- КЛАСС ДВУСТВОРЧАТЫЕ (BVALVA)
- КЛАСС ДВУСТВОРЧАТЫЕ (BVALVA)
-
Класс двустворчатых моллюсков имеет, как известно, четыре различных названия, каждое из которых в какой-то мере отражает главнейшие черты их строения. Название «двустворчатые» (Bivalvia) было впервые предложено Линнеем (1758) и является наиболее правильным, поскольку оно применимо ко всем представителям этого класса. Безголовыми (Асеphala) они были названы Линком (1807), что отразило факт редукции у них головного отдела тела по мере развития у них в процессе эволюции двух створок раковины и смыкания этих створок вокруг тела моллюска. Третье название — «пластинчатожаберные» (Lamellibranchia), предложенное Блэнвилем в 1814 г., может быть полностью применимо лишь к одному отряду данного класса, поскольку остальные отряды имеют жабры иного строения; таким образом, это название неприменимо, как и четвертое — «топороногие» (Pelecypoda, Гольдфус, 1880), так как строение ноги у двустворчатых моллюсков весьма разнообразно. Таким образом, наиболее правильным и всеобъемлющим является первое, линнеевское наименование, которое и следует сохранить применительно к этому классу.Двустворчатые широко распространены в Мировом океане и его краевых морях, в реках и озерах и даже в прудах.Общее количество видов двустворчатых составляет около пятнадцати тысяч, и большая их часть связана в своем обитании с солеными морскими водами, и лишь около одной пятой общего числа их видов населяет пресные воды. На суше двустворчатые моллюски не встречаются.В морских водах они распространены чрезвычайно широко, встречаясь во всех климатических зонах, от теплых вод тропических морей до Арктики и Антарктики и до холодных глубин океанической абиссали. Они населяют практически все глубины Мирового океана — от приливоотливной зоны (литорали) и прибрежных мелководий вплоть до огромных глубин впадин Мирового океана, где они были: найдены на глубине почти 10, 8 км.Особенно изобилуют двустворчатые моллюски в прибрежных мелководных районах морей (до глубины 100—200 м), где они по своей биомассе (весу на 1 м2 площади дна) и по плотности поселений составляют часто большую часть всей обитающей здесь донной фауны.В настоящее время количество видов глубоководных двустворчатых моллюсков, обитающих в абиссали Мирового океана (т. е. на глубине более 2000 м), судя по неполным еще данным, составляет около 400 видов, но и это число следует считать сильно преуменьшенным.Велико разнообразие размеров, строения и окраски раковин двустворчатых моллюсков. Так, гигант среди моллюсков вообще, обитатель тропических морей тридакна может достигать 200 кг веса, а длина ее мощной раковины 1, 4 м. Наряду с этим размеры ряда обычных глубоководных моллюсков не превышают 2-3 мм.Пестрыми узорами и яркими красками блещут раковины и края мантии многих тропических мелководных моллюсков, незаметных среди зарослей водорослей, белых, розоватых, лиловых и желтоватых кораллов, ярких звезд и других беспозвоночных. Разнообразные выросты, шипы, чешуйки и ребра украшают створки этих моллюсков, что помогает им укрепляться в этих зарослях и противостоять действию морских волн и течений.Более скромную окраску имеют раковины моллюсков, живущих на песчаных или илистых грунтах умеренной или арктической области.Глубоководные формы имеют, как правило, бледно окрашенную раковину, часто очень тонкую и полупрозрачную.Скромно окрашена в зеленоватые и коричневатые тона большая часть пресноводных форм.Большое разнообразие строения тела и раковины двустворчатых моллюсков тесно связано с образом их жизни, местом их обитания, с глубиной и с качеством грунта, на котором они живут, или прикрепляются, или в который закапываются. Это сказывается прежде всего на строении их раковины; на наличии на ней ребер с тем или иным вооружением ее так называемого замка, при помощи которого скрепляются створки; на наличии или -отсутствии сифонов — особых выростов мантии (двух мягких лопастей, окружающих тело моллюска и выделяющих его раковину); на форме и размерах ноги и наличии в ней особой железы, выделяющей нити так называемого б и с с у с а, при помощи которого они могут прикрепляться к грунту, а также и на многом другом. У форм, закапывающихся более или менее глубоко в мягкий грунт, сзади развиваются особые выросты мантии — сифоны, через которые засасывается и выводится вода, необходимая для дыхания и питания моллюска, погруженного в грунт. Таковы различные макомы, теллины, йольдии и др. Моллюски, живущие на поверхности грунта, ползающие или слегка закапывающиеся в него, имеют лишь зачаточные сифоны или вовсе лишены их (например, сердцевидки, венуси, астарты и др.). Моллюски, обитающие в прибрежных мелководных районах на более жестких песчаных грунтах с примесью камней, имеют более крепкие, толстые раковины (например, арки, скафарки, морские гребешки — пектены и хламисы), а различные обитатели мягких илистых грунтов имеют более тонкие раковины (батиарки, морские гребешки пропеамуссиумы и др.).Многие формы, живущие на прибрежных мелководьях, прикрепляются нитями биссуса к камням, скалам, друг к другу, нередко образуя целые гроздья, сростки (многие мидии), или даже прирастают своими створками к камням, или срастаются друг с другом (устрицы).Очень крепкой раковиной с острыми зубчиками на ребрах обладают многие моллюски-камнеточцы; некоторые из них выделяют особый кислый секрет, растворяющий известь прибрежных скал и камней, в которых они протачивают себе норки. Мягкое червеобразное тело древоточца тередо (Teredo) прикрыто лишь спереди маленькой сложной раковиной, служащей ему для сверления, а не для защиты тела; проводя всю жизнь в прогрызаемых в древесине ходах, эти моллюски не нуждаются в защите своего слабого длинного тела раковиной. Наконец, огромное разнообразие различных тропических моллюсков, постоянных обитателей коралловых рифов, тесно связано с их жизнью в мелководной, крайне разнообразной по характеру субстрата зоне.Крепкие известковые раковины двустворчатых, как и других моллюсков, хорошо сохраняются в отложениях (глинах и песках) в течение целых геологических эпох. Остатки их поселений чрезвычайно ценны для геологов и палеонтологов. Эти остатки могут прекрасно характеризовать не только гидрологические и климатические условия, при которых эти отложения образовались (т. е. при которых жили найденные здесь виды моллюсков), но и возраст данной толщи отложений. Так, скопления ископаемых раковин ныне живущего в арктических морях холодноводного моллюска портландии арктической (Portlandia arctica) в отложениях севера Европы прекрасно указывают на то, что эти районы были ранее заняты холодными, слегка распресненными водами мелководного так называемого Йольдиева моря. Это море с холодноводной фауной, где руководящую роль играла арктическая портландия, было связано с периодом похолодания в послеледниковое время (примерно около 8—10 тыс. лет назад). И наоборот, отложения теплого Литторинового моря, образовавшегося позднее (3—5 тыс. лет назад), характеризуются присутствием остатков совсем других, тепловодных моллюсков, как например исландской циприны (Cyprina islandica), съедобной сердцевидки (Cerastoderma edu1е), цирфеи гребенчатой (Zirfaea crispata) и др. Эти виды живут теперь лишь в Северной Атлантике, в наиболее теплых районах Баренцева и отчасти Белого морей, тогда как в эпоху Литторинового моря они продвигались дальше на север.Представители класса двустворчатых моллюсков впервые появляются в отложениях в палеозое, т. е. в древнейших отложениях нашей планеты, а именно в верхнекембрийских слоях, образование которых насчитывает около 450—500 млн. лет. Найденные здесь первые двустворчатые моллюски относились к четырем родам, из которых такие, как Ctenodonta и Paleoneilo, имели гребенчатый замок и внешне напоминали современных ореховидок (Nuculidae) и маллеций (Malletiidae) из отряда гребенчатозубых (Тахоdonta). Наибольшего видового разнообразия двустворчатые моллюски достигали в меловое время, т. е. за 100— 130 млн. лет до нашего времени.Таким образом, класс двустворчатых моллюсков является одной из самых древних групп донных беспозвоночных.С давних времен многие двустворчатые моллюски используются человеком, они служили и служат предметом добычи. Их раковины постоянно встречаются в так называемых «кухонных кучах» доисторического человека, жившего у берегов морей, рек, озер. В раскопках стоянок человека палеолита в Крыму неизменно находят большое количество раковин устриц, мидий, гребешков и других моллюсков, которых промышляют ив настоящее время. Двустворчатых моллюсков добывают из-за их вкусного, очень полезного и легко усвояемого организмом человека мяса (как, например, устриц, мидий, морских гребешков, петушков-тапесов и венеруписов, мактр, песчаную ракушку, сердцевидок, арок, морских черенков и синовакул, пресноводных перловиц, лампсилии, беззубок, корбикул и др.).По калорийности они могут даже превосходить мясо многих рыб, как морских, так и пресноводных. Пищевая ценность мяса моллюсков обусловливается также большим содержанием в них витаминов А, В, С, D и др., высоким содержанием таких редких в обычной пище человека минеральных веществ, как йод, железо, цинк, медь и др. Последние, как известно, входя в состав ряда ферментов, гормонов, играют исключительно большую роль в окислительном, углеводном и белковом обмене веществ, в регуляции гормональной деятельности. Мясо и раковины моллюсков широко используют для изготовления кормовой муки, идущей на откорм домашней птицы, а также для изготовления удобрительных туков.В последние десятилетия, ввиду того что естественные природные запасы наиболее ценных съедобных моллюсков (даже в морях) истощены, а спрос на них продолжает увеличиваться, во многих странах их стали переселять в новые районы, акклиматизировать, а также и разводить искусственно как в морских, так и в пресных водах, на «фермах»—специально подготовленных отмелях и в охраняемых от хищников небольших бухтах, искусственных водоемах. Успешно разводят и культивируют не только морских моллюсков (устриц, мидий, морских петушков, тапесов), но и пресноводных (лампсилин).В настоящее время значительно более половины добываемых двустворчатых моллюсков получают в результате их искусственного разведения. Вылов моллюсков в местах их естественного обитания в водоемах и их искусственное разведение стали теперь в ряде стран доходной и важной частью пищевой индустрии.Двустворчатых моллюсков теперь нередко добывают на больших судах специально сконструированными орудиями лова; широко используется отлов моллюсков водолазами. Моллюски поступают на рынок не только в свежем и сушеном виде, но особенно в мороженом; сильно развилось также приготовление различных консервов из моллюсков.Добыча двустворчатых моллюсков в последние десятилетия резко возросла. Если перед началом мировой войны ежегодная добыча их составляла около 5 млн. цу то уже в 1962 г. она возросла примерно до 17 млн. ц и стала составлять около 50% мировой добычи всех морских беспозвоночных, или 4 % от общей мировой добычи (426 млн. ц) всех продуктов моря (рыбы, китов, беспозвоночных, водорослей).Наибольшее количество (около 90%) двустворчатых моллюсков добывается в северном полушарии — в Тихом и в Атлантическом океанах. Промысел пресноводных двустворчатых моллюсков дает лишь несколько процентов от общей их мировой добычи. Особенно большое значение имеет промысел двустворчатых моллюсков в таких странах, как Япония, США, Корея, Китай, Индонезия, Филиппинские острова и другие острова Тихого океана. Так, в Японии добывается около 90 видов двустворчатых моллюсков, из которых около двух десятков видов имеют большое промысловое значение, а 10 видов разводятся искусственно. В европейских странах промысел и разведение двустворчатых моллюсков наиболее развиты во Франции, Италии и др.В СССР промысловое значение имеют главным образом крупный приморский гребешок (Pecten (Patinopecten) yessoensis), а также различные мидии, белая ракушка (Spisula sachalinensis), песчаная ракушка (Муа (Arenomya) arenaria), петушки (Tapes, Venerupis) и некоторые другие.Двустворчатые моллюски издавна добывались и ради их раковин, дающих не только прекрасное сырье для перламутровых изделий (многие пресноводные перловицы и жемчужницы, морские жемчужницы — пинктады, птерии и др.)» но и ценнейший жемчуг. В начале текущего столетия были найдены промышленные способы более быстрым искусственным путем получать жемчуг (находки которого являются в природных условиях довольно редкой случайностью), не отличимый от жемчуга, образовавшегося естественным путем. Фермы для содержания морских жемчужниц и выращивания в них жемчуга особенно успешно работают в Японии. Так, уже в 1936 г. здесь было выращено 140 тыс. раковин морских жемчужниц и получено 26, 5 тыс. жемчужин.В некоторых странах, особенно в тропической части Тихого океана, добываемые раковины двустворчатых моллюсков широко используются для получения извести.Почти все двустворчатые моллюски, за исключением крупных форм с крепкой, толстой раковиной, служат излюбленной пищей придонных рыб — бентофагов (т. е. питающихся донными животными), в том числе и многих промысловых рыб, как морских, так и пресноводных: камбал, некоторых тресковых (пикши), осетровых, многих карповых (лещ, сазан), зубаток, бычков и др. Некоторые рыбы благодаря преобладанию в их пище мелких моллюсков получили название «моллюскоедов», как, например, каспийская вобла. Районы, где наряду с другими донными животными (многощетинковыми червями, офиурами и др.) наблюдается массовое развитие мелких двустворчатых моллюсков, служат местами откорма различных придонных промысловых рыб.Моллюсков охотно поедают многие крупные десятиногие раки (омары, ракиотшельники, крабы), морские звезды— исконные враги двустворчатых моллюсков. Промысловые устричные банки периодически очищают от морских звезд при помощи особых швабр, которые волокут по дну небольшие суда.Немалую роль играют двустворчатые в питании промысловых камчатских «крабов» (Paralithodes kamtschatica).
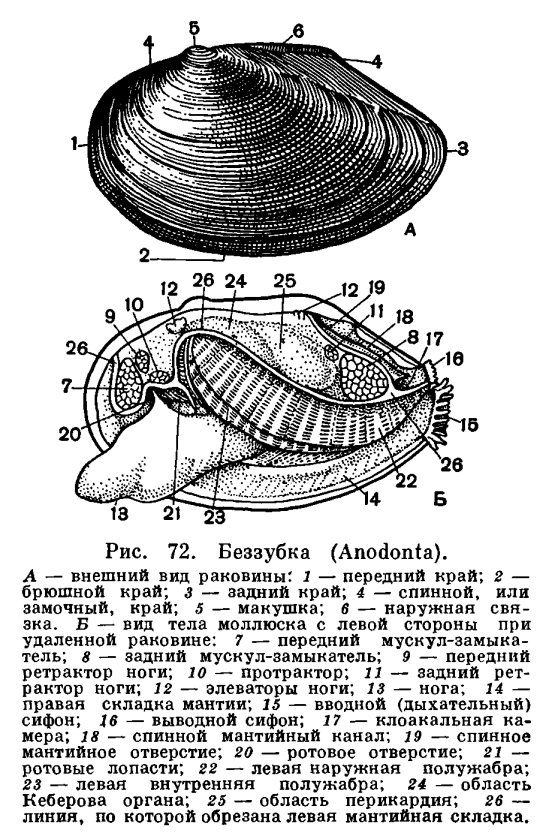 Каковы же основные черты строения двустворчатых моллюсков? Чтобы легче понять их устройство, представим себе книгу в переплете, поставленную корешком вверх. Обе половинки переплета будут соответствовать правой и левой створкам раковины, охватывающим тело моллюска с боков. Корешок книги будет аналогичен эластичной наружной связке (лигаменту), соединяющей обе створки на спинной стороне раковины и одновременно растягивающей их. Первый и последний листы книги соответствуют двум лопастям мантии, охватывающим тело с правой и левой сторон, а следующие два листа книги спереди и сзади будут аналогичны двум парам жабр с каждой стороны тела. И наконец, между обеими парами жабр внутри расположено само тело и нога — обычно довольно крупный мускулистый топоровидный или клиновидный орган, направленный вперед; у прикрепленных или малоподвижных форм нога может превращаться в небольшой вырост, и, наоборот, у активно двигающихся видов (например, у сердцевидок) нога становится сильной, слегка коленчатоизогнутой, приспособленной для продвижения в мягком песчанистом грунте.Расположение частей тела двустворчатого моллюска станет яснее, если рассмотреть вскрытого моллюска, например беззубку, обычную в наших пресноводных водоемах с илистым дном и медленно текущей или стоячей водой. Наиболее распространенной является обыкновенная беззубка (Anodonta cygnea) — довольно крупный моллюск из отряда настоящих пластинчатожаберных (Eulamellibranchia). При осмотре моллюска важно определить передний и задний концы раковины. Передний конец у беззубки легко узнать по более округлой форме раковины и по направленной вперед ноге; на заднем, несколько более суженном конце между створок видны короткие выросты мантии— сифоны. Вдоль верхнего спинного края, позади макушек, расположена довольно крупная наружная связк а , или л и г а м е н т, — упругий эластичный тяж, при «сокращении» которого створки раскрываются. Он состоит из волокнистого рогового вещества, близкого к хитину, — конхиолина: образуется он из наружного покрова самой раковины (периострака). «Работа» лигамента определяется взаимодействием различно расположенных волокон конхиолина, из которых он состоит. Когда мускулы-замыкатели, сокращаясь, стягивают створки раковины, то волокна в нижней части лигамента сжимаются, а в верхней — растягиваются, а при расслаблении мускулов — наоборот; поэтому у мертвых моллюсков створки раковины всегда полуоткрыты. У двустворчатых моллюсков связка может быть наружной или внутренней или той и другой вместе.У анодонты нет замковых зубов и спинной край гладкий, отсюда и ее название— беззубка (Anodonta). У большинства же двустворчатых моллюсков для более прочцого соединения створок между собой под макушкой, изнутри, на спинном или замоч ном краю раковины, имеются разнообразные (по форме, числу и расположению) выросты, так называемые зубы, каждый из которых входит в соответствующее углубление на противоположной створке. Все это, вместе взятое, образует замок раковины. Устройство замка, характер, количество и расположение его зубов служит важным систематическим признаком у двустворчатых моллюсков и характерно для семейства, рода и вида. Лигамент также является частью замкового аппарата двустворчатых моллюсков, поскольку служит для соединения створок между собой.Поверхность раковины у большинства двустворчатых моллюсков, в том числе и у беззубки, покрыта различно окрашенным наружным слоем, или периостраком. Он легко соскабливается ножом, и тогда под ним обнажается белый фарфоровидный, или призматический, известковый слой (ostracum). На нем хорошо видны концентрические линии — следы нарастания раковины, идущие параллельно ее краям. Внутренняя поверхность раковины у многих моллюсков, в том числе и у беззубки, выстлана перламутровым слоем (hypostracum).Периострак, состоящий из конхиолина, устойчив к внешним воздействиям (как механическим, так и химическим) и служит, таким образом, хорошей защитой для внутреннего известкового слоя раковины. Особенно хорошо заметна стойкость периострака к действию растворенной в морской воде угольной кислоты. Она может накапливаться у дна, в самых придонных слоях, и в грунте, где живут моллюски (благодаря разложению органического вещества, отчасти и дыханию водных организмов), и возрастать по мере увеличения глубины и давления. Так, в Карском море часто встречаются совсем мягкие отмершие раковины астарт, йольдий или портландий с растворенной известковой частью раковины, от которой остался один неповрежденный мягкий роговой слой — периостракум.Оба других слоя раковины состоят из известковых призмочек или пластинок, соединенных небольшим количеством конхиолина. В среднем (фарфоровом) слое они располагаются перпендикулярно к поверхности раковины, а во внутреннем (перламутровом) — параллельно ей; благодаря такому расположению получается интерференция света, которая и дает блеск и радужные переливы перламутра. Чем тоньше пластинки этого слоя, тем красивее и ярче этот блеск. Наиболее красивый перламутр бывает у тех моллюсков, у которых толщина пластинок перламутра в слое составляет 0, 4—0, 6 мк.Раковина моллюска образуется в результате секреторной работы его мантии: вдоль ее края расположено большое количество железистых клеток, вырабатывающих различные слои раковины. Так, клетки особой мантийной бороздки, идущей вдоль всего края мантии, образуют наружный конхиолиновый слой, эпителиальные клетки так называемой краевой складки дают призматический слой раковины, а наружная поверхность мантии выделяет перламутровый слой.Раковина двустворчатых моллюсков, состоящая более чем на 90% из СаС03, содержит его в виде кальцита или арагонита, находящихся в различных соотношениях. У тропических моллюсков раковина содержит больше арагонита, а также довольно много стронция. Кристаллографическое изучение состава раковин ископаемых моллюсков позволяет теперь судить о температуре тех морей, в которых жили эти моллюски.Кальций, откладывающийся в раковину мантией, поступает в нее не только через кровь, куда она попадает из пищи через кишечник, но, как показали последние эксперименты с радиоактивным кальцием, клетки мантии могут и сами извлекать кальций из воды.Рост раковины происходит как путем общего утолщения створок в результате наслаивания все новых и новых известковых пластинок на внутренней поверхности створок, так и путем нарастания всей раковины по ее свободному краю. При наступлении неблагоприятных условий (в зимний период, при ухудшении питания и т. д.) рост раковины замедляется или даже приостанавливается, что хорошо заметно на поверхности раковины у мно гих моллюсков, где в это время образуются характерные сгущения линий, имеющих вид концентрических полос, идущих параллельно брюшному краю раковины. По зимним кольцам — сезонным остановкам роста — иногда можно определить приблизительно возраст моллюска. Однако у некоторых видов такие кольца неразличимы; у тропических форм, где нет сезонных явлений, такие кольца обычно вообще не образуются. У наших пресноводных перловиц и беззубок такие сезонные зимние перерывы роста имеются, поэтому годовые кольца выражены обычно хорошо.Чтобы открыть створки раковины беззубки, надо прежде всего перерезать два имеющихся у нее внутри спереди и сзади довольно сильных мускула-замыкателя, стягивающих обе створки в поперечном направлении и закрывающих раковину. У живой беззубки легче сломать ее тонкую раковину, чем открыть ее, не перерезав этих мускулов.Когда мускулы перерезаны, то створки сами свободно раскрываются, растягиваемые лигаментом, и можно видеть две мягкие полупрозрачные розоватые или желтоватые лопасти — мантию, охватывающую тело с боков. Края мантии слегка утолщены. Этим местом она прикрепляется к раковине, на внутренней поверхности створок которой образуется так называемая мантийная линия. Мантия беззубок, срастаясь сзади, образует два коротких сифона, опушенных короткими чувствительными выростами.У моллюсков, закапывающихся в грунт, образуются длинные сократимые сифоны; места прикрепления втягивающих их мышц образуют на внутренней поверхности раковины отпечаток, так называемый синус. Чем глубже синус, тем длиннее сифоны у моллюсков, тем глубже они могут закапываться в грунт.На брюшной стороне из-под края мантии вперед выдается довольно крупная клиновидная нога, направленная острым концом вперед. Нога у беззубки очень подвижна (как и у многих других моллюсков), и ее действие легко наблюдать в аквариуме. Как только анодонта успокоится, ее раковина приоткрывается, показываются розовато-желтые края мантии и наружу высовывается кончик ноги. Если все кругом спокойно, нога высовывается еще дальше (у крупных анодонт на 4—5 см), погружается в песок, и моллюск начинает двигаться вперед или закапываться в грунт передним концом, слегка приподнимаясь на своей ноге. На пройденном ею пути остается след в виде неглубокой бороздки.Большая подвижность ноги беззубки обусловливается главным образом сокращением имеющихся в ней различных групп гладких мышц. Имеются парные передний и задний мускулы: ретракторы, втягивающие ногу косо вверх, протракторы, выдвигающие ногу вперед, и группа более мелких мускулов-поднимателей (элеваторов) ноги кверху. Все эти мышцы прикрепляются к внутренней поверхности створок раковины, где отпечатки мест их прикрепления довольно хорошо заметны (у ретракторов вдоль замочного края раковины). Кроме того, в ноге имеется еще целый ряд более мелких мышц, не прикрепляющихся к створкам и расположенных в ноге послойно и поперечно.
Каковы же основные черты строения двустворчатых моллюсков? Чтобы легче понять их устройство, представим себе книгу в переплете, поставленную корешком вверх. Обе половинки переплета будут соответствовать правой и левой створкам раковины, охватывающим тело моллюска с боков. Корешок книги будет аналогичен эластичной наружной связке (лигаменту), соединяющей обе створки на спинной стороне раковины и одновременно растягивающей их. Первый и последний листы книги соответствуют двум лопастям мантии, охватывающим тело с правой и левой сторон, а следующие два листа книги спереди и сзади будут аналогичны двум парам жабр с каждой стороны тела. И наконец, между обеими парами жабр внутри расположено само тело и нога — обычно довольно крупный мускулистый топоровидный или клиновидный орган, направленный вперед; у прикрепленных или малоподвижных форм нога может превращаться в небольшой вырост, и, наоборот, у активно двигающихся видов (например, у сердцевидок) нога становится сильной, слегка коленчатоизогнутой, приспособленной для продвижения в мягком песчанистом грунте.Расположение частей тела двустворчатого моллюска станет яснее, если рассмотреть вскрытого моллюска, например беззубку, обычную в наших пресноводных водоемах с илистым дном и медленно текущей или стоячей водой. Наиболее распространенной является обыкновенная беззубка (Anodonta cygnea) — довольно крупный моллюск из отряда настоящих пластинчатожаберных (Eulamellibranchia). При осмотре моллюска важно определить передний и задний концы раковины. Передний конец у беззубки легко узнать по более округлой форме раковины и по направленной вперед ноге; на заднем, несколько более суженном конце между створок видны короткие выросты мантии— сифоны. Вдоль верхнего спинного края, позади макушек, расположена довольно крупная наружная связк а , или л и г а м е н т, — упругий эластичный тяж, при «сокращении» которого створки раскрываются. Он состоит из волокнистого рогового вещества, близкого к хитину, — конхиолина: образуется он из наружного покрова самой раковины (периострака). «Работа» лигамента определяется взаимодействием различно расположенных волокон конхиолина, из которых он состоит. Когда мускулы-замыкатели, сокращаясь, стягивают створки раковины, то волокна в нижней части лигамента сжимаются, а в верхней — растягиваются, а при расслаблении мускулов — наоборот; поэтому у мертвых моллюсков створки раковины всегда полуоткрыты. У двустворчатых моллюсков связка может быть наружной или внутренней или той и другой вместе.У анодонты нет замковых зубов и спинной край гладкий, отсюда и ее название— беззубка (Anodonta). У большинства же двустворчатых моллюсков для более прочцого соединения створок между собой под макушкой, изнутри, на спинном или замоч ном краю раковины, имеются разнообразные (по форме, числу и расположению) выросты, так называемые зубы, каждый из которых входит в соответствующее углубление на противоположной створке. Все это, вместе взятое, образует замок раковины. Устройство замка, характер, количество и расположение его зубов служит важным систематическим признаком у двустворчатых моллюсков и характерно для семейства, рода и вида. Лигамент также является частью замкового аппарата двустворчатых моллюсков, поскольку служит для соединения створок между собой.Поверхность раковины у большинства двустворчатых моллюсков, в том числе и у беззубки, покрыта различно окрашенным наружным слоем, или периостраком. Он легко соскабливается ножом, и тогда под ним обнажается белый фарфоровидный, или призматический, известковый слой (ostracum). На нем хорошо видны концентрические линии — следы нарастания раковины, идущие параллельно ее краям. Внутренняя поверхность раковины у многих моллюсков, в том числе и у беззубки, выстлана перламутровым слоем (hypostracum).Периострак, состоящий из конхиолина, устойчив к внешним воздействиям (как механическим, так и химическим) и служит, таким образом, хорошей защитой для внутреннего известкового слоя раковины. Особенно хорошо заметна стойкость периострака к действию растворенной в морской воде угольной кислоты. Она может накапливаться у дна, в самых придонных слоях, и в грунте, где живут моллюски (благодаря разложению органического вещества, отчасти и дыханию водных организмов), и возрастать по мере увеличения глубины и давления. Так, в Карском море часто встречаются совсем мягкие отмершие раковины астарт, йольдий или портландий с растворенной известковой частью раковины, от которой остался один неповрежденный мягкий роговой слой — периостракум.Оба других слоя раковины состоят из известковых призмочек или пластинок, соединенных небольшим количеством конхиолина. В среднем (фарфоровом) слое они располагаются перпендикулярно к поверхности раковины, а во внутреннем (перламутровом) — параллельно ей; благодаря такому расположению получается интерференция света, которая и дает блеск и радужные переливы перламутра. Чем тоньше пластинки этого слоя, тем красивее и ярче этот блеск. Наиболее красивый перламутр бывает у тех моллюсков, у которых толщина пластинок перламутра в слое составляет 0, 4—0, 6 мк.Раковина моллюска образуется в результате секреторной работы его мантии: вдоль ее края расположено большое количество железистых клеток, вырабатывающих различные слои раковины. Так, клетки особой мантийной бороздки, идущей вдоль всего края мантии, образуют наружный конхиолиновый слой, эпителиальные клетки так называемой краевой складки дают призматический слой раковины, а наружная поверхность мантии выделяет перламутровый слой.Раковина двустворчатых моллюсков, состоящая более чем на 90% из СаС03, содержит его в виде кальцита или арагонита, находящихся в различных соотношениях. У тропических моллюсков раковина содержит больше арагонита, а также довольно много стронция. Кристаллографическое изучение состава раковин ископаемых моллюсков позволяет теперь судить о температуре тех морей, в которых жили эти моллюски.Кальций, откладывающийся в раковину мантией, поступает в нее не только через кровь, куда она попадает из пищи через кишечник, но, как показали последние эксперименты с радиоактивным кальцием, клетки мантии могут и сами извлекать кальций из воды.Рост раковины происходит как путем общего утолщения створок в результате наслаивания все новых и новых известковых пластинок на внутренней поверхности створок, так и путем нарастания всей раковины по ее свободному краю. При наступлении неблагоприятных условий (в зимний период, при ухудшении питания и т. д.) рост раковины замедляется или даже приостанавливается, что хорошо заметно на поверхности раковины у мно гих моллюсков, где в это время образуются характерные сгущения линий, имеющих вид концентрических полос, идущих параллельно брюшному краю раковины. По зимним кольцам — сезонным остановкам роста — иногда можно определить приблизительно возраст моллюска. Однако у некоторых видов такие кольца неразличимы; у тропических форм, где нет сезонных явлений, такие кольца обычно вообще не образуются. У наших пресноводных перловиц и беззубок такие сезонные зимние перерывы роста имеются, поэтому годовые кольца выражены обычно хорошо.Чтобы открыть створки раковины беззубки, надо прежде всего перерезать два имеющихся у нее внутри спереди и сзади довольно сильных мускула-замыкателя, стягивающих обе створки в поперечном направлении и закрывающих раковину. У живой беззубки легче сломать ее тонкую раковину, чем открыть ее, не перерезав этих мускулов.Когда мускулы перерезаны, то створки сами свободно раскрываются, растягиваемые лигаментом, и можно видеть две мягкие полупрозрачные розоватые или желтоватые лопасти — мантию, охватывающую тело с боков. Края мантии слегка утолщены. Этим местом она прикрепляется к раковине, на внутренней поверхности створок которой образуется так называемая мантийная линия. Мантия беззубок, срастаясь сзади, образует два коротких сифона, опушенных короткими чувствительными выростами.У моллюсков, закапывающихся в грунт, образуются длинные сократимые сифоны; места прикрепления втягивающих их мышц образуют на внутренней поверхности раковины отпечаток, так называемый синус. Чем глубже синус, тем длиннее сифоны у моллюсков, тем глубже они могут закапываться в грунт.На брюшной стороне из-под края мантии вперед выдается довольно крупная клиновидная нога, направленная острым концом вперед. Нога у беззубки очень подвижна (как и у многих других моллюсков), и ее действие легко наблюдать в аквариуме. Как только анодонта успокоится, ее раковина приоткрывается, показываются розовато-желтые края мантии и наружу высовывается кончик ноги. Если все кругом спокойно, нога высовывается еще дальше (у крупных анодонт на 4—5 см), погружается в песок, и моллюск начинает двигаться вперед или закапываться в грунт передним концом, слегка приподнимаясь на своей ноге. На пройденном ею пути остается след в виде неглубокой бороздки.Большая подвижность ноги беззубки обусловливается главным образом сокращением имеющихся в ней различных групп гладких мышц. Имеются парные передний и задний мускулы: ретракторы, втягивающие ногу косо вверх, протракторы, выдвигающие ногу вперед, и группа более мелких мускулов-поднимателей (элеваторов) ноги кверху. Все эти мышцы прикрепляются к внутренней поверхности створок раковины, где отпечатки мест их прикрепления довольно хорошо заметны (у ретракторов вдоль замочного края раковины). Кроме того, в ноге имеется еще целый ряд более мелких мышц, не прикрепляющихся к створкам и расположенных в ноге послойно и поперечно.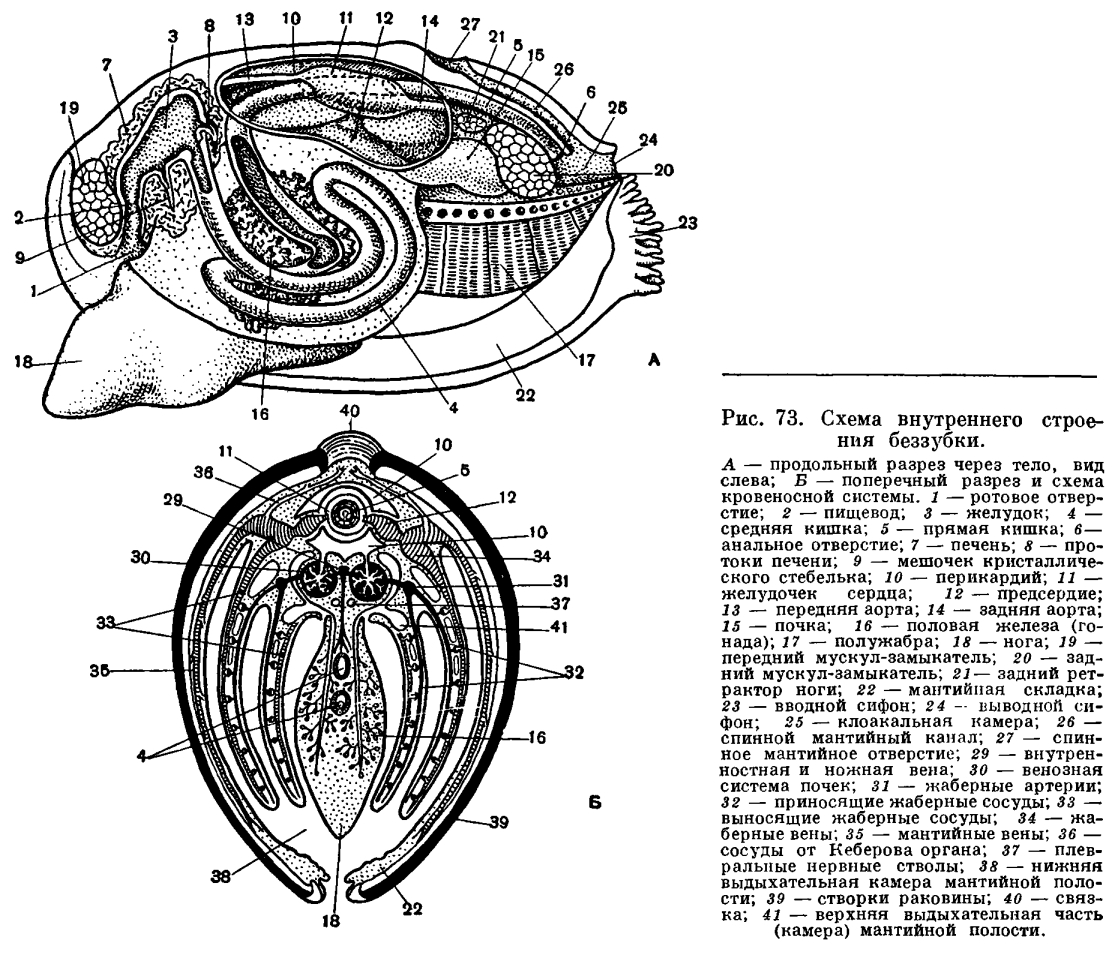 Если отвернуть мантийную лопасть кверху, то откроется мантийная полость беззубки, где расположены ее основные органы: ротовые лопасти, коричневатые жаберные листки (по два с каждой стороны тела), нога, основание которой находится между правой и левой жабрами. Спереди, в углублении между ногой и передним мускулом-замыкателем, помещается ротовое отверстие, окруженное двумя парами небольших треугольных сократимых околоротовых лопастей. Каждая жабра у анодонты состоит из двух листков полужабр, которые в свою очередь сложены двумя пластинками — восходящей и нисходящей.
Если отвернуть мантийную лопасть кверху, то откроется мантийная полость беззубки, где расположены ее основные органы: ротовые лопасти, коричневатые жаберные листки (по два с каждой стороны тела), нога, основание которой находится между правой и левой жабрами. Спереди, в углублении между ногой и передним мускулом-замыкателем, помещается ротовое отверстие, окруженное двумя парами небольших треугольных сократимых околоротовых лопастей. Каждая жабра у анодонты состоит из двух листков полужабр, которые в свою очередь сложены двумя пластинками — восходящей и нисходящей.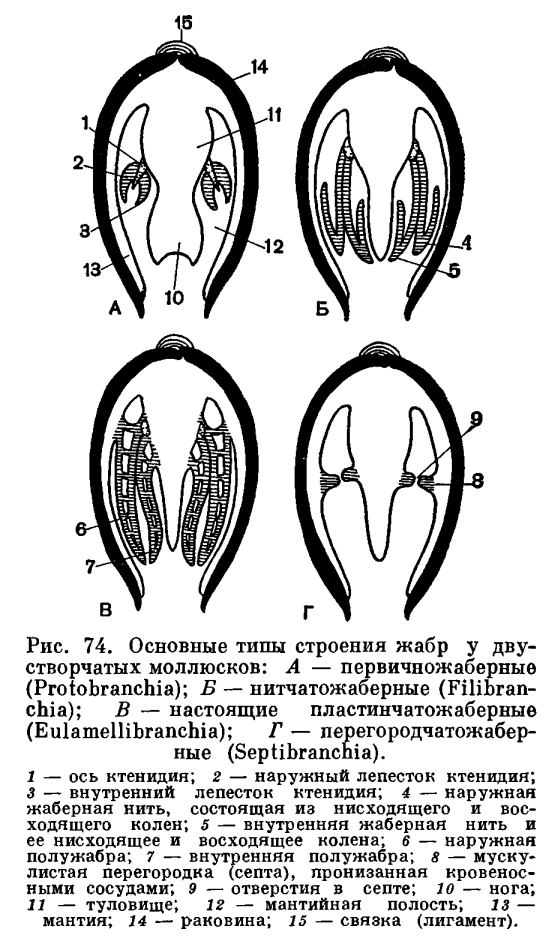 Каждая жаберная пластинка состоит из рядов отдельных нитей (филаментов), а каждая нить соответственно образует восходящее и нисходящее колена. У анодонты между соседними нитями и между образуемыми ими коленами имеются сосудистые соединения (мостики), что характерно для всего отряда настоящих пластинчатожаберных. Каждая полужабра представляет собой, таким образом, решетчатую, сложно продырявленную двуслойную пластинку.У представителей других отрядов двустворчатых моллюсков жабры имеют иное устройство (о чем будет сказано ниже).Мантийная полость и расположенные в ней жабры все время омываются током воды, который создается главным образом мерцанием мельчайших ресничек эпителия, покрывающего поверхность мантии, жабр, ротовых лопастей и стенок тела. Вода поступает в мантийную полость беззубки через нижний (дыхательный) сифон, попадая сначала в ее большую, нижнюю часть — дыхательную камеру, затем процеживается через щели в жабрах и уходит в верхнюю часть мантийной полости —в ыдыхательную камеру, откуда наконец и выходит наружу через верхний (выводной, или анальный) сифон. Засасывание воды через входной сифон происходит в результате разницы в гидростатическом давлении между поджаберным и наджаберным пространствами мантийной полости и между нею и окружающей моллюска водой; эта разница вызывается не только работой ресничного эпителия, но также и сокращением жаберных нитей и мускулатуры мантии и сифонов. Когда ток воды замедляется, входя в большую «вдыхательную камеру» мантии, грубые и крупные частицы выпадают из него и оседают на поверхности мантии, а затем удаляются из моллюска. Легко убедиться в наличии интенсивных токов воды, поступающей в мантийную полость, если положить беззубку в неглубокий сосуд с водой так, чтобы вода лишь слегка покрывала раковину. Дав ей успокоиться, надо подсыпать в воду вблизи заднего ее конца какойнибудь порошок, остающийся взвешенным в воде (например, тушь, кармин, сухие тертые водоросли). Тогда можно увидеть, как крупинки порошка уходят через нижний (вводной) сифон внутрь раковины и через некоторое время с сильной струей воды выбрасываются наружу через верхний (выводной) сифон. Время от времени беззубка, часто без всякого внешнего раздражения, с силой захлопывает створки раковины и выбрасывает наружу струи воды, обновляя сразу всю воду, содержащуюся в мантийной полости. Вскоре створки раковины опять приоткрываются и нормальная медленная циркуляция воды возобновляется.Чтобы убедиться в интенсивности работы ресничного эпителия, можно вырезать кусочек мантии беззубки и положить его внутренней поверхностью книзу на дно сосуда. Благодаря продолжающейся еще некоторое время работе ресничек этот кусочек будет слегка перемещаться и даже слегка ползти по наклонной плоскости.
Каждая жаберная пластинка состоит из рядов отдельных нитей (филаментов), а каждая нить соответственно образует восходящее и нисходящее колена. У анодонты между соседними нитями и между образуемыми ими коленами имеются сосудистые соединения (мостики), что характерно для всего отряда настоящих пластинчатожаберных. Каждая полужабра представляет собой, таким образом, решетчатую, сложно продырявленную двуслойную пластинку.У представителей других отрядов двустворчатых моллюсков жабры имеют иное устройство (о чем будет сказано ниже).Мантийная полость и расположенные в ней жабры все время омываются током воды, который создается главным образом мерцанием мельчайших ресничек эпителия, покрывающего поверхность мантии, жабр, ротовых лопастей и стенок тела. Вода поступает в мантийную полость беззубки через нижний (дыхательный) сифон, попадая сначала в ее большую, нижнюю часть — дыхательную камеру, затем процеживается через щели в жабрах и уходит в верхнюю часть мантийной полости —в ыдыхательную камеру, откуда наконец и выходит наружу через верхний (выводной, или анальный) сифон. Засасывание воды через входной сифон происходит в результате разницы в гидростатическом давлении между поджаберным и наджаберным пространствами мантийной полости и между нею и окружающей моллюска водой; эта разница вызывается не только работой ресничного эпителия, но также и сокращением жаберных нитей и мускулатуры мантии и сифонов. Когда ток воды замедляется, входя в большую «вдыхательную камеру» мантии, грубые и крупные частицы выпадают из него и оседают на поверхности мантии, а затем удаляются из моллюска. Легко убедиться в наличии интенсивных токов воды, поступающей в мантийную полость, если положить беззубку в неглубокий сосуд с водой так, чтобы вода лишь слегка покрывала раковину. Дав ей успокоиться, надо подсыпать в воду вблизи заднего ее конца какойнибудь порошок, остающийся взвешенным в воде (например, тушь, кармин, сухие тертые водоросли). Тогда можно увидеть, как крупинки порошка уходят через нижний (вводной) сифон внутрь раковины и через некоторое время с сильной струей воды выбрасываются наружу через верхний (выводной) сифон. Время от времени беззубка, часто без всякого внешнего раздражения, с силой захлопывает створки раковины и выбрасывает наружу струи воды, обновляя сразу всю воду, содержащуюся в мантийной полости. Вскоре створки раковины опять приоткрываются и нормальная медленная циркуляция воды возобновляется.Чтобы убедиться в интенсивности работы ресничного эпителия, можно вырезать кусочек мантии беззубки и положить его внутренней поверхностью книзу на дно сосуда. Благодаря продолжающейся еще некоторое время работе ресничек этот кусочек будет слегка перемещаться и даже слегка ползти по наклонной плоскости.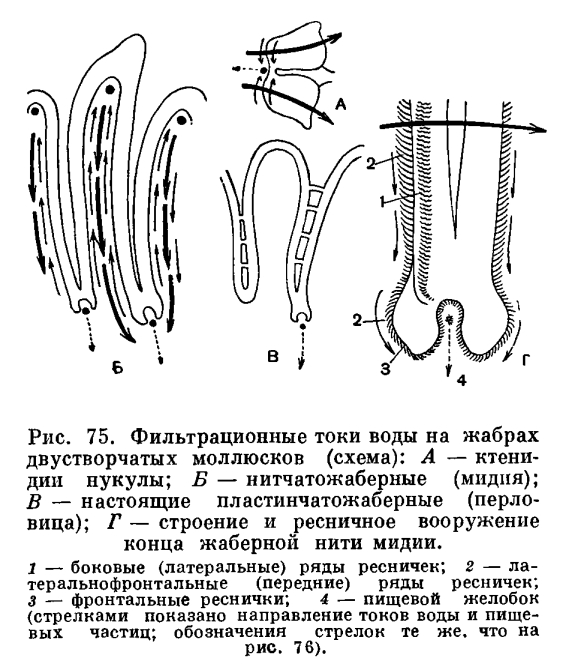 Засасывание воды и ее циркуляция внутри мантийной полости обеспечивают беззубку не только необходимым для ее дыхания кислородом, но и пищей. Как и все двустворчатые моллюски, беззубка лишена головы и целого ряда связанных с ней органов—обособленной глотки, слюнных желез, жестких образований для пережевывания пищи (как, например, хитиновых пластиночек — терки, имеющейся у брюхоногих моллюсков). Поэтому беззубка не может питаться крупными организмами. Она и большинство двустворчатых моллюсков (Eulamellibranchia и Filibranchia) являются активными фильтраторами. Такие моллюски питаются взвешенными в толще воды детритом (мельчайшими остатками отмерших растений и животных) и микропланктоном (одноклеточными водорослями, бактериями и очень мелкими животными). При помощи сложного ресничного механизма жабр и околоротовых лопастей моллюски отфильтровывают их из воды, отделяя несъедобную минеральную взвесь и крупные для них пищевые частицы.
Засасывание воды и ее циркуляция внутри мантийной полости обеспечивают беззубку не только необходимым для ее дыхания кислородом, но и пищей. Как и все двустворчатые моллюски, беззубка лишена головы и целого ряда связанных с ней органов—обособленной глотки, слюнных желез, жестких образований для пережевывания пищи (как, например, хитиновых пластиночек — терки, имеющейся у брюхоногих моллюсков). Поэтому беззубка не может питаться крупными организмами. Она и большинство двустворчатых моллюсков (Eulamellibranchia и Filibranchia) являются активными фильтраторами. Такие моллюски питаются взвешенными в толще воды детритом (мельчайшими остатками отмерших растений и животных) и микропланктоном (одноклеточными водорослями, бактериями и очень мелкими животными). При помощи сложного ресничного механизма жабр и околоротовых лопастей моллюски отфильтровывают их из воды, отделяя несъедобную минеральную взвесь и крупные для них пищевые частицы.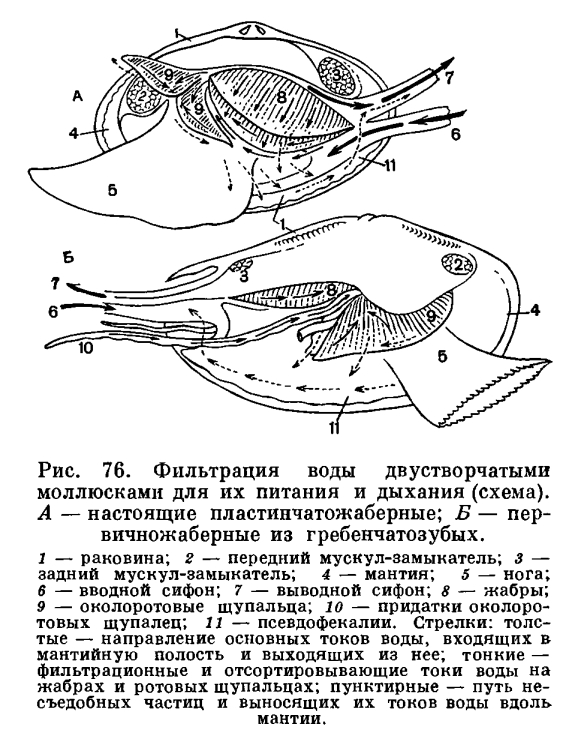 Жаберные нити моллюсков имеют расположенные в определенных местах ряды различных по своим размерам ресничек, которые и могут отфильтровывать и отсортировывать пищевые частицы, обволакивать их слизью, а затем направлять в пищевые бороздки, расположенные вдоль брюшного края полужабр (в местах перехода нисходящих жаберных колен в восходящие) или у их основания. Ряды довольно крупных боковых, наиболее интенсивно работающих ресничек на жаберных нитях производят фильтрацию воды через узкие щели между жаберными нитями и обеспечивают прохождение ее из «вдыхательной» в «выдыхательную» камеру мантийной полости. Особенно крупные латеральнофронтальные реснички, сидящие по бокам жаберных нитей, отцеживают пищевые частички из воды или улавливают их обильно выделяемой слизью и проталкивают на наружную сторону жаберных нитей. Здесь расположены фронтальные реснички, которые собирают пищевые частички и направляют их вниз, к пищевому желобку. Пищевые частицы, собирающиеся в пищевых желобках, также обволакиваются слизью, образуют здесь комочки, уплотняются и благодаря работе ресничек желобка направляются к ротовым лопастям. Ротовые лопасти моллюсков — очень эффективный сортирующий аппарат, освобождающий пищу от несъедобных частиц. Они вооружены многими чувствительными элементами— хемои механорецепторами. Они имеют ряды поперечных бороздок, вооруженных особо длинными ресничками; самые мелкие частицы, пригодные для питания, по ряду таких бороздок направляются к ротовому желобку (расположенному у основания обеих лопастей), по которому они далее направляются к ротовому отверстию, где и заглатываются. По другим бороздкам (с ресничками, работающими в противоположном, чему предшествующих, направлении) более крупные частички и слизистые комочки, непригодные для питания, скатываются вниз и падают на мантию. Сильные реснички мантийных краев гонят эти частицы обратно к основанию вводного сифона; по мере продвижения туда эти частицы склеиваются, уплотняются и в виде так называемых псевдофекалий выбрасываются наружу.У двустворчатых моллюсков из группы Protobranchia (ореховидок, йольдий, портландий и т. п.), имеющих наиболее просто устроенные лепестковидные жабры-к т е н и д и и, ротовые щупальца очень большие, сократимые и снабжены длинным желобчатым выростом. При помощи его они собирают с поверхности грунта мелкие пищевые частички — детрит, которые затем переносятся ресничками по желобку на пластинки ротовых щупалец, где и подвергаются сортировке; жабры-ктенидии служат главным образом для создания токов воды. Работа отфильтровывающего и сортирующего аппарата двустворчатых моллюсков довольно совершенна. Так, мидии могут отфильтровывать частицы размером от 40 до 1, 5—2 мк (лучше всего — в 7—8 мк), полностью извлекая их из воды. Они задерживают одноклеточные водоросли и жгутиковых; более тяжелые частицы минеральных взвесей, даже размером в 4—5 мк, не задерживаются мидиями. Из смеси водорослей и пурпурных бактерий устрицы извлекают только водоросли; они обычно задерживают жгутиковых, водоросли и органические частицы размером более 2—3 мк и пропускают все частицы размером в 1 мк и меньше.Двустворчатые моллюски профильтровывают очень большие объемы воды. Так, устрица может профильтровать за час около 10 л воды; мидия — до 2—5 л (при более высокой температуре воды больше, при более низкой — меньше); съедобная сердцевидка при температуре воды в 17—19, 5°С — от 0, 2 до 2, 5 л, в среднем 0, 5 л воды за час; мелкие морские гребешки фильтруют со скоростью 1 л в час на 1 г их веса, а старые — только 0, 7 л.Пищеварительная система беззубки, как и у всех двустворчатых моллюсков, состоит из короткого пищевода, более или менее округлого желудка, средней и задней кишки; в желудок открываются протоки парной пищеварительной железы — печени, а на брюшной стороне выступает конец так называемого кристаллического стебелька. Отходящий от желудка кишечник (средняя кишка) в основании ноги, в массе половых желез образует 1—2 оборота, затем переходит на спинную сторону и, пронизывая нижнюю стенку перикардиальной сумки, проходит через желудочек сердца, выходит за пределы перикардия через его спинную часть, проходит над задним замыкательным мускулом и оканчивается анальным отверстием, открывающимся в клоакальную камеру мантийной полости с ее выводным сифоном. Часть кишечника, идущая от перикардия до анального отверстия, обычно называется прямой или задней кишкой. Кишечный тракт двустворчатых моллюсков не имеет мышечных волокон, и передвижение пищи в нем происходит благодаря работе выстилающего его ресничного эпителия. Удалению непереваренных остатков способствует мышечная связка, окружающая анальное отверстие.Попав через короткий пищевод в желудок, пищевые частицы благодаря деятельности ресничного тока и желудочного желобка отсортировываются на мелкие и крупные. Крупные пищевые частицы попадают в кишечник, а более мелкие разносятся по складкам желудка и собираются у выступающего конца кристаллического стебелька. Его выступающий конец все время вращается, что способствует перемешиванию пищевых частиц и их сортировке. Кристаллический стебелек образуется в особом мешковидном органе и представляет собой стекловидную палочку студенистого вещества, состоящего из белка типа глобулина с адсорбированными в нем ферментами (амилазой и др.), способными переваривать углеводы (крахмал, гликоген). Попадая в слабокислую среду кишечника, он начинает растворяться и выделять адсорбированные в нем ферменты — единственные выделяемые двустворчатыми моллюсками в кишечный тракт для внеклеточного переваривания пищи. Мелкие пищевые частицы, обработанные энзимами кристаллического стебелька, поступают из желудка в выросты печени. Она состоит из очень большого числа удлиненных слепых трубочек — дивертикул и не является пищеварительной железой в обычном смысле; в ней не вырабатываются и не выделяются в кишечный тракт какие-либо пищеварительные ферменты, и она является органом внутриклеточного (а не внеклеточного) переваривания пищи и всасывания. Внутриклеточное переваривание у двустворчатых моллюсков производится главным образом фагоцитирующими блуждающими клетками — амебоцитами. Они во множестве встречаются не только в дивертикулах печени, но и в желудке и в средней кишке. Амебоциты обладают различными ферментами и способны переваривать не только углеводы, но и белки и жиры и т. д. Блуждающие клетки могут проходить через эпителий кишечного тракта в его просвет и обратно возвращаться в ткани. Заглатывают и переваривают пищевые частицы также и клетки печени; они также могут блуждать по просвету дивертикул и обратно возвращаться в стенки печени. Блуждающим клеткам и при надлежит основная роль в переваривании пищи у двустворчатых моллюсков.При отмирании амебоцитов и клеток печени их пищеварительные ферменты могут поступать в просвет кишечного тракта. Поэтому следы различных ферментов (протеаз, липаз) находят в вытяжках из печени и желудка двустворчатых моллюсков.Далеко не все попавшие в кишечный тракт организмы перевариваются двустворчатыми моллюсками. Нередко, особенно при большом количестве пищи, в фекальных массах моллюсков находят живых диатомей (одноклеточные водоросли с кремниевым скелетом), мелких веслоногих рачков и т. п. Поэтому при искусственном разведении двустворчатых моллюсков (устриц) в выростных бассейнах стараются поддерживать определенную (не избыточную) концентрацию планктонных водорослей, которыми они питаются.Из сказанного видно, что пищеварение у двустворчатых моллюсков очень своеобразно. Они могут переваривать внеклеточно лишь углеводы, а белковые и жировые компоненты их пищи перевариваются фагоцитирующими блуждающими амебоцитами и клетками их «печени». Таким образом, двустворчатые моллюски являются очень специализированной группой животных, приспособленной к питанию детритом, одноклеточными водорослями и бактериями.Кровеносная система у беззубок, как у всех двустворчатых моллюсков, незамкнутая, и кровь — гемолимфа— циркулирует не только по кровеносным сосудам — артериям и венам, но и в пространствах между органами, и в соединительной ткани по целой системе лакун и синусов, не имеющих собственных стенок. Артериальная кровь течет главным образом по сосудам, а венозная система имеет преимущественно лакунарный характер. Кровь прогоняется через всю систему благодаря сокращению сердца, а также мускулатуры тела. Сердце у двустворчатых моллюсков (у анодонты) состоит из желудочка и двух предсердий и лежит в перикардиальной полости, или околосердечной сумке, расположенной на спинной стороне тела. Перикардий представляет собой вытянутый тонкостенный мешочек, наполненный гемолимфой, и является у двустворчатых моллюсков частью сильно уменьшенной в объеме их вторичной полости тела. Желудочек обладает мощными мышечными стенками и имеет вид мешка грушевидной формы, своим широким концом обращенного назад. Предсердия очень тонкостенны, полупрозрачны и чаще всего имеют вид вытянутых треугольников, вершины которых открываются в желудочек; при входе в последний они снабжены небольшими полулунными складками — клапанами, пропускающими кровь только из предсердия в желудочек.У беззубки, как у большинства двустворчатых, желудочек пронизан проходящей через него задней кишкой, но его полость при этом вполне замкнута и отделена от кишки своей стенкой. Из желудочка кровь расходится по телу: к заднему концу — через заднюю аорту, которая разделяется на две артерии, питающие сосуды задней части мантии и заднего замыкательного мускула; к переднему концу — через переднюю аорту и отходящие от нее артерии к ноге, к внутренностям и к передней части мантии. Из артериальных сосудов кровь изливается в не заполненные тканями промежутки, и через систему лакун кровь, ставшая венозной, собирается по синусам и венам в большой продольный венозный синус, лежащий между выделительными органами. Пройдя отсюда через венозную систему почек, она вливается в приносящие парные жаберные артерии, проходящие у основания каждой жабры. Из них венозная кровь по приносящим жаберным сосудам нисходящих жаберных пластинок растекается по жаберным нитям и их сосудистым перемычкам. Окисленная в жабрах артериальная кровь, насыщенная кислородом, по выносящим сосудам восходящих жаберных пластинок вливается в парные (с каждой стороны моллюска) жаберные вены, откуда поступает в предсердия. В предсердия поступает и та часть крови, которая, минуя жабры и почки, окислилась в сосудах мантийных складок и по мантийным венам поступила в наружные жаберяые вены. Мантия с ее сильно развет вленными кровеносными сосудами играет у двустворчатых моллюсков очень большую роль в дыхании и обогащении крови кислородом.То, что у большинства двустворчатых моллюсков желудочек сердца оказывается пронизанным прямой кишкой, находит свое объяснение в особенностях их эмбрионального развития и в эволюции всей этой группы. У ряда низших представителей двустворчатых имеется не только два предсердия, но и два раздельных желудочка, лежащих по бокам кишки (у арок); у других — непарный желудочек лежит над кишкой (ореховидки, аномии, лимы), у третьих — он лежит книзу от кишки (устрицы, жемчужницы и др.)- Все это указывает на то, что расположение кишки и сердца по отношению друг к другу претерпевало большие изменения в процессе эволюции двустворчатых моллюсков, и на то, что первоначально у них было два раздельных желудочка, которые затем слились вместе. Частота сокращения сердца у двустворчатых моллюсков, являющихся в общем малоподвижными организмами, невелика, обычно не более 15—30 раз в минуту, в то время как у таких подвижных и активных моллюсков, как головоногие, сердце сокращается 40—80 раз в минуту. Все отделы сердца двустворчатых моллюсков могут сокращаться автономно.У двустворчатых моллюсков, как и вообще у беспозвоночных с незамкнутой кровеносной системой, кровяное давление очень низкое и сильно изменчиво.Кровь—гемолимфа—двустворчатых моллюсков играет огромную роль в их жизни, обмене веществ. Она выполняет целый ряд функций: обеспечивает внутренние органы и ткани кислородом и питательными веществами, уносит продукты их жизнедеятельности (углекислоту, продукты азотистого обмена и т. п.), создает и поддерживает постоянство их внутренней среды (ионного состава, осмотического давления). Наконец, играет очень большую роль в создании гидравлического механизма давления, необходимого тургора (напряжения), а также при движении моллюсков. Изучение циркуляции крови в теле двустворчатых моллюсков объяснило явление разбухания ноги, что наблюдается при движении и закапывании животного. Оно происходит благодаря наполнению ее кровью, которая и придает ноге необходимую упругость, создает необходимый тургор. Когда нога вытягивается и ножные мышцы расслабляются, кровь поступает по артерии в ногу, а при ее сокращении уходит обратно в тело. Так, у морского черенка, который может очень быстро закапываться, нога сначала погружается в грунт и кровь быстро втекает в нее, расширяя конец ноги в виде диска; последний служит якорем, когда ножные мышцы, сокращаясь, тянут моллюска вниз. При подъеме моллюска из грунта на поверхность ножные мышцы расслабляются, а конец ноги снова расширяется, заполняясь кровью; держась на таком «якоре», нога вытягивается, так как часть крови поступает в верхнюю часть ноги и подталкивает моллюска вверх. Приток, нагнетание необходимого для набухания ноги количества крови и ее отток регулируются так называемым кеберовым органом, играющим при этом как бы роль клапана.В противоположность животным с замкнутой кровеносной системой двустворчатые моллюски, как и все беспозвоночные, обладающие незамкнутой кровеносной системой, имеют довольно значительный объем крови — гемолимфы. У моллюсков (кроме головоногих) он составляет 40—60% (объемных процентов) к весу их тела без раковины. У пресноводной жемчужницы (Margaritifera) и у мидии (Mytilus californianus) на 100 г веса тела объем крови составляет около 50 мл.В крови двустворчатых моллюсков имеется много форменных элементов, главным образом различных форм амебоцитов (лейкоцитов). Количество их колеблется у разных видов от 6000 до 40 000 в 1 мм3. У двустворчатых моллюсков имеются и эритроциты; иногда их даже может быть больше, чем некоторых форм лейкоцитов. Гемоглобин обнаружен у довольно многих видов (арки, морские черенки, теллины, пектункулюсы, астарты и др.).Важная для газообмена (для снабжения органов и тканей кислородом) способность крови насыщаться кислородом у двустворчатых моллюсков очень мала и составляет 1—5% к объему их крови. Так, 100 мл крови беззубки способны поглотить всего лишь 0, 7 мл кислорода, а у мидий — 0, 3 мл. Беззубка потребляет 0, 002 мл 02 на 1 г своего веса за один час (при 10°С); устрицы соответственно — 0, 006 мл 02 (при 20°С); мидии—0, 055 мл 02. Несколько больше обычно потребляют виды более подвижные, как, например, морские гребешки Pecten grandis, потребляющие 0, 07 мл 02 на 1 г своего веса за 1 час (или 70 см3 02 на 1 кг веса). Мелкие формы также нередко потребляют больше кислорода, чем крупные. Например, шаровка роговая при оптимальной для нее температуре воды (18°С) потребляет 0, 05 мг 02 на 1 г веса за час, но при снижении температуры воды до 0, 5°С потребление кислорода у нее почти прекращается. В потреблении кислорода, т. е. в интенсивности обмена, у многих двустворчатых моллюсков наблюдаются сезонные колебания; так, мидии в летнее, наиболее активное время их жизни потребляют примерно в два раза больше кислорода, чем в зимнее, холодное время года.Многие двустворчатые моллюски могут довольно долгое время жить при очень малом содержании кислорода в воде или даже при его отсутствии. Так, песчаная ракушка (Муа arenaria) может жить в бескислородных условиях при 14°С до 8 дней, а при 0° даже несколько недель; виргинская устрица также выносит такие условия в течение недели и более. Обмен веществ в такие периоды анаэробиоза резко снижается, но моллюски могут при этом получать необходимый для их жизни кислород путем интрамолекулярного дыхания — гликолитического расщепления их запасных веществ (углеводов и жиров) по типу брожения. Такая способность ко временному (факультативному) анаэробиозу (аноксибиозу) особенно характерна и необходима для видов, живущих на литорали (как, например, для песчаной ракушки, мидий, балтийской макомы, съедобной сердцевидки). Во время отлива они закрывают свои раковины, небольшое количество кислорода в их мантийной полости довольно быстро исчезает, и они начинают жить за счет процессов аноксибиоза. Во время прилива они раскрывают свои раковины, непрерывно фильтруют воду и дышат кислородом, растворенным в воде; вначале у них резко (в несколько раз) повышается интенсивность фильтрации и потребления кислорода, а затем через некоторое время приходит в норму, характерную для их жизни в воде.Органами выделения у двустворчатых моллюсков являются почки, а также, но в меньшей степени так называемый кеберов орган; последний представляет собой железистое утолщение передней части и передне-боковых стенок перикардиальной сумки.Почки, или боянусовы органы, открываются своими внутренними концами в перикардий, а наружными — в мантийную полость. Почки у беззубки имеют вид двух темно-зеленых изогнутых трубчатых мешочков; один конец имеет железистые стенки и представляет собственно выделительную часть почки, другой имеет вид пузырька, где накапливаются продукты обмена веществ, подлежащие удалению из тела.У двустворчатых моллюсков нет такой концентрации частей центральной нервной системы (нервных узелков, или ганглиев), как у брюхоногих моллюсков. У беззубки, например, имеется одна пара ганглиев над ртом, немного позади него, другая пара — глубоко в ноге и третья — позади заднего замыкательного мускула. Между первой и второй парой ганглиев, а также между первой и третьей проходит по паре нервных стволов, а каждая пара узелков соединена между собой поперечными перемычками (комиссурами).Органы чувств у двустворчатых по сравнению с другими классами моллюсков развиты довольно слабо. Однако эти органы у них довольно разнообразны по своему строению и рассеяны в различных частях тела: вдоль наружных краев мантии, на концах сифонов, на первых жаберных нитях, около ротового отверстия на околоротовых щупальцах, на краях заднего замыкательного мускула, в выдыхательной камере, около задней кишки и т. д. Эти органы чувств представляют собой как довольно сложные образования — глаза, или фоторецепторы, органы равновесия — статоцисты, или статорецепторы, так и более простые — осфрадии, различные чувствительные выросты, а иногда и просто скопления пигментированных чувствительных клеток.Фоторецепторы у двустворчатых моллюсков могут быть устроены очень различно: от простых эпителиальных пигментных (оптических органелл) до довольно сложных глаз с хрусталиком и сетчаткой. Такие глаза могут быть очень многочисленными, особенно у свободноживущих форм, как например мантийные глаза у гребешков, у которых их насчитывается иногда до 100 на обоих краях мантии.Различно устроенные глаза и глазки могут располагаться у двустворчатых также на первых жаберных нитях (жаберные глаза у арок, аномий), на коротких выростах вокруг отверстий сифонов (у некоторых сердцевидок и др.).У многих двустворчатых имеются так называемые оптические органеллы, сферические или удлиненные, концентрирующие свет на особом внутриклеточном иннервированном образовании (ретинелле). Такие фоторецепторы разбросаны на концах сифонов и в других частях тела моллюсков.Органы равновесия у двустворчатых представляют собой пузыревидное впячивание эпителия, хорошо иннервируемое, выстланное изнутри ресничным эпителием, замкнутое (статоцист) или открытое (статокрипт). Они содержат внутри твердое минеральное зерно (статолит) или мелкие песчинки (статоконии). Обычно, например у беззубки, статоцисты расположены недалеко от ножного ганглия или на спинной стороне моллюска. Органы равновесия хорошо развиты у свободноживущих форм, например у морских гребешков.Осфрадии представляют собой обычно очень мелкие парные пигментированные хорошо иннервируемые группы чувствительных клеток. Они могут располагаться в различных местах — на ноге, в области жабр, задней кишки и т. д. Их роль еще недостаточно ясна: являются ли они хеморецепторами или органами осязания.
Жаберные нити моллюсков имеют расположенные в определенных местах ряды различных по своим размерам ресничек, которые и могут отфильтровывать и отсортировывать пищевые частицы, обволакивать их слизью, а затем направлять в пищевые бороздки, расположенные вдоль брюшного края полужабр (в местах перехода нисходящих жаберных колен в восходящие) или у их основания. Ряды довольно крупных боковых, наиболее интенсивно работающих ресничек на жаберных нитях производят фильтрацию воды через узкие щели между жаберными нитями и обеспечивают прохождение ее из «вдыхательной» в «выдыхательную» камеру мантийной полости. Особенно крупные латеральнофронтальные реснички, сидящие по бокам жаберных нитей, отцеживают пищевые частички из воды или улавливают их обильно выделяемой слизью и проталкивают на наружную сторону жаберных нитей. Здесь расположены фронтальные реснички, которые собирают пищевые частички и направляют их вниз, к пищевому желобку. Пищевые частицы, собирающиеся в пищевых желобках, также обволакиваются слизью, образуют здесь комочки, уплотняются и благодаря работе ресничек желобка направляются к ротовым лопастям. Ротовые лопасти моллюсков — очень эффективный сортирующий аппарат, освобождающий пищу от несъедобных частиц. Они вооружены многими чувствительными элементами— хемои механорецепторами. Они имеют ряды поперечных бороздок, вооруженных особо длинными ресничками; самые мелкие частицы, пригодные для питания, по ряду таких бороздок направляются к ротовому желобку (расположенному у основания обеих лопастей), по которому они далее направляются к ротовому отверстию, где и заглатываются. По другим бороздкам (с ресничками, работающими в противоположном, чему предшествующих, направлении) более крупные частички и слизистые комочки, непригодные для питания, скатываются вниз и падают на мантию. Сильные реснички мантийных краев гонят эти частицы обратно к основанию вводного сифона; по мере продвижения туда эти частицы склеиваются, уплотняются и в виде так называемых псевдофекалий выбрасываются наружу.У двустворчатых моллюсков из группы Protobranchia (ореховидок, йольдий, портландий и т. п.), имеющих наиболее просто устроенные лепестковидные жабры-к т е н и д и и, ротовые щупальца очень большие, сократимые и снабжены длинным желобчатым выростом. При помощи его они собирают с поверхности грунта мелкие пищевые частички — детрит, которые затем переносятся ресничками по желобку на пластинки ротовых щупалец, где и подвергаются сортировке; жабры-ктенидии служат главным образом для создания токов воды. Работа отфильтровывающего и сортирующего аппарата двустворчатых моллюсков довольно совершенна. Так, мидии могут отфильтровывать частицы размером от 40 до 1, 5—2 мк (лучше всего — в 7—8 мк), полностью извлекая их из воды. Они задерживают одноклеточные водоросли и жгутиковых; более тяжелые частицы минеральных взвесей, даже размером в 4—5 мк, не задерживаются мидиями. Из смеси водорослей и пурпурных бактерий устрицы извлекают только водоросли; они обычно задерживают жгутиковых, водоросли и органические частицы размером более 2—3 мк и пропускают все частицы размером в 1 мк и меньше.Двустворчатые моллюски профильтровывают очень большие объемы воды. Так, устрица может профильтровать за час около 10 л воды; мидия — до 2—5 л (при более высокой температуре воды больше, при более низкой — меньше); съедобная сердцевидка при температуре воды в 17—19, 5°С — от 0, 2 до 2, 5 л, в среднем 0, 5 л воды за час; мелкие морские гребешки фильтруют со скоростью 1 л в час на 1 г их веса, а старые — только 0, 7 л.Пищеварительная система беззубки, как и у всех двустворчатых моллюсков, состоит из короткого пищевода, более или менее округлого желудка, средней и задней кишки; в желудок открываются протоки парной пищеварительной железы — печени, а на брюшной стороне выступает конец так называемого кристаллического стебелька. Отходящий от желудка кишечник (средняя кишка) в основании ноги, в массе половых желез образует 1—2 оборота, затем переходит на спинную сторону и, пронизывая нижнюю стенку перикардиальной сумки, проходит через желудочек сердца, выходит за пределы перикардия через его спинную часть, проходит над задним замыкательным мускулом и оканчивается анальным отверстием, открывающимся в клоакальную камеру мантийной полости с ее выводным сифоном. Часть кишечника, идущая от перикардия до анального отверстия, обычно называется прямой или задней кишкой. Кишечный тракт двустворчатых моллюсков не имеет мышечных волокон, и передвижение пищи в нем происходит благодаря работе выстилающего его ресничного эпителия. Удалению непереваренных остатков способствует мышечная связка, окружающая анальное отверстие.Попав через короткий пищевод в желудок, пищевые частицы благодаря деятельности ресничного тока и желудочного желобка отсортировываются на мелкие и крупные. Крупные пищевые частицы попадают в кишечник, а более мелкие разносятся по складкам желудка и собираются у выступающего конца кристаллического стебелька. Его выступающий конец все время вращается, что способствует перемешиванию пищевых частиц и их сортировке. Кристаллический стебелек образуется в особом мешковидном органе и представляет собой стекловидную палочку студенистого вещества, состоящего из белка типа глобулина с адсорбированными в нем ферментами (амилазой и др.), способными переваривать углеводы (крахмал, гликоген). Попадая в слабокислую среду кишечника, он начинает растворяться и выделять адсорбированные в нем ферменты — единственные выделяемые двустворчатыми моллюсками в кишечный тракт для внеклеточного переваривания пищи. Мелкие пищевые частицы, обработанные энзимами кристаллического стебелька, поступают из желудка в выросты печени. Она состоит из очень большого числа удлиненных слепых трубочек — дивертикул и не является пищеварительной железой в обычном смысле; в ней не вырабатываются и не выделяются в кишечный тракт какие-либо пищеварительные ферменты, и она является органом внутриклеточного (а не внеклеточного) переваривания пищи и всасывания. Внутриклеточное переваривание у двустворчатых моллюсков производится главным образом фагоцитирующими блуждающими клетками — амебоцитами. Они во множестве встречаются не только в дивертикулах печени, но и в желудке и в средней кишке. Амебоциты обладают различными ферментами и способны переваривать не только углеводы, но и белки и жиры и т. д. Блуждающие клетки могут проходить через эпителий кишечного тракта в его просвет и обратно возвращаться в ткани. Заглатывают и переваривают пищевые частицы также и клетки печени; они также могут блуждать по просвету дивертикул и обратно возвращаться в стенки печени. Блуждающим клеткам и при надлежит основная роль в переваривании пищи у двустворчатых моллюсков.При отмирании амебоцитов и клеток печени их пищеварительные ферменты могут поступать в просвет кишечного тракта. Поэтому следы различных ферментов (протеаз, липаз) находят в вытяжках из печени и желудка двустворчатых моллюсков.Далеко не все попавшие в кишечный тракт организмы перевариваются двустворчатыми моллюсками. Нередко, особенно при большом количестве пищи, в фекальных массах моллюсков находят живых диатомей (одноклеточные водоросли с кремниевым скелетом), мелких веслоногих рачков и т. п. Поэтому при искусственном разведении двустворчатых моллюсков (устриц) в выростных бассейнах стараются поддерживать определенную (не избыточную) концентрацию планктонных водорослей, которыми они питаются.Из сказанного видно, что пищеварение у двустворчатых моллюсков очень своеобразно. Они могут переваривать внеклеточно лишь углеводы, а белковые и жировые компоненты их пищи перевариваются фагоцитирующими блуждающими амебоцитами и клетками их «печени». Таким образом, двустворчатые моллюски являются очень специализированной группой животных, приспособленной к питанию детритом, одноклеточными водорослями и бактериями.Кровеносная система у беззубок, как у всех двустворчатых моллюсков, незамкнутая, и кровь — гемолимфа— циркулирует не только по кровеносным сосудам — артериям и венам, но и в пространствах между органами, и в соединительной ткани по целой системе лакун и синусов, не имеющих собственных стенок. Артериальная кровь течет главным образом по сосудам, а венозная система имеет преимущественно лакунарный характер. Кровь прогоняется через всю систему благодаря сокращению сердца, а также мускулатуры тела. Сердце у двустворчатых моллюсков (у анодонты) состоит из желудочка и двух предсердий и лежит в перикардиальной полости, или околосердечной сумке, расположенной на спинной стороне тела. Перикардий представляет собой вытянутый тонкостенный мешочек, наполненный гемолимфой, и является у двустворчатых моллюсков частью сильно уменьшенной в объеме их вторичной полости тела. Желудочек обладает мощными мышечными стенками и имеет вид мешка грушевидной формы, своим широким концом обращенного назад. Предсердия очень тонкостенны, полупрозрачны и чаще всего имеют вид вытянутых треугольников, вершины которых открываются в желудочек; при входе в последний они снабжены небольшими полулунными складками — клапанами, пропускающими кровь только из предсердия в желудочек.У беззубки, как у большинства двустворчатых, желудочек пронизан проходящей через него задней кишкой, но его полость при этом вполне замкнута и отделена от кишки своей стенкой. Из желудочка кровь расходится по телу: к заднему концу — через заднюю аорту, которая разделяется на две артерии, питающие сосуды задней части мантии и заднего замыкательного мускула; к переднему концу — через переднюю аорту и отходящие от нее артерии к ноге, к внутренностям и к передней части мантии. Из артериальных сосудов кровь изливается в не заполненные тканями промежутки, и через систему лакун кровь, ставшая венозной, собирается по синусам и венам в большой продольный венозный синус, лежащий между выделительными органами. Пройдя отсюда через венозную систему почек, она вливается в приносящие парные жаберные артерии, проходящие у основания каждой жабры. Из них венозная кровь по приносящим жаберным сосудам нисходящих жаберных пластинок растекается по жаберным нитям и их сосудистым перемычкам. Окисленная в жабрах артериальная кровь, насыщенная кислородом, по выносящим сосудам восходящих жаберных пластинок вливается в парные (с каждой стороны моллюска) жаберные вены, откуда поступает в предсердия. В предсердия поступает и та часть крови, которая, минуя жабры и почки, окислилась в сосудах мантийных складок и по мантийным венам поступила в наружные жаберяые вены. Мантия с ее сильно развет вленными кровеносными сосудами играет у двустворчатых моллюсков очень большую роль в дыхании и обогащении крови кислородом.То, что у большинства двустворчатых моллюсков желудочек сердца оказывается пронизанным прямой кишкой, находит свое объяснение в особенностях их эмбрионального развития и в эволюции всей этой группы. У ряда низших представителей двустворчатых имеется не только два предсердия, но и два раздельных желудочка, лежащих по бокам кишки (у арок); у других — непарный желудочек лежит над кишкой (ореховидки, аномии, лимы), у третьих — он лежит книзу от кишки (устрицы, жемчужницы и др.)- Все это указывает на то, что расположение кишки и сердца по отношению друг к другу претерпевало большие изменения в процессе эволюции двустворчатых моллюсков, и на то, что первоначально у них было два раздельных желудочка, которые затем слились вместе. Частота сокращения сердца у двустворчатых моллюсков, являющихся в общем малоподвижными организмами, невелика, обычно не более 15—30 раз в минуту, в то время как у таких подвижных и активных моллюсков, как головоногие, сердце сокращается 40—80 раз в минуту. Все отделы сердца двустворчатых моллюсков могут сокращаться автономно.У двустворчатых моллюсков, как и вообще у беспозвоночных с незамкнутой кровеносной системой, кровяное давление очень низкое и сильно изменчиво.Кровь—гемолимфа—двустворчатых моллюсков играет огромную роль в их жизни, обмене веществ. Она выполняет целый ряд функций: обеспечивает внутренние органы и ткани кислородом и питательными веществами, уносит продукты их жизнедеятельности (углекислоту, продукты азотистого обмена и т. п.), создает и поддерживает постоянство их внутренней среды (ионного состава, осмотического давления). Наконец, играет очень большую роль в создании гидравлического механизма давления, необходимого тургора (напряжения), а также при движении моллюсков. Изучение циркуляции крови в теле двустворчатых моллюсков объяснило явление разбухания ноги, что наблюдается при движении и закапывании животного. Оно происходит благодаря наполнению ее кровью, которая и придает ноге необходимую упругость, создает необходимый тургор. Когда нога вытягивается и ножные мышцы расслабляются, кровь поступает по артерии в ногу, а при ее сокращении уходит обратно в тело. Так, у морского черенка, который может очень быстро закапываться, нога сначала погружается в грунт и кровь быстро втекает в нее, расширяя конец ноги в виде диска; последний служит якорем, когда ножные мышцы, сокращаясь, тянут моллюска вниз. При подъеме моллюска из грунта на поверхность ножные мышцы расслабляются, а конец ноги снова расширяется, заполняясь кровью; держась на таком «якоре», нога вытягивается, так как часть крови поступает в верхнюю часть ноги и подталкивает моллюска вверх. Приток, нагнетание необходимого для набухания ноги количества крови и ее отток регулируются так называемым кеберовым органом, играющим при этом как бы роль клапана.В противоположность животным с замкнутой кровеносной системой двустворчатые моллюски, как и все беспозвоночные, обладающие незамкнутой кровеносной системой, имеют довольно значительный объем крови — гемолимфы. У моллюсков (кроме головоногих) он составляет 40—60% (объемных процентов) к весу их тела без раковины. У пресноводной жемчужницы (Margaritifera) и у мидии (Mytilus californianus) на 100 г веса тела объем крови составляет около 50 мл.В крови двустворчатых моллюсков имеется много форменных элементов, главным образом различных форм амебоцитов (лейкоцитов). Количество их колеблется у разных видов от 6000 до 40 000 в 1 мм3. У двустворчатых моллюсков имеются и эритроциты; иногда их даже может быть больше, чем некоторых форм лейкоцитов. Гемоглобин обнаружен у довольно многих видов (арки, морские черенки, теллины, пектункулюсы, астарты и др.).Важная для газообмена (для снабжения органов и тканей кислородом) способность крови насыщаться кислородом у двустворчатых моллюсков очень мала и составляет 1—5% к объему их крови. Так, 100 мл крови беззубки способны поглотить всего лишь 0, 7 мл кислорода, а у мидий — 0, 3 мл. Беззубка потребляет 0, 002 мл 02 на 1 г своего веса за один час (при 10°С); устрицы соответственно — 0, 006 мл 02 (при 20°С); мидии—0, 055 мл 02. Несколько больше обычно потребляют виды более подвижные, как, например, морские гребешки Pecten grandis, потребляющие 0, 07 мл 02 на 1 г своего веса за 1 час (или 70 см3 02 на 1 кг веса). Мелкие формы также нередко потребляют больше кислорода, чем крупные. Например, шаровка роговая при оптимальной для нее температуре воды (18°С) потребляет 0, 05 мг 02 на 1 г веса за час, но при снижении температуры воды до 0, 5°С потребление кислорода у нее почти прекращается. В потреблении кислорода, т. е. в интенсивности обмена, у многих двустворчатых моллюсков наблюдаются сезонные колебания; так, мидии в летнее, наиболее активное время их жизни потребляют примерно в два раза больше кислорода, чем в зимнее, холодное время года.Многие двустворчатые моллюски могут довольно долгое время жить при очень малом содержании кислорода в воде или даже при его отсутствии. Так, песчаная ракушка (Муа arenaria) может жить в бескислородных условиях при 14°С до 8 дней, а при 0° даже несколько недель; виргинская устрица также выносит такие условия в течение недели и более. Обмен веществ в такие периоды анаэробиоза резко снижается, но моллюски могут при этом получать необходимый для их жизни кислород путем интрамолекулярного дыхания — гликолитического расщепления их запасных веществ (углеводов и жиров) по типу брожения. Такая способность ко временному (факультативному) анаэробиозу (аноксибиозу) особенно характерна и необходима для видов, живущих на литорали (как, например, для песчаной ракушки, мидий, балтийской макомы, съедобной сердцевидки). Во время отлива они закрывают свои раковины, небольшое количество кислорода в их мантийной полости довольно быстро исчезает, и они начинают жить за счет процессов аноксибиоза. Во время прилива они раскрывают свои раковины, непрерывно фильтруют воду и дышат кислородом, растворенным в воде; вначале у них резко (в несколько раз) повышается интенсивность фильтрации и потребления кислорода, а затем через некоторое время приходит в норму, характерную для их жизни в воде.Органами выделения у двустворчатых моллюсков являются почки, а также, но в меньшей степени так называемый кеберов орган; последний представляет собой железистое утолщение передней части и передне-боковых стенок перикардиальной сумки.Почки, или боянусовы органы, открываются своими внутренними концами в перикардий, а наружными — в мантийную полость. Почки у беззубки имеют вид двух темно-зеленых изогнутых трубчатых мешочков; один конец имеет железистые стенки и представляет собственно выделительную часть почки, другой имеет вид пузырька, где накапливаются продукты обмена веществ, подлежащие удалению из тела.У двустворчатых моллюсков нет такой концентрации частей центральной нервной системы (нервных узелков, или ганглиев), как у брюхоногих моллюсков. У беззубки, например, имеется одна пара ганглиев над ртом, немного позади него, другая пара — глубоко в ноге и третья — позади заднего замыкательного мускула. Между первой и второй парой ганглиев, а также между первой и третьей проходит по паре нервных стволов, а каждая пара узелков соединена между собой поперечными перемычками (комиссурами).Органы чувств у двустворчатых по сравнению с другими классами моллюсков развиты довольно слабо. Однако эти органы у них довольно разнообразны по своему строению и рассеяны в различных частях тела: вдоль наружных краев мантии, на концах сифонов, на первых жаберных нитях, около ротового отверстия на околоротовых щупальцах, на краях заднего замыкательного мускула, в выдыхательной камере, около задней кишки и т. д. Эти органы чувств представляют собой как довольно сложные образования — глаза, или фоторецепторы, органы равновесия — статоцисты, или статорецепторы, так и более простые — осфрадии, различные чувствительные выросты, а иногда и просто скопления пигментированных чувствительных клеток.Фоторецепторы у двустворчатых моллюсков могут быть устроены очень различно: от простых эпителиальных пигментных (оптических органелл) до довольно сложных глаз с хрусталиком и сетчаткой. Такие глаза могут быть очень многочисленными, особенно у свободноживущих форм, как например мантийные глаза у гребешков, у которых их насчитывается иногда до 100 на обоих краях мантии.Различно устроенные глаза и глазки могут располагаться у двустворчатых также на первых жаберных нитях (жаберные глаза у арок, аномий), на коротких выростах вокруг отверстий сифонов (у некоторых сердцевидок и др.).У многих двустворчатых имеются так называемые оптические органеллы, сферические или удлиненные, концентрирующие свет на особом внутриклеточном иннервированном образовании (ретинелле). Такие фоторецепторы разбросаны на концах сифонов и в других частях тела моллюсков.Органы равновесия у двустворчатых представляют собой пузыревидное впячивание эпителия, хорошо иннервируемое, выстланное изнутри ресничным эпителием, замкнутое (статоцист) или открытое (статокрипт). Они содержат внутри твердое минеральное зерно (статолит) или мелкие песчинки (статоконии). Обычно, например у беззубки, статоцисты расположены недалеко от ножного ганглия или на спинной стороне моллюска. Органы равновесия хорошо развиты у свободноживущих форм, например у морских гребешков.Осфрадии представляют собой обычно очень мелкие парные пигментированные хорошо иннервируемые группы чувствительных клеток. Они могут располагаться в различных местах — на ноге, в области жабр, задней кишки и т. д. Их роль еще недостаточно ясна: являются ли они хеморецепторами или органами осязания. Анодонты, как и большинство двустворчатых, раздельнополы, но в условиях водоемов со стоячей водой могут встречаться отдельные гермафродитные особи или даже целые их колонии. При этом во избежание самооплодотворения сначала продуцируются мужские половые продукты — сперматозоиды, а затем женские — яйца. Парные, сильно расчлененные половые железы (гонады) у двустворчатых (в том числе и у анодонты) залегают в спинной части ноги, где их окружает петля кишечника и выросты печени; выводные протоки гонад открываются в мантийную полость рядом с отверстиями выделительной системы. Только у некоторых самых примитивных двустворчатых моллюсков протоки половых желез открываются общим отверстием с выделительной порой. У некоторых пресноводных двустворчатых половой диморфизм выражен настолько резко, что самцов и самок одного и того же вида иногда относят к разным видам.
Анодонты, как и большинство двустворчатых, раздельнополы, но в условиях водоемов со стоячей водой могут встречаться отдельные гермафродитные особи или даже целые их колонии. При этом во избежание самооплодотворения сначала продуцируются мужские половые продукты — сперматозоиды, а затем женские — яйца. Парные, сильно расчлененные половые железы (гонады) у двустворчатых (в том числе и у анодонты) залегают в спинной части ноги, где их окружает петля кишечника и выросты печени; выводные протоки гонад открываются в мантийную полость рядом с отверстиями выделительной системы. Только у некоторых самых примитивных двустворчатых моллюсков протоки половых желез открываются общим отверстием с выделительной порой. У некоторых пресноводных двустворчатых половой диморфизм выражен настолько резко, что самцов и самок одного и того же вида иногда относят к разным видам. Развитие молоди у двустворчатых моллюсков происходит разнообразно. Почти все морские формы, обитающие на мелководьях, откладывают яйца прямо в воду, где и происходит оплодотворение, или же оно происходит в мантийной полости материнской особи. Яйца свободно плавают в воде, реже склеиваются или прикрепляются к раковинам, водорослям. Исключение представляют живородящие (точнее, личинкородящие) формы, как некоторые устрицы, арки и др.Оплодотворенные яйца двустворчатых моллюсков, пройдя стадию дробления спирального типа, образуют трохофорообразную личинку, сходную с личинкой многощетинковых червей (полихет). Однако в процессе эмбрионального развития двустворчатых моллюсков почти отсутствует процесс сегментации, столь характерный для развития личиночных стадий кольчатых червей. У личинки двустворчатых моллюсков имеется зачаток ноги и первичной раковины (продиссоконха), которая первоначально закладывается в виде одной пластинки, расположенной на спинной стороне личинки. После ряда изменений трохофора, у которой возникает парус (velum) — ресничный теменной диск, двустворчатая раковинка и зачатки других органов, превращаются в велигер. Наличие свободноплавающей личинки (велигер) представляет собой очень важную жизненную стадию моллюсков, так как обеспечивает им возможности широкого расселения, поскольку взрослые двустворчатые моллюски ведут обычно малоподвижный или даже прикрепленный образ жизни. Вместе с тем эта личиночная стадия жизни, как и у других беспозвоночных, является самой чувствительной к неблагоприятным внешним условиям, и лишь высокая плодовитость двустворчатых моллюсков обеспечивает сохранение вида и его расселение.У некоторых видов двустворчатых эта пелагическая личиночная стадия утрачена и яйца развиваются внутри мантийной полости материнского организма (как у многих пресноводных форм), так что из материнской раковины выходят уже почти готовые молодые особи. У некоторых пресноводных же форм цикл развития осложняется своеобразным паразитизмом личинок на теле рыб. Этот период следует за периодом первоначального развития на жабрах материнской особи, куда и откладываются их яйца, при этом у некоторых видов на жабрах образуются специальные выводковые камеры (марзупиум). Примером может служить развитие личинки анодонты —глохидия, протекающее, как и у всех Unionidae, в наружных жабрах материнской особи. В дальнейшем он становится паразитом рыб, прикрепляясь к их коже или к жабрам биссусом или зубцами раковинки. Погружаясь в ткани рыбы, глохидий окружается цистой, питается за счет соков тела хозяина. Этот паразитический период продолжается 1—2 месяца, после чего моллюски выходят в воду и, опускаясь на дно, переходят к свободному образу жизни.У ряда морских холодноводных и, видимо, у многих глубоководных океанических видов двустворчатых моллюсков развитие может происходить с очень укороченной стадией плавающей личинки или совсем без нее. В последнем случае образуются немногочисленные крупные яйца, снабженные большим количеством питательного вещества. Это позволяет им развиваться вне зависимости от наличия пищи в окружающей воде, что особенно важно для глубоководных форм, где количество пищи для молоди у дна очень ограниченно.Основываясь на строении раковины, замка, жабр, на расположении и количестве мускулов-замыкателей, связки и пр., различают отряды: Гребенчатозубые, Связочнозубые, Настоящие пластинчатожаберные и Перегородчатожаберные.
Развитие молоди у двустворчатых моллюсков происходит разнообразно. Почти все морские формы, обитающие на мелководьях, откладывают яйца прямо в воду, где и происходит оплодотворение, или же оно происходит в мантийной полости материнской особи. Яйца свободно плавают в воде, реже склеиваются или прикрепляются к раковинам, водорослям. Исключение представляют живородящие (точнее, личинкородящие) формы, как некоторые устрицы, арки и др.Оплодотворенные яйца двустворчатых моллюсков, пройдя стадию дробления спирального типа, образуют трохофорообразную личинку, сходную с личинкой многощетинковых червей (полихет). Однако в процессе эмбрионального развития двустворчатых моллюсков почти отсутствует процесс сегментации, столь характерный для развития личиночных стадий кольчатых червей. У личинки двустворчатых моллюсков имеется зачаток ноги и первичной раковины (продиссоконха), которая первоначально закладывается в виде одной пластинки, расположенной на спинной стороне личинки. После ряда изменений трохофора, у которой возникает парус (velum) — ресничный теменной диск, двустворчатая раковинка и зачатки других органов, превращаются в велигер. Наличие свободноплавающей личинки (велигер) представляет собой очень важную жизненную стадию моллюсков, так как обеспечивает им возможности широкого расселения, поскольку взрослые двустворчатые моллюски ведут обычно малоподвижный или даже прикрепленный образ жизни. Вместе с тем эта личиночная стадия жизни, как и у других беспозвоночных, является самой чувствительной к неблагоприятным внешним условиям, и лишь высокая плодовитость двустворчатых моллюсков обеспечивает сохранение вида и его расселение.У некоторых видов двустворчатых эта пелагическая личиночная стадия утрачена и яйца развиваются внутри мантийной полости материнского организма (как у многих пресноводных форм), так что из материнской раковины выходят уже почти готовые молодые особи. У некоторых пресноводных же форм цикл развития осложняется своеобразным паразитизмом личинок на теле рыб. Этот период следует за периодом первоначального развития на жабрах материнской особи, куда и откладываются их яйца, при этом у некоторых видов на жабрах образуются специальные выводковые камеры (марзупиум). Примером может служить развитие личинки анодонты —глохидия, протекающее, как и у всех Unionidae, в наружных жабрах материнской особи. В дальнейшем он становится паразитом рыб, прикрепляясь к их коже или к жабрам биссусом или зубцами раковинки. Погружаясь в ткани рыбы, глохидий окружается цистой, питается за счет соков тела хозяина. Этот паразитический период продолжается 1—2 месяца, после чего моллюски выходят в воду и, опускаясь на дно, переходят к свободному образу жизни.У ряда морских холодноводных и, видимо, у многих глубоководных океанических видов двустворчатых моллюсков развитие может происходить с очень укороченной стадией плавающей личинки или совсем без нее. В последнем случае образуются немногочисленные крупные яйца, снабженные большим количеством питательного вещества. Это позволяет им развиваться вне зависимости от наличия пищи в окружающей воде, что особенно важно для глубоководных форм, где количество пищи для молоди у дна очень ограниченно.Основываясь на строении раковины, замка, жабр, на расположении и количестве мускулов-замыкателей, связки и пр., различают отряды: Гребенчатозубые, Связочнозубые, Настоящие пластинчатожаберные и Перегородчатожаберные.
Жизнь животных: в 6-ти томах. — М.: Просвещение. Под редакцией профессоров Н.А.Гладкова, А.В.Михеева. 1970.
.