. Сбросив с себя пыльник, насекомое с прикрепившимися поллиниями летит на следующий цветок, производя перекрестное опыление. У дендробиума золотистоцветкового (Dendrobium chrysanthum), по наблюдению Ч. Дарвина, после посещения насекомого включается еще дополнительный механизм, гарантирующий самоопыление. Поллинии здесь, если не успели приклеиться к опылителю после скачкообразного движения пыльника, провисают на эластичной нити до самой губы венчика. Губа под действием толчка от взлетающего насекомого подбрасывает поллиний так, что он попадает на рыльце собственного цветка, где и прикрепляется.В опылении вандовых большое значение имеет характерная для их поллиниев ножка, которая выполняет здесь ту же роль, что и каудикула в трибе ятрышниковых. Ножка вследствие высыхания на открытом воздухе и сокращения способна к движению, может свертываться, наклоняться, складываться, благодаря чему поллинии занимают положение, необходимое для точного попадания на рыльце; длина ее в сложенном виде обычно соответствует глубине рыльцевой полости. От движения ножки происходит также сбрасывание с поллиниев крышечки пыльника. Ножки вандовых бывают очень разной формы. Каудикулы, как правило, скрыты в поллиниях, тем не менее они прочно прикрепляют их к ножке и благодаря их прочности поллинии могут быть удержаны и оторваны только очень липким рыльцем, что соответствует состоянию его зрелости. Тем самым также обеспечивается перекрестное опыление. Насекомых привлекает нектар, выделяемый в глубоком нектарнике, расположенном на внутренней стенке рыльцевой полости ниже рыльца или в шпорце губы. Опыление производят крупные насекомые, нередко пчелиные и бабочки, вынужденные глубоко просовывать в узкую рыльцевую щель свои хоботки, чтобы добыть нектар. Во многих случаях эта щель так узка, что способ введения в нее поллиниев остается не вполне ясным.Успеху перекрестного опыления нередко способствует особая структура губы. У прекрасной орхидеи азиатских тропиков фаленопсиса (Phalaenopsis) губа снабжена у своего основания разной формы придатками. У фаленопсиса Шиллера (P. schillerana) этот придаток имеет форму кресла (рис. 151, 1 — 5), на котором удобно устраивается муха, просовывающая свою голову в круглое отверстие рыльцевой полости, где имеется нектар. Прямо над этим отверстием нависает клювик, действительно имеющий здесь форму птичьего клюва, верхушка которого легко прилипает к голове мухи. Когда муха закончит свой обед, она вытягивает из пыльника оба поллиния, соединенные с прилипальцем клювика, и летит с ними на другой цветок. За краткое время перелета ножка поллиниев изгибается, по выражению А. Кернера фон Марилауна (1891), подобно лебединой шее, и в другом цветке направленные вперед поллинии входят в рыльцевую полость и приклеиваются к рыльцевой поверхности. У некоторых других видов фаленопсиса придатки губы захлопываются как щипцы вокруг насекомого, заставляя его извлечь поллинии вместе с пыльпиками.
,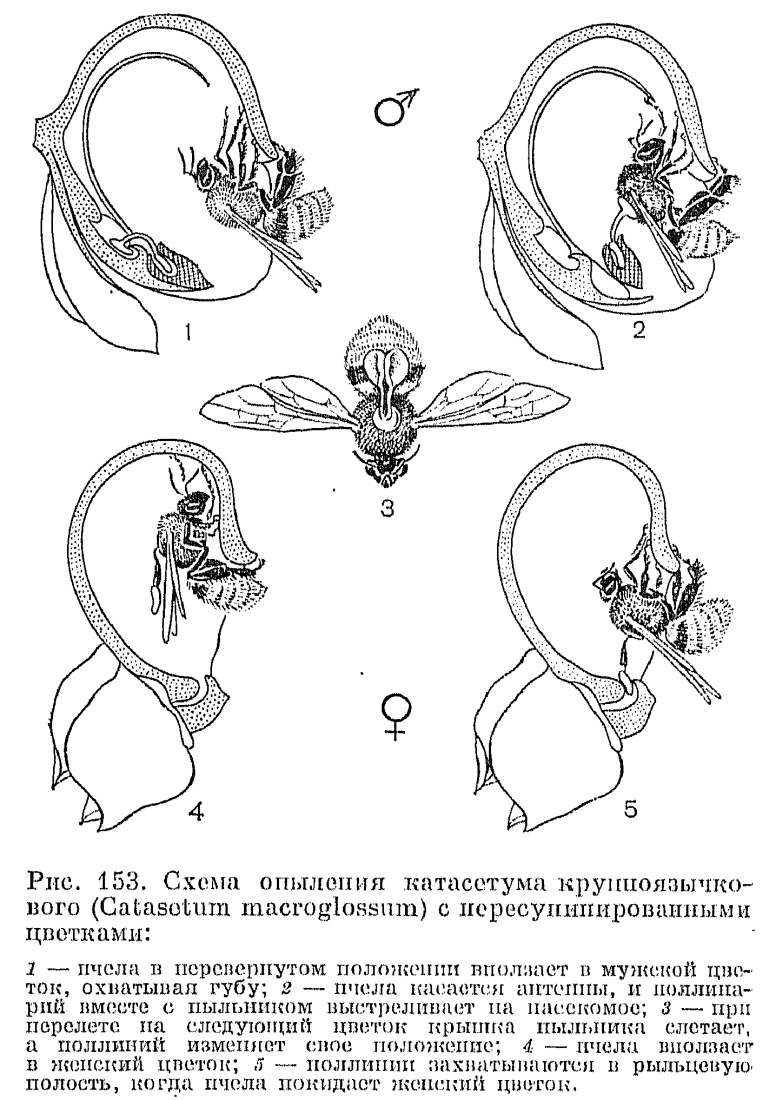 Но наиболее интересный механизм опыления среди вандовых имеет, пожалуй, представитель неотропической подтрибы катасетовых (Catasetinae) род катасетум (Catasetum, рис. 152). Для видов этого рода характерны однополые цветки, причем мужские и женские резко различаются по внешнему облику. Иногда образуются еще и обоеполые цветки, причем все три варианта могут встречаться в одном соцветии. Собранные отдельно растения с мужскими и женскими цветками раньше принимали за особые роды, пока не выяснилось, что одно и то же растение может производить женские цветки в один сезон, мужские — в другой и смешанное соцветие — в третий. Факторы, вызывающие такое чередование, недостаточно изучены. По мнению К. Додеона (1962), изучавшего опыление этой группы орхидных, это обусловлено главным образом изменением экологических условий. В мужских цветках видов катасетума колонка имеет длинные роговидные выросты-антенны, отходящие от клювика по обеим сторонам пыльника к центру цветка или к полости губы. Раздражимые антенны при прикосновении к ним насекомого срабатывают как спусковой механизм, освобождая натянутую как тетива ножку поллинария, который при этом выстреливает вперед прилипальцем на грудь насекомого. Опыление катасетума крупноязычкового (С. macroglossum) представлено на схеме (рис. 153, из К. Додеона, 1962). Цветки его с губой, занимающей верхнее положение, начинают испускать сильный мускусный запах через 2 — 3 дня после расцветания. Этот запах неудержимо влечет самцов пчел из рода эвлема (Eulaema). Они охватывают губу, вползают в цветок в перевернутом положении и начинают передними ножками скрести внутреннюю поверхность губы. Выделяющийся при этом пахучий секрет воздействует на их хеморецепторы, расположенные на подушечках передних ножек. Во время этой работы пчела слегка задевает кончик антенны, расположенной как раз над углублением в губе, наиболее привлекательным местом, откуда исходит самый сильный запах. И спущенные стрелы — поллинарии вместе с крышечкой пыльника прикрепляются к ее тораксу. Клейкое вещество прилипалец вмиг затвердевает, поллинарии слегка отклоняются вниз и назад вдоль средней линии брюшка, и в таком положении пчела вносит их в женский, также пересупинированный цветок, где ведет себя так же, как в мужском. В женском цветке провисающие в направлении рыльцевой щели поллинарии захватываются липкой рыльцевой поверхностью при выходе пчелы из цветка. Женские цветки катасетума испускают сильный запах до тех пор, пока не будут опылены, иногда в течение месяца, но через несколько часов после опыления запах исчезает.Другие виды подтрибы катасетовых, включающей еще 2 рода — мормодес (Mormodes) и цикнохес (Cycnoches), также способны выбрасывать поллинарий на опылителя, отличаясь деталями строения спускового механизма и способами прикрепления поллинариев. Опыление производят исключительно самцы пчел подсемейства эвглоссовых (Euglossinae), особенно из родов эвлема и эвглосса. Прежние сообщения о том, что пчелы обгрызают различные выросты на губе, не подтверждаются современными наблюдениями. Цветки не отделяют и нектара, так что единственным привлекающим фактором являются вещества, выделяемые осмофорами на губе растений. Эти пахучие вещества оказывают на насекомых наркотическое действие, пчелы совершенно теряют осторожность и обнаруживают острое пристрастие к «наркотику», по нескольку раз возвращаясь в один и тот же цветок. По некоторым предположениям (С. Фогель, 1966), самцы пчел, собирающиеся в скопления, используют этот цветочный запах, чтобы привлекать самок. Такие же «наркотические» вещества, сходным образом воздействующие на самцов пчел, выделяют и некоторые другие орхидеи из вандовых, в частности эпифитный род стангопея, цветок одного из видов которого — стангопеи тигровой (Stanhopea tigrina) — изображен на рисунке 150, 8 и на таблице 38, 1.
Но наиболее интересный механизм опыления среди вандовых имеет, пожалуй, представитель неотропической подтрибы катасетовых (Catasetinae) род катасетум (Catasetum, рис. 152). Для видов этого рода характерны однополые цветки, причем мужские и женские резко различаются по внешнему облику. Иногда образуются еще и обоеполые цветки, причем все три варианта могут встречаться в одном соцветии. Собранные отдельно растения с мужскими и женскими цветками раньше принимали за особые роды, пока не выяснилось, что одно и то же растение может производить женские цветки в один сезон, мужские — в другой и смешанное соцветие — в третий. Факторы, вызывающие такое чередование, недостаточно изучены. По мнению К. Додеона (1962), изучавшего опыление этой группы орхидных, это обусловлено главным образом изменением экологических условий. В мужских цветках видов катасетума колонка имеет длинные роговидные выросты-антенны, отходящие от клювика по обеим сторонам пыльника к центру цветка или к полости губы. Раздражимые антенны при прикосновении к ним насекомого срабатывают как спусковой механизм, освобождая натянутую как тетива ножку поллинария, который при этом выстреливает вперед прилипальцем на грудь насекомого. Опыление катасетума крупноязычкового (С. macroglossum) представлено на схеме (рис. 153, из К. Додеона, 1962). Цветки его с губой, занимающей верхнее положение, начинают испускать сильный мускусный запах через 2 — 3 дня после расцветания. Этот запах неудержимо влечет самцов пчел из рода эвлема (Eulaema). Они охватывают губу, вползают в цветок в перевернутом положении и начинают передними ножками скрести внутреннюю поверхность губы. Выделяющийся при этом пахучий секрет воздействует на их хеморецепторы, расположенные на подушечках передних ножек. Во время этой работы пчела слегка задевает кончик антенны, расположенной как раз над углублением в губе, наиболее привлекательным местом, откуда исходит самый сильный запах. И спущенные стрелы — поллинарии вместе с крышечкой пыльника прикрепляются к ее тораксу. Клейкое вещество прилипалец вмиг затвердевает, поллинарии слегка отклоняются вниз и назад вдоль средней линии брюшка, и в таком положении пчела вносит их в женский, также пересупинированный цветок, где ведет себя так же, как в мужском. В женском цветке провисающие в направлении рыльцевой щели поллинарии захватываются липкой рыльцевой поверхностью при выходе пчелы из цветка. Женские цветки катасетума испускают сильный запах до тех пор, пока не будут опылены, иногда в течение месяца, но через несколько часов после опыления запах исчезает.Другие виды подтрибы катасетовых, включающей еще 2 рода — мормодес (Mormodes) и цикнохес (Cycnoches), также способны выбрасывать поллинарий на опылителя, отличаясь деталями строения спускового механизма и способами прикрепления поллинариев. Опыление производят исключительно самцы пчел подсемейства эвглоссовых (Euglossinae), особенно из родов эвлема и эвглосса. Прежние сообщения о том, что пчелы обгрызают различные выросты на губе, не подтверждаются современными наблюдениями. Цветки не отделяют и нектара, так что единственным привлекающим фактором являются вещества, выделяемые осмофорами на губе растений. Эти пахучие вещества оказывают на насекомых наркотическое действие, пчелы совершенно теряют осторожность и обнаруживают острое пристрастие к «наркотику», по нескольку раз возвращаясь в один и тот же цветок. По некоторым предположениям (С. Фогель, 1966), самцы пчел, собирающиеся в скопления, используют этот цветочный запах, чтобы привлекать самок. Такие же «наркотические» вещества, сходным образом воздействующие на самцов пчел, выделяют и некоторые другие орхидеи из вандовых, в частности эпифитный род стангопея, цветок одного из видов которого — стангопеи тигровой (Stanhopea tigrina) — изображен на рисунке 150, 8 и на таблице 38, 1.
, Очень крупные, пятнистые, сильно пахнущие цветки этой орхидеи парами свисают с ветвей деревьев. Занимающие самое нижнее положение в цветке губа и колонка располагаются напротив друг друга. Губа отчетливо разделена на 3 части (эпихилий, мезохилий, гипохилий), из них средняя часть имеет 2 крупных, направленных к колонке выроста, похожих на рога. «Голова быка» — называют это растение на его родине в Мексике. Привлекаемые запахом самцы эвглоссы скребут передними ножками углубленную поверхность верхней части губы, «пьянеют» и сползают вниз по гладкой, как каток, поверхности, «рога» же точно направляют их к колонке. При этом эвглосса задевает острие клювика со стороны прилипальца и уносит с собой поллинии.Опишем еще несколько хитроумных приспособлений к опылению у орхидных. Кориантес (Coryanthos), замечательный род американских орхидей, часто поселяется на гнездах древесных муравьев. Его крупные, у некоторых видов достигающие в диаметре 13 см, цветки парами свисают вниз на длинных, ножках. Замечателен этот род оригинальной формой своей губы, состоящей, как и у стангопеи, из трех частей (рис. 150, 5). Гипохилий имеет форму ножки, отклоняющейся под прямым углом от оси цветка и от основания колонки и расширенной пузыревидно в месте перехода в цилиндрический мезохилий. От мезохилия отходит горизонтально и параллельно «ножке» (гипохилию) нижняя часть губы (эпихилий), которая и определяет все «события» опыления. Эпихилий имеет форму ванночки с очень гладкими стенками. Когда цветок полностью открыт, в «ванночку» стекает капельками бесцветная жидкость, выделяемая двумя желёзками, расположенными у основания колонки. Избыток жидкости вытекает через «сточную трубочку» — трехрогий отросток, действующий как носик у чайника. Колонка свисает вниз, причем ее изогнутая верхушка располагается как раз над этой трубочкой. Цветение продолжается 3 — 4 дня, чашелистики увядают раньше, делая губу еще более заметной насекомым. Привлекаемые сильным запахом, самцы эвглоссовых садятся на губу и, как и в предыдущих случаях, скребут поверхность гипохилия. «Пьянея», а также в результате борьбы друг с другом за место у «вкусного стола» они сваливаются в приготовленную для них ванну. Приняв «холодную ванну», они начинают барахтаться в поисках выхода и выползают через «сточную трубочку», тесно примыкающую к колонке, поллинии при этом приклеиваются к насекомому. По наблюдениям П. Аллена (1950), для того чтобы выбраться из ловушки, пчеле требуется около 45 мин. После удаления поллиниев цветок перестает издавать запах, но на следующее утро запах появляется вновь и снова привлекает насекомых, чем достигается перекрестное опыление.Отметим еще несколько способов опыления, связанных с подвижностью губы и цветков. У многих орхидных части губы подвижно сочленены друг с другом или губа соединена эластичным сочленением с основанием или выростом (ножкой) колонки и совершает колебательные движения на ветру. Мелкие невзрачные цветки бульбофиллума бородконосного (Bulbophyllum barbigerum) из Западной Африки в результате дрожания губы и булавовидных волосков на ее верхушке становятся похожими на маленьких трепещущих насекомых и привлекают опылителей-мух. А у цирропеталума украшенного (Cirrhopetalum ornatissimum) эффект аналогичного колебания цветков еще усиливается благодаря постоянному мерцательному движению ресничек на их боковых лепестках.Дрожание цветков видов южноамериканского рода онцидиум (Oncidium) предназначено для использования в опылении сложных непищевых инстинктов насекомых. Самцы некоторых перепончатокрылых, например рода центрис (Centris), покидают гнезда и сидят в засаде, охраняя свою территорию от вторжения пришельцев. И когда желто-коричневые пятнистые цветки онцидиума начинают при порыве ветра дрожать, насекомые принимают их за вторгшихся представителей другого вида, подвергают внезапной атаке с целью изгнания соперников со своей территории и производят опыление.В ряде других случаев подвижно сочлененная губа действует как механическое приспособление для приведения насекомого в контакт с колонкой. У американской наземной орхидеи калопогона красивейшего (Calopogon pulchellus), цветки которого напоминают стайку расположившихся на стебле розовато-лиловых бабочек, губа обращена вверх и заканчивается желтой бородкой. Пчелы летят на яркую бородку, цепляются за нее, и под действием тяжести пчелы губа, обладающая подвижным сочленением, прогибается вниз, приводя спинку пчелы в контакт с колонкой. А у многих видов орхидных с перевернутыми цветками губа способна благодаря своей раздражимости или смещению центра тяжести к внезапному движению вверх в направлении колонки, опрокидывающему насекомое спиной на рыльце и пыльник. Такое замечательное устройство имеют многие виды бульбофиллума, виды плакоглоттиса (Placoglottis), чувствительной подвижной губой известны австралийские представители трибы неоттиевых калеана (Caleana), дракея (Drakaea), спикулея (Spicullaea), птеростилис (Pterostylis). Цветок рода птеростилис с губой в двух положениях изображен на рисунке 150, 6, 7. Два лепестка и один из чашелистиков этого цветка образуют колпачок над колонкой, и, когда взметнувшаяся вверх губа закроет вход в этот колпачок, насекомое оказывается заключенным во временной тюрьме, из которой оно выпускается после того, как губа снова откинется и станет опять чувствительной к прикосновению. За это время пытающееся освободиться насекомое успевает извлечь поллиний или произвести опыление.Приведенные примеры не дают, конечно, сколько-нибудь полного представления о бесконечно разнообразных и удивительных способах опыления орхидных. Интересующегося читателя мы отсылаем к увлекательной, выше цитированной книге Дарвина, многочисленным работам К. Додсона, Л. ван дер Пейла, Л. Нильсона, Р. Бейтса, М. Проктора и др.Рассмотрим лишь еще некоторые общие аспекты опыления орхидных. Среди множества приспособлений к перекрестному опылению следует отметить одно, универсальное для семейства, — длительность цветения. Только у некоторых родов (стангопея, собралия, некоторые виды дендробиума) цветки остаются свежими 1 — 5 суток. У большинства орхидных цветки в ожидании опыления не увядают неделями и месяцами, до 2 — 3 месяцев у ванды, одонтоглоссума, онцидиума. У многих орхидных цветки в соцветии распускаются неодновременно и все соцветие имеет свежий вид еще более длительный срок. Иногда цветение его растягивается на целый вегетационный период, а новые ветви могут образоваться и на следующий год. В большинстве случаев соцветия выносятся далеко в пространство, для того чтобы они были видны и удобны для посещения летающим насекомым, но иногда, наоборот, ось соцветия и цветоносы сильно укорачиваются и соцветия прижимаются к субстрату, для того чтобы опыление могли производить ползающие насекомые. Пыльца у большинства представителей семейства находится в связанном состоянии и не может разноситься ветром. Да и рыльца не приспособлены для улавливания пыльцы из воздуха. Орхидные поэтому опыляются только животными (насекомыми и птицами) или самоопыляются. Как установлено К. Додсоном (1966), основную часть (50%) опылителей семейства составляют перепончатокрылые (Hymenoptera), особенно те из них, для которых пищей является нектар. На втором месте (18%) — чешуекрылые (Lepidopterа), особенно ночные бабочки, на третьем (12%) — двукрылые (Diptera). Колибри как опылители составляют всего 3%, установлено опыление ими красных цветков дизы (Disa), масдеваллии (Masdevallia), эллеантуса (Elleanthus) и др. Некоторые крупные роды орхидных, например эпидендрум (Epidendrum), могут опыляться разными опылителями, но многие другие роды адаптированы к опылению преимущественно одной группой насекомых. Так, большинство видов цимбидиума и дендробиума опыляют пчелы, многие виды масдеваллии, бульбофиллума с губой, часто превращенной в ловчий аппарат, опыляют мясные мухи. Любку, ангрекум, хабенарию опыляют ночные бабочки, которые способны парить в воздухе и не нуждаются в посадочной площадке (губа у растений этих родов утратила свою функцию и либо обращена вверх, либо рассечена).Бесконечно разнообразные форма и окраска околоцветника, различные вспомогательные устройства способствуют успеху перекрестного опыления орхидных. Они предлагают красные тона птицам, все оттенки красок — пчелам, грязно-фиолетовые или коричневато-красные — мясным мухам, снежно-белые — ночным бабочкам. А запахи их варьируют от запаха гниющего мяса до тончайшего аромата духов, исходящего часто от невзрачных и некрупных зеленовато-белых цветков.Пыльца как пищевой фактор не имеет значения в опылении орхидных. Она экономично упакована в поллинии, не тратится впустую и у большинства видов вся целиком попадает на рыльце, что способствует оплодотворению сотен тысяч семязачатков, которые находятся в каждой завязи и из которых разовьются сотни тысяч семян. Основной «товар», который предлагают насекомым растения с помощью яркой «рекламы», — это нектар. Иногда он открытый и выделяется в бороздках губы, у основания колонки, или он скапливается в глубине рыльцевой полости или в шпорцах, отходящих от основания губы, иногда от чашелистиков. У некоторых орхидных нектарники осевые, они пронизывают на разную глубину стенки завязи и иногда пузыревидно вздуты. Разная длина шпорцев соответствует разной длине хоботков опылителей. Поразительной длины достигают шпорцы у некоторых видов ангрекума, аэридеса (Aerides), аэрангиса (Aerangis). У ангрекума полуторафутового (Angraecum sesquipedale) их длина составляет 21 — 30 см! Долго оставалось загадкой, какое же насекомое может достать нектар, скапливающийся на дне этого шпорца. И хотя Ч. Дарвин еще в 1877 г. предсказал существование гигантской бабочки с очень длинным хоботком, открыта она была на Мадагаскаре только в 1903 г. Бабочка ксантопан Моргана-предсказанная (Xanthopan morgani-praedicta) имеет свернутый в спираль, хоботок длиной 22, 5 см. Эта или подобные ей бабочки вполне могут достать нектар из длинного шпорца ангрекума.Иногда насекомые отгрызают волоски и разнообразные выросты на губе растения. Известны случаи собирания ими воска с губы орнитидиума (Ornithidium). У эрии (Eria), полистахии (Polystachia) и максиллярии (Maxillaria) на губе образуется пыльцевидный порошок, поедаемый насекомыми. Но не всегда они находят в цветках то, что ищут: шпорцы оказываются пустыми, желтые, похожие на наполненный пыльцой пыльник пятна на губе — окрашенными волосками, а заманчивые резервуары с жидкостью — хитрыми ловушками, из которых не выбраться без «принудительной работы» по опылению. Обман — это еще один и весьма эффективный способ привлечения насекомых к опылению в этой группе растений. По наиболее замечательным приспособлением можно считать использование ими в опылении непищевых инстинктов насекомых (сексуальных, защитных) и широкое привлечение к опылению самцов пчел и ос, т.е. группы насекомых, обычно не участвующей в опылении растений. Ф. Г. Бригер (1971), Р. Л. Дресслер (1981) объясняют такую своеобразную адаптацию тем, что семейство орхидных вступило на арену эволюции, когда уже существовало множество конкурентов, снабжавших опылителей пищей — нектаром и пыльцой, и орхидеи вынуждены были искать другие пути обеспечения опыления.Казалось бы, при таком совершенстве приспособлений к опылению результатом должна быть 100-процентная завязываемость семян. На самом деле это сверхсовершенство оборачивается обратной стороной. Именно в силу очень узкой специализации цветки орхидных часто остаются неопыленными. Ч. Дарвин приводит следующие цифры и наблюдения: в тропических лесах Южной Бразилии у эпидендрума на 370 цветков образовалась только одна коробочка, 223 цветка опали неопыленными, а из 146 оставшихся только у 4 были удалены поллинии. Очень редко образует коробочки обильно цветущая в Бразилии ваниль, не более чем 1 цветок из 1000 дает коробочку у дендробиума красивого (Dendrobium speciosum) в Новом Южном Уэльсе. Только 5 коробочек на 200 цветков завязалось у кориантеса трехлопастного (Coryanthes triloba). Не очень плодовиты и орхидные в Европе. Многие из них прибегают к самоопылению для поддержания своего существования. Не являются ли в таком случае все достижения длительного приспособительного развития орхидных по отношению к определенным насекомым своего рода «ошибкой эволюции»? Ответом на этот вопрос служит тот факт, что орхидные — одно из двух крупнейших семейств цветковых растений, освоившее почти все экологические ниши на Земле. Ограниченные в силу узкой специализации возможности опыления компенсируются у них невероятно высоким количеством семян, которое образуется в каждой коробочке. Ч. Дарвин подсчитал, что если бы все семена пальчатокоренника пятнистого (Dactylorhiza maculata, табл. 33, 3) благополучно прорастали, то правнуки одного растения «почти могли бы покрыть однообразным зеленым ковром всю поверхность суши на земном шаре». И того небольшого количества семян, которые превращаются во взрослые растения, орхидным вполне достаточно для продолжения рода в ненарушенных природных условиях.
Очень крупные, пятнистые, сильно пахнущие цветки этой орхидеи парами свисают с ветвей деревьев. Занимающие самое нижнее положение в цветке губа и колонка располагаются напротив друг друга. Губа отчетливо разделена на 3 части (эпихилий, мезохилий, гипохилий), из них средняя часть имеет 2 крупных, направленных к колонке выроста, похожих на рога. «Голова быка» — называют это растение на его родине в Мексике. Привлекаемые запахом самцы эвглоссы скребут передними ножками углубленную поверхность верхней части губы, «пьянеют» и сползают вниз по гладкой, как каток, поверхности, «рога» же точно направляют их к колонке. При этом эвглосса задевает острие клювика со стороны прилипальца и уносит с собой поллинии.Опишем еще несколько хитроумных приспособлений к опылению у орхидных. Кориантес (Coryanthos), замечательный род американских орхидей, часто поселяется на гнездах древесных муравьев. Его крупные, у некоторых видов достигающие в диаметре 13 см, цветки парами свисают вниз на длинных, ножках. Замечателен этот род оригинальной формой своей губы, состоящей, как и у стангопеи, из трех частей (рис. 150, 5). Гипохилий имеет форму ножки, отклоняющейся под прямым углом от оси цветка и от основания колонки и расширенной пузыревидно в месте перехода в цилиндрический мезохилий. От мезохилия отходит горизонтально и параллельно «ножке» (гипохилию) нижняя часть губы (эпихилий), которая и определяет все «события» опыления. Эпихилий имеет форму ванночки с очень гладкими стенками. Когда цветок полностью открыт, в «ванночку» стекает капельками бесцветная жидкость, выделяемая двумя желёзками, расположенными у основания колонки. Избыток жидкости вытекает через «сточную трубочку» — трехрогий отросток, действующий как носик у чайника. Колонка свисает вниз, причем ее изогнутая верхушка располагается как раз над этой трубочкой. Цветение продолжается 3 — 4 дня, чашелистики увядают раньше, делая губу еще более заметной насекомым. Привлекаемые сильным запахом, самцы эвглоссовых садятся на губу и, как и в предыдущих случаях, скребут поверхность гипохилия. «Пьянея», а также в результате борьбы друг с другом за место у «вкусного стола» они сваливаются в приготовленную для них ванну. Приняв «холодную ванну», они начинают барахтаться в поисках выхода и выползают через «сточную трубочку», тесно примыкающую к колонке, поллинии при этом приклеиваются к насекомому. По наблюдениям П. Аллена (1950), для того чтобы выбраться из ловушки, пчеле требуется около 45 мин. После удаления поллиниев цветок перестает издавать запах, но на следующее утро запах появляется вновь и снова привлекает насекомых, чем достигается перекрестное опыление.Отметим еще несколько способов опыления, связанных с подвижностью губы и цветков. У многих орхидных части губы подвижно сочленены друг с другом или губа соединена эластичным сочленением с основанием или выростом (ножкой) колонки и совершает колебательные движения на ветру. Мелкие невзрачные цветки бульбофиллума бородконосного (Bulbophyllum barbigerum) из Западной Африки в результате дрожания губы и булавовидных волосков на ее верхушке становятся похожими на маленьких трепещущих насекомых и привлекают опылителей-мух. А у цирропеталума украшенного (Cirrhopetalum ornatissimum) эффект аналогичного колебания цветков еще усиливается благодаря постоянному мерцательному движению ресничек на их боковых лепестках.Дрожание цветков видов южноамериканского рода онцидиум (Oncidium) предназначено для использования в опылении сложных непищевых инстинктов насекомых. Самцы некоторых перепончатокрылых, например рода центрис (Centris), покидают гнезда и сидят в засаде, охраняя свою территорию от вторжения пришельцев. И когда желто-коричневые пятнистые цветки онцидиума начинают при порыве ветра дрожать, насекомые принимают их за вторгшихся представителей другого вида, подвергают внезапной атаке с целью изгнания соперников со своей территории и производят опыление.В ряде других случаев подвижно сочлененная губа действует как механическое приспособление для приведения насекомого в контакт с колонкой. У американской наземной орхидеи калопогона красивейшего (Calopogon pulchellus), цветки которого напоминают стайку расположившихся на стебле розовато-лиловых бабочек, губа обращена вверх и заканчивается желтой бородкой. Пчелы летят на яркую бородку, цепляются за нее, и под действием тяжести пчелы губа, обладающая подвижным сочленением, прогибается вниз, приводя спинку пчелы в контакт с колонкой. А у многих видов орхидных с перевернутыми цветками губа способна благодаря своей раздражимости или смещению центра тяжести к внезапному движению вверх в направлении колонки, опрокидывающему насекомое спиной на рыльце и пыльник. Такое замечательное устройство имеют многие виды бульбофиллума, виды плакоглоттиса (Placoglottis), чувствительной подвижной губой известны австралийские представители трибы неоттиевых калеана (Caleana), дракея (Drakaea), спикулея (Spicullaea), птеростилис (Pterostylis). Цветок рода птеростилис с губой в двух положениях изображен на рисунке 150, 6, 7. Два лепестка и один из чашелистиков этого цветка образуют колпачок над колонкой, и, когда взметнувшаяся вверх губа закроет вход в этот колпачок, насекомое оказывается заключенным во временной тюрьме, из которой оно выпускается после того, как губа снова откинется и станет опять чувствительной к прикосновению. За это время пытающееся освободиться насекомое успевает извлечь поллиний или произвести опыление.Приведенные примеры не дают, конечно, сколько-нибудь полного представления о бесконечно разнообразных и удивительных способах опыления орхидных. Интересующегося читателя мы отсылаем к увлекательной, выше цитированной книге Дарвина, многочисленным работам К. Додсона, Л. ван дер Пейла, Л. Нильсона, Р. Бейтса, М. Проктора и др.Рассмотрим лишь еще некоторые общие аспекты опыления орхидных. Среди множества приспособлений к перекрестному опылению следует отметить одно, универсальное для семейства, — длительность цветения. Только у некоторых родов (стангопея, собралия, некоторые виды дендробиума) цветки остаются свежими 1 — 5 суток. У большинства орхидных цветки в ожидании опыления не увядают неделями и месяцами, до 2 — 3 месяцев у ванды, одонтоглоссума, онцидиума. У многих орхидных цветки в соцветии распускаются неодновременно и все соцветие имеет свежий вид еще более длительный срок. Иногда цветение его растягивается на целый вегетационный период, а новые ветви могут образоваться и на следующий год. В большинстве случаев соцветия выносятся далеко в пространство, для того чтобы они были видны и удобны для посещения летающим насекомым, но иногда, наоборот, ось соцветия и цветоносы сильно укорачиваются и соцветия прижимаются к субстрату, для того чтобы опыление могли производить ползающие насекомые. Пыльца у большинства представителей семейства находится в связанном состоянии и не может разноситься ветром. Да и рыльца не приспособлены для улавливания пыльцы из воздуха. Орхидные поэтому опыляются только животными (насекомыми и птицами) или самоопыляются. Как установлено К. Додсоном (1966), основную часть (50%) опылителей семейства составляют перепончатокрылые (Hymenoptera), особенно те из них, для которых пищей является нектар. На втором месте (18%) — чешуекрылые (Lepidopterа), особенно ночные бабочки, на третьем (12%) — двукрылые (Diptera). Колибри как опылители составляют всего 3%, установлено опыление ими красных цветков дизы (Disa), масдеваллии (Masdevallia), эллеантуса (Elleanthus) и др. Некоторые крупные роды орхидных, например эпидендрум (Epidendrum), могут опыляться разными опылителями, но многие другие роды адаптированы к опылению преимущественно одной группой насекомых. Так, большинство видов цимбидиума и дендробиума опыляют пчелы, многие виды масдеваллии, бульбофиллума с губой, часто превращенной в ловчий аппарат, опыляют мясные мухи. Любку, ангрекум, хабенарию опыляют ночные бабочки, которые способны парить в воздухе и не нуждаются в посадочной площадке (губа у растений этих родов утратила свою функцию и либо обращена вверх, либо рассечена).Бесконечно разнообразные форма и окраска околоцветника, различные вспомогательные устройства способствуют успеху перекрестного опыления орхидных. Они предлагают красные тона птицам, все оттенки красок — пчелам, грязно-фиолетовые или коричневато-красные — мясным мухам, снежно-белые — ночным бабочкам. А запахи их варьируют от запаха гниющего мяса до тончайшего аромата духов, исходящего часто от невзрачных и некрупных зеленовато-белых цветков.Пыльца как пищевой фактор не имеет значения в опылении орхидных. Она экономично упакована в поллинии, не тратится впустую и у большинства видов вся целиком попадает на рыльце, что способствует оплодотворению сотен тысяч семязачатков, которые находятся в каждой завязи и из которых разовьются сотни тысяч семян. Основной «товар», который предлагают насекомым растения с помощью яркой «рекламы», — это нектар. Иногда он открытый и выделяется в бороздках губы, у основания колонки, или он скапливается в глубине рыльцевой полости или в шпорцах, отходящих от основания губы, иногда от чашелистиков. У некоторых орхидных нектарники осевые, они пронизывают на разную глубину стенки завязи и иногда пузыревидно вздуты. Разная длина шпорцев соответствует разной длине хоботков опылителей. Поразительной длины достигают шпорцы у некоторых видов ангрекума, аэридеса (Aerides), аэрангиса (Aerangis). У ангрекума полуторафутового (Angraecum sesquipedale) их длина составляет 21 — 30 см! Долго оставалось загадкой, какое же насекомое может достать нектар, скапливающийся на дне этого шпорца. И хотя Ч. Дарвин еще в 1877 г. предсказал существование гигантской бабочки с очень длинным хоботком, открыта она была на Мадагаскаре только в 1903 г. Бабочка ксантопан Моргана-предсказанная (Xanthopan morgani-praedicta) имеет свернутый в спираль, хоботок длиной 22, 5 см. Эта или подобные ей бабочки вполне могут достать нектар из длинного шпорца ангрекума.Иногда насекомые отгрызают волоски и разнообразные выросты на губе растения. Известны случаи собирания ими воска с губы орнитидиума (Ornithidium). У эрии (Eria), полистахии (Polystachia) и максиллярии (Maxillaria) на губе образуется пыльцевидный порошок, поедаемый насекомыми. Но не всегда они находят в цветках то, что ищут: шпорцы оказываются пустыми, желтые, похожие на наполненный пыльцой пыльник пятна на губе — окрашенными волосками, а заманчивые резервуары с жидкостью — хитрыми ловушками, из которых не выбраться без «принудительной работы» по опылению. Обман — это еще один и весьма эффективный способ привлечения насекомых к опылению в этой группе растений. По наиболее замечательным приспособлением можно считать использование ими в опылении непищевых инстинктов насекомых (сексуальных, защитных) и широкое привлечение к опылению самцов пчел и ос, т.е. группы насекомых, обычно не участвующей в опылении растений. Ф. Г. Бригер (1971), Р. Л. Дресслер (1981) объясняют такую своеобразную адаптацию тем, что семейство орхидных вступило на арену эволюции, когда уже существовало множество конкурентов, снабжавших опылителей пищей — нектаром и пыльцой, и орхидеи вынуждены были искать другие пути обеспечения опыления.Казалось бы, при таком совершенстве приспособлений к опылению результатом должна быть 100-процентная завязываемость семян. На самом деле это сверхсовершенство оборачивается обратной стороной. Именно в силу очень узкой специализации цветки орхидных часто остаются неопыленными. Ч. Дарвин приводит следующие цифры и наблюдения: в тропических лесах Южной Бразилии у эпидендрума на 370 цветков образовалась только одна коробочка, 223 цветка опали неопыленными, а из 146 оставшихся только у 4 были удалены поллинии. Очень редко образует коробочки обильно цветущая в Бразилии ваниль, не более чем 1 цветок из 1000 дает коробочку у дендробиума красивого (Dendrobium speciosum) в Новом Южном Уэльсе. Только 5 коробочек на 200 цветков завязалось у кориантеса трехлопастного (Coryanthes triloba). Не очень плодовиты и орхидные в Европе. Многие из них прибегают к самоопылению для поддержания своего существования. Не являются ли в таком случае все достижения длительного приспособительного развития орхидных по отношению к определенным насекомым своего рода «ошибкой эволюции»? Ответом на этот вопрос служит тот факт, что орхидные — одно из двух крупнейших семейств цветковых растений, освоившее почти все экологические ниши на Земле. Ограниченные в силу узкой специализации возможности опыления компенсируются у них невероятно высоким количеством семян, которое образуется в каждой коробочке. Ч. Дарвин подсчитал, что если бы все семена пальчатокоренника пятнистого (Dactylorhiza maculata, табл. 33, 3) благополучно прорастали, то правнуки одного растения «почти могли бы покрыть однообразным зеленым ковром всю поверхность суши на земном шаре». И того небольшого количества семян, которые превращаются во взрослые растения, орхидным вполне достаточно для продолжения рода в ненарушенных природных условиях.
Жизнь растений: в 6-ти томах. — М.: Просвещение. Под редакцией А. Л. Тахтаджяна, главный редактор чл.-кор. АН СССР, проф. А.А. Федоров. 1974.
.