- Общий очерк3
- Общий очерк3
-
Краткая характеристика. Земноводные занимают особое место среди других животных, так как представляют собой первых и наиболее просто организованных наземных позвоночных.Как обитатели суши, земноводные дышат легкими, имеют два круга кровообращения, трехкамерное сердце; у них намечается разделение крови на артериальную и венозную. Передвигаются земноводные при помощи конечностей пятипалого типа с шарообразными суставами. Череп сочленяется с позвоночником подвижно (двумя мыщелками).Первичная верхняя челюсть (нёбно-квадратный хрящ) срастается с черепной коробкой (аутостилия), и подвесок (гиомандибуляре) превращается в слуховую косточку. Строение органа слуха более совершенно, чем у рыб: кроме внутреннего уха, имеется еще и среднее. Глаза приспособлены к видению на большом расстоянии. В крыше переднего мозга образуется первичный мозговой свод (архипаллиум).Несовершенство приспособлений к жизни на суше сказывается у амфибий во всех системах органов. Температура их тела зависит от температуры и влажности окружающей среды; они отличаются ограниченными возможностями распространения, передвижения и ориентации на суше. Примитивность амфибий как наземных животных особенно ярко выражена в том, что их яйца лишены оболочек, защищающих их от высыхания, и, как правило, не могут развиваться вне воды. В связи с этим у земноводных развивается личинка, обитающая в воде. Развитие протекает с превращением (метаморфоз), в результате которого водная личинка превращается в животное, обитающее на суше.Земноводные — самый малочисленный класс позвоночных, включающий лишь около 2100 современных видов, представленных тремя отрядами: хвостатых, безногих и бесхвостых.Бесхвостые земноводные (Anura) содержат наибольшее число — около 1800 — видов, приспособившихся к передвижению по суше прыжками при помощи удлиненных задних конечностей. Сюда относятся различные лягушки, жабы, чесночницы, жерлянки, квакши и т.п. Распространены бесхвостые по всем материкам, кроме Антарктиды.Хвостатые земноводные (Caudata, или Urodela) более примитивны; их всего около 280 видов. К ним относятся всевозможные саламандры и тритоны, распространенные почти исключительно в северном полушарии.Безногие (Apoda) объединяют примерно 55 видов тропических червяг, большинство из которых ведет подземный образ жизни. По-видимому, это очень древние земноводные, дожившие до наших дней благодаря приспособленности к роющему образу жизни.СТРОЕНИЕ И ЖИЗНЕДЕЯТЕЛЬНОСТЬ ОРГАНИЗМА ЗЕМНОВОДНЫХУсловия обитания в воде и на суше резко отличны, и организация амфибий глубочайшим образом отличается от организации их водных предков.Влажность, постоянная для водной среды, на суше резко изменяется, и на большей части земной поверхности она относительно мала. Поскольку газообмен между организмом и средой возможен только через водную пленку, жабры, легко высыхающие на воздухе, оказываются непригодными как органы дыхания для наземных животных. Они заменяются у амфибий легкими. В отличие от жабр легкие расположены глубоко внутри тела и защищены от высыхания.Легкие земноводных представляют собой парные мешки, полые внутри. Их тонкие стенки на внутренней поверхности имеют более или менее развитое ячеистое строение. Однако поверхность легких у земноводных еще невелика. Так, у большинства лягушек отношение ее к поверхности кожи равно 2 : 3, в то время как у млекопитающих поверхность легкого в 50—100 раз больше поверхности кожи. Незначительное развитие легких у земноводных характеризует их как примитивных обитателей суши.
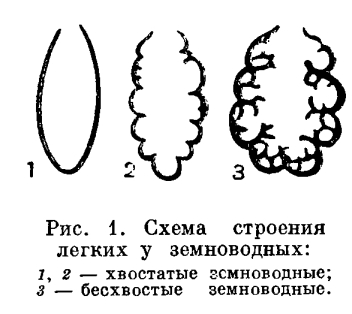 Дыхательные пути у амфибий развиты также слабо. У хвостатых они представлены в виде довольно длинной трубки — трахеи; у бесхвостых это лишь короткая трахейно-гортанная камера, непосредственно переходящая в полость легких.Соответственно преобразованию органов дыхания у наземных позвоночных животных изменяется и механизм дыхания. У земноводных он еще примитивного нагнетательного типа. Животное набирает воздух в ротовую полость, для чего открывает ноздри и опускает дно ротовой полости. Затем ноздри закрываются клапанами, дно ротовой полости поднимается и воздух нагнетается в легкие. Удаление воздуха из легких происходит благодаря действию брюшной мускулатуры и спадению стенок легких. Механизм дыхания отличает земноводных не только от рыб, но и от других наземных позвоночных, дыхание которых осуществляется при помощи изменения объема грудной клетки.
Дыхательные пути у амфибий развиты также слабо. У хвостатых они представлены в виде довольно длинной трубки — трахеи; у бесхвостых это лишь короткая трахейно-гортанная камера, непосредственно переходящая в полость легких.Соответственно преобразованию органов дыхания у наземных позвоночных животных изменяется и механизм дыхания. У земноводных он еще примитивного нагнетательного типа. Животное набирает воздух в ротовую полость, для чего открывает ноздри и опускает дно ротовой полости. Затем ноздри закрываются клапанами, дно ротовой полости поднимается и воздух нагнетается в легкие. Удаление воздуха из легких происходит благодаря действию брюшной мускулатуры и спадению стенок легких. Механизм дыхания отличает земноводных не только от рыб, но и от других наземных позвоночных, дыхание которых осуществляется при помощи изменения объема грудной клетки. Так как у земноводных относительная поверхность легких мала и они слабо вентилируются, то насыщение крови кислородом происходит не только через легкие, но и через кожу. Так, например, травяная лягушка получает через кожу 33% кислорода, а прудовая — 51%. Еще сложнее обстоит дело с удалением из организма углекислого газа. Благодаря недостаточной вентиляции легких углекислый газ скопляется в них и затрудняет дальнейшую диффузию этого вещества из крови. Основная масса углекислого газа диффундирует в окружающую среду через кожу (у прудовой лягушки 86%). У высших наземных позвоночных кожное дыхание ничтожно мало.Покровы земноводных, функционирующие как дополнительный орган дыхания, отличаются в связи с этим рядом особенностей. Кожа у земноводных голая, что способствует свободному газообмену в кровеносных сосудах, образующих в ней густую сеть. Так как обмен газами между организмом и средой идет только через водную пленку, кожа земноводных постоянно увлажняется слизью, вырабатываемой многочисленными железами. Кроме слизистых, в коже земноводных помещаются еще и ядовитые железы, секрет которых защищает животное от врагов и различных микроорганизмов, для которых влажная кожа земноводных могла бы служить отличным белковым субстратом.
Так как у земноводных относительная поверхность легких мала и они слабо вентилируются, то насыщение крови кислородом происходит не только через легкие, но и через кожу. Так, например, травяная лягушка получает через кожу 33% кислорода, а прудовая — 51%. Еще сложнее обстоит дело с удалением из организма углекислого газа. Благодаря недостаточной вентиляции легких углекислый газ скопляется в них и затрудняет дальнейшую диффузию этого вещества из крови. Основная масса углекислого газа диффундирует в окружающую среду через кожу (у прудовой лягушки 86%). У высших наземных позвоночных кожное дыхание ничтожно мало.Покровы земноводных, функционирующие как дополнительный орган дыхания, отличаются в связи с этим рядом особенностей. Кожа у земноводных голая, что способствует свободному газообмену в кровеносных сосудах, образующих в ней густую сеть. Так как обмен газами между организмом и средой идет только через водную пленку, кожа земноводных постоянно увлажняется слизью, вырабатываемой многочисленными железами. Кроме слизистых, в коже земноводных помещаются еще и ядовитые железы, секрет которых защищает животное от врагов и различных микроорганизмов, для которых влажная кожа земноводных могла бы служить отличным белковым субстратом.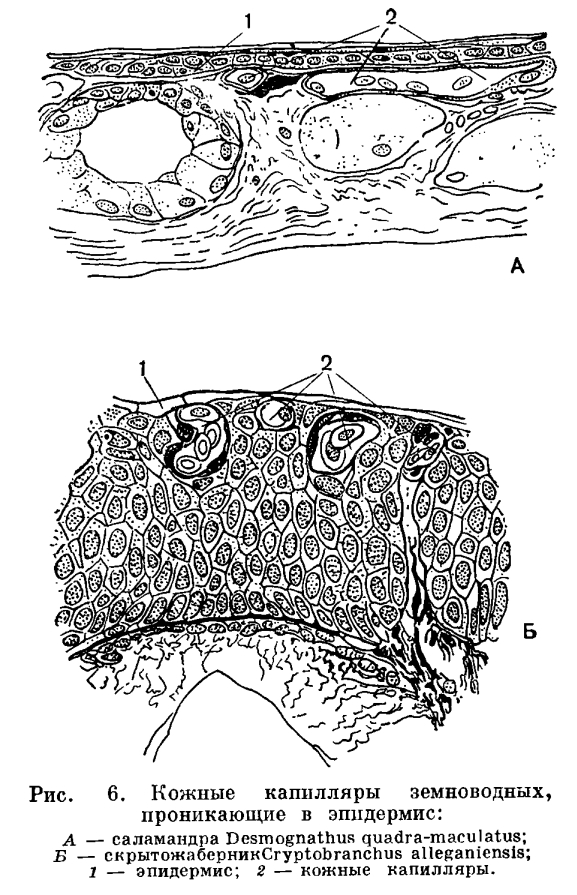 С развитием наземного органа дыхания тесно связана и перестройка системы кровообращения. Сердце земноводных состоит из двух вполне обособленных предсердий, общего желудочка и артериального конуса, от которого отходит общий ствол аорты, разделяющийся затем на три пары артериальных сосудов. Передние из них — сонные артерии — несут кровь к голове. Следующие за ними сосуды носят название системных дуг аорты. Правая и левая дуги аорты, отсылая каждая по мощной артерии к передним конечностям, соединяются ниже сердца в непарную спинную аорту. Последняя тянется вдоль позвоночника, отсылая от себя артерии к различным органам. От общего ствола аорты отходят также легочно-кожные артерии, несущие кровь к легким и коже.
С развитием наземного органа дыхания тесно связана и перестройка системы кровообращения. Сердце земноводных состоит из двух вполне обособленных предсердий, общего желудочка и артериального конуса, от которого отходит общий ствол аорты, разделяющийся затем на три пары артериальных сосудов. Передние из них — сонные артерии — несут кровь к голове. Следующие за ними сосуды носят название системных дуг аорты. Правая и левая дуги аорты, отсылая каждая по мощной артерии к передним конечностям, соединяются ниже сердца в непарную спинную аорту. Последняя тянется вдоль позвоночника, отсылая от себя артерии к различным органам. От общего ствола аорты отходят также легочно-кожные артерии, несущие кровь к легким и коже.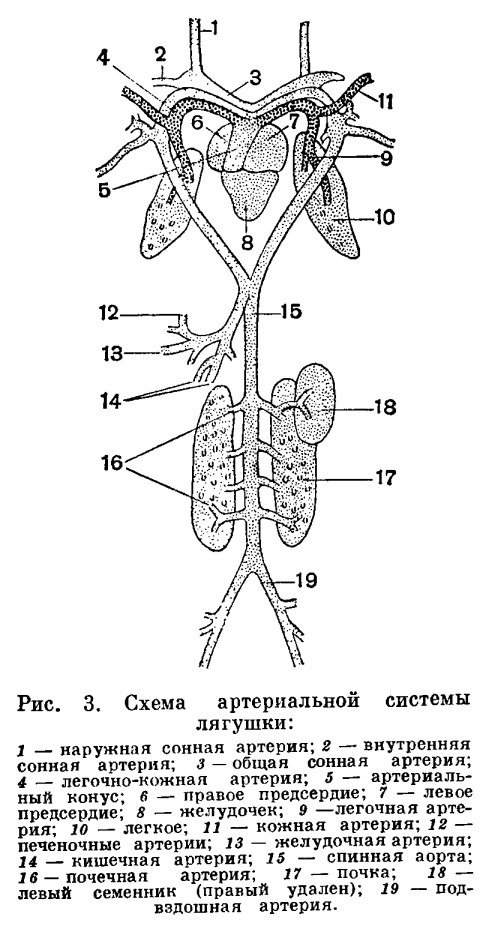 Венозная кровь от передних отделов тела собирается в парные передние полые вены, куда открываются также очень характерные для земноводных большие кожные вены, несущие артериальную кровь от кожи. Передние полые вены, как и непарная задняя полая вена, впадают в правое предсердие. В заднюю полую вену собирается кровь из задних отделов тела. Она принимает в себя и печеночную вену, собирающую кровь от кишечника. Наконец, в левое предсердие впадает общая легочная вена, образующаяся при слиянии парных легочных вен.
Венозная кровь от передних отделов тела собирается в парные передние полые вены, куда открываются также очень характерные для земноводных большие кожные вены, несущие артериальную кровь от кожи. Передние полые вены, как и непарная задняя полая вена, впадают в правое предсердие. В заднюю полую вену собирается кровь из задних отделов тела. Она принимает в себя и печеночную вену, собирающую кровь от кишечника. Наконец, в левое предсердие впадает общая легочная вена, образующаяся при слиянии парных легочных вен.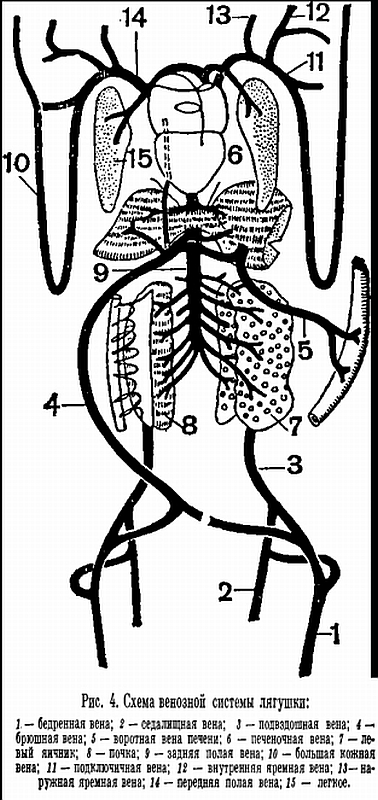 Следовательно, в отличие от рыб, у земноводных возникает типичный для всех наземных позвоночных второй круг кровообращения, по которому кровь из сердца по легочным артериям поступает в легкие и возвращается в него по легочной вене. При одном круге кровообращения у рыб в сердце попадает только венозная кровь, а у наземных позвоночных и венозная и артериальная. В связи с этим возникает разделение сердца на два отдела: правый — венозный и левый — артериальный. У земноводных это разделение лишь частичное и выражается в существовании двух предсердий. Однако уже в правом предсердии кровь смешивается, так как верхние полые вены приносят в него не только венозную кровь, но и идущую по кожным венам артериальную. В желудочке к этой смешанной крови добавляется еще порция артериальной крови из левого предсердия. Разделение желудочка при таком кровообращении сделало бы бесцельным кожное дыхание, так как артериальная кровь из кожных вен поступала бы тогда только в легкие. Отсутствие перегородки в желудочке приобретает исключительно важную роль тогда, когда животное находится под водой и дышит только кожей.
Следовательно, в отличие от рыб, у земноводных возникает типичный для всех наземных позвоночных второй круг кровообращения, по которому кровь из сердца по легочным артериям поступает в легкие и возвращается в него по легочной вене. При одном круге кровообращения у рыб в сердце попадает только венозная кровь, а у наземных позвоночных и венозная и артериальная. В связи с этим возникает разделение сердца на два отдела: правый — венозный и левый — артериальный. У земноводных это разделение лишь частичное и выражается в существовании двух предсердий. Однако уже в правом предсердии кровь смешивается, так как верхние полые вены приносят в него не только венозную кровь, но и идущую по кожным венам артериальную. В желудочке к этой смешанной крови добавляется еще порция артериальной крови из левого предсердия. Разделение желудочка при таком кровообращении сделало бы бесцельным кожное дыхание, так как артериальная кровь из кожных вен поступала бы тогда только в легкие. Отсутствие перегородки в желудочке приобретает исключительно важную роль тогда, когда животное находится под водой и дышит только кожей.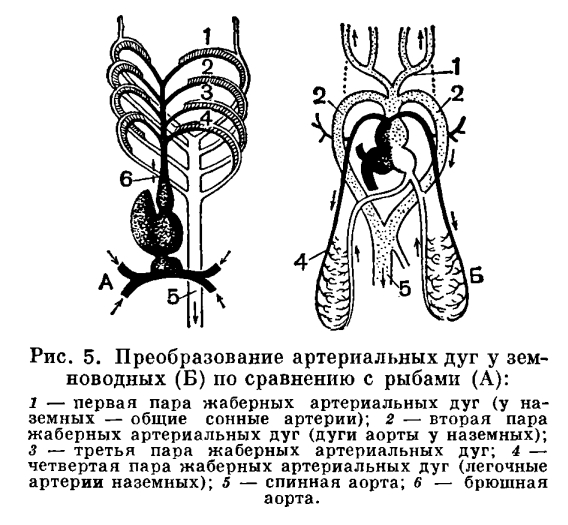 Не менее существенное преобразование в кровеносной системе земноводных состоит в том, что в связи с исчезновением жаберного дыхания артерии жаберных дуг изменяются и приобретают новую функцию. Как показывает эмбриональное развитие земноводных, их легочные артерии формируются за счет артерий четвертой жаберной дуги, артерия третьей жаберной дуги у взрослых бесхвостых земноводных исчезает, из второй — развиваются дуги аорты, а из первой — сонные артерии.Передние и задние кардинальные вены, свойственные рыбам, остаются еще у некоторых хвостатых земноводных, а у бесхвостых их целиком заменяют типичные для наземных позвоночных передняя и задняя полые вены.С примитивностью органов дыхания земноводных теснейшим образом связаны и особенности строения их органов выделения. Голая, ничем не защищенная кожа земноводных не способна, как у других наземных позвоночных, противостоять поступлению в организм воды по законам осмотического давления. Подобно рыбам, они «пьют» непроизвольно, всей поверхностью тела. Избыток воды удаляется из организма органами выделения. Они представлены, как и у рыб, двумя туловищными (мезонефрическими) почками, каждая из которых состоит из клубочков и канальцев. В клубочках происходит фильтрация мочи из крови. По мере прохождения ее по канальцам часть веществ, в том числе и вода, всасывается обратно. Моча в окончательном виде из канальцев попадает в мочеточники (вольфовы каналы) и выделяется наружу. Отличительная особенность туловищной почки состоит в том, что клубочки здесь велики, а канальцы коротки. Такое строение почки приспособлено к выведению из организма большого количества воды.Следовательно, особенности строения легких определяют у земноводных своеобразие строения кожи, выполняющей роль важного дополнительного органа дыхания. Последнее, в свою очередь, определяет тип органов выделения, остающихся на уровне развития, свойственном рыбам. Однако избыточное поступление воды в организм не грозило бы земноводным, если бы они не собирались для размножения в водоемы. Иными словами, характер органов выделения находится в связи и с особенностями размножения.Температурные условия обитания в воде и на суше так же различны, как и условия влажности. Колебания температуры в воде в различные сезоны года не превышают 25—30°, а на суше они достигают 120°. При этом здесь очень велика не только сезонная, но и суточная амплитуда, доходящая, если учитывать температуру поверхности почвы, до 60°.В связи с громадным разнообразием температур в окружающей среде весьма важным приспособлением для наземных животных оказывается способность поддерживать температуру тела на постоянном уровне. Однако и в этом отношении земноводные характеризуются как примитивные обитатели суши, относящиеся, подобно рыбам, к хладнокровным (пойкилотермным) животным.Слабое развитие легких и снабжение клеток и тканей смешанной, недостаточно богатой кислородом кровью приводит к тому, что обмен веществ у земноводных находится на низком уровне и в теле продуцируется мало тепла. Кроме того, кожа земноводных, функционирующая как дополнительный орган дыхания, лишена защитных образований, способных удерживать тепло в теле. Постоянное испарение влаги через кожу в связи с ее дыхательной функцией значительно снижает температуру тела. При низкой влажности воздуха температура тела благодаря испарению с его поверхности может оказаться ниже температуры окружающей среды почти на 10°.Сохранение у земноводных хладнокровности есть следствие особенностей строения их органов дыхания и кровообращения, а также кожных покровов.Выход позвоночных в новую среду обитания сопровождался не только изменением влажности и температуры, т. е. изменением факторов неживой природы, но и изменением взаимоотношений с другими живыми организмами, прежде всего теми, которые служат пищей. Первые наземные позвоночные, видимо, имели достаточно корма, и стимул к дальнейшему прогрессивному развитию пищеварительного тракта, обеспечивающему максимальное переваривание и усвоение пищи, у них отсутствовал. Кроме того, при низком уровне окислительных процессов, непостоянной температуре тела и незначительной подвижности животного потребность в пище у земноводных невелика. Дифференциация пищевдрительного тракта у этих животных осталась примерно на том же уровне, что и у их предков — рыб. Земноводные имеют общую ро-тоглоточную полость, короткий пищевод, переходящий в сравнительно слабо обособленный желудок. Желудок без резкой границы переходит в кишечник, в котором хорошо отграничена только задняя (прямая) кишка, открывающаяся в клоаку. Большая печень снабжена желчным пузырем; поджелудочная железа расположена, как всегда, в первой петле кишечника. Зубы всех современных земноводных, если они у них есть, имеют вид простых конусов, приращенных основанием к кости, а вершиной направленных назад. Все зубы однородны и служат лишь для удержания добычи, которая заглатывается целиком. Они очень малы, по мере снашивания выпадают и заменяются другими. Сидят зубы не только на челюстях, но и на сошниках. Присутствие сошнико вых зубов характерно только для рыб и земноводных.
Не менее существенное преобразование в кровеносной системе земноводных состоит в том, что в связи с исчезновением жаберного дыхания артерии жаберных дуг изменяются и приобретают новую функцию. Как показывает эмбриональное развитие земноводных, их легочные артерии формируются за счет артерий четвертой жаберной дуги, артерия третьей жаберной дуги у взрослых бесхвостых земноводных исчезает, из второй — развиваются дуги аорты, а из первой — сонные артерии.Передние и задние кардинальные вены, свойственные рыбам, остаются еще у некоторых хвостатых земноводных, а у бесхвостых их целиком заменяют типичные для наземных позвоночных передняя и задняя полые вены.С примитивностью органов дыхания земноводных теснейшим образом связаны и особенности строения их органов выделения. Голая, ничем не защищенная кожа земноводных не способна, как у других наземных позвоночных, противостоять поступлению в организм воды по законам осмотического давления. Подобно рыбам, они «пьют» непроизвольно, всей поверхностью тела. Избыток воды удаляется из организма органами выделения. Они представлены, как и у рыб, двумя туловищными (мезонефрическими) почками, каждая из которых состоит из клубочков и канальцев. В клубочках происходит фильтрация мочи из крови. По мере прохождения ее по канальцам часть веществ, в том числе и вода, всасывается обратно. Моча в окончательном виде из канальцев попадает в мочеточники (вольфовы каналы) и выделяется наружу. Отличительная особенность туловищной почки состоит в том, что клубочки здесь велики, а канальцы коротки. Такое строение почки приспособлено к выведению из организма большого количества воды.Следовательно, особенности строения легких определяют у земноводных своеобразие строения кожи, выполняющей роль важного дополнительного органа дыхания. Последнее, в свою очередь, определяет тип органов выделения, остающихся на уровне развития, свойственном рыбам. Однако избыточное поступление воды в организм не грозило бы земноводным, если бы они не собирались для размножения в водоемы. Иными словами, характер органов выделения находится в связи и с особенностями размножения.Температурные условия обитания в воде и на суше так же различны, как и условия влажности. Колебания температуры в воде в различные сезоны года не превышают 25—30°, а на суше они достигают 120°. При этом здесь очень велика не только сезонная, но и суточная амплитуда, доходящая, если учитывать температуру поверхности почвы, до 60°.В связи с громадным разнообразием температур в окружающей среде весьма важным приспособлением для наземных животных оказывается способность поддерживать температуру тела на постоянном уровне. Однако и в этом отношении земноводные характеризуются как примитивные обитатели суши, относящиеся, подобно рыбам, к хладнокровным (пойкилотермным) животным.Слабое развитие легких и снабжение клеток и тканей смешанной, недостаточно богатой кислородом кровью приводит к тому, что обмен веществ у земноводных находится на низком уровне и в теле продуцируется мало тепла. Кроме того, кожа земноводных, функционирующая как дополнительный орган дыхания, лишена защитных образований, способных удерживать тепло в теле. Постоянное испарение влаги через кожу в связи с ее дыхательной функцией значительно снижает температуру тела. При низкой влажности воздуха температура тела благодаря испарению с его поверхности может оказаться ниже температуры окружающей среды почти на 10°.Сохранение у земноводных хладнокровности есть следствие особенностей строения их органов дыхания и кровообращения, а также кожных покровов.Выход позвоночных в новую среду обитания сопровождался не только изменением влажности и температуры, т. е. изменением факторов неживой природы, но и изменением взаимоотношений с другими живыми организмами, прежде всего теми, которые служат пищей. Первые наземные позвоночные, видимо, имели достаточно корма, и стимул к дальнейшему прогрессивному развитию пищеварительного тракта, обеспечивающему максимальное переваривание и усвоение пищи, у них отсутствовал. Кроме того, при низком уровне окислительных процессов, непостоянной температуре тела и незначительной подвижности животного потребность в пище у земноводных невелика. Дифференциация пищевдрительного тракта у этих животных осталась примерно на том же уровне, что и у их предков — рыб. Земноводные имеют общую ро-тоглоточную полость, короткий пищевод, переходящий в сравнительно слабо обособленный желудок. Желудок без резкой границы переходит в кишечник, в котором хорошо отграничена только задняя (прямая) кишка, открывающаяся в клоаку. Большая печень снабжена желчным пузырем; поджелудочная железа расположена, как всегда, в первой петле кишечника. Зубы всех современных земноводных, если они у них есть, имеют вид простых конусов, приращенных основанием к кости, а вершиной направленных назад. Все зубы однородны и служат лишь для удержания добычи, которая заглатывается целиком. Они очень малы, по мере снашивания выпадают и заменяются другими. Сидят зубы не только на челюстях, но и на сошниках. Присутствие сошнико вых зубов характерно только для рыб и земноводных. В ротоглоточную полость у амфибий открываются отсутствующие у рыб слюнные железы, секрет которых смачивает ротовую полость и пищу, но не действует на последнюю химически. Появление слюнных желез типично для наземных позвоночных и служит приспособлением против иссушающего действия воздушной среды.По сравнению с рыбами у земноводных прогрессивно развивается язык и, обладая собственной мускулатурой, принимает участие в захватывании пищи.
В ротоглоточную полость у амфибий открываются отсутствующие у рыб слюнные железы, секрет которых смачивает ротовую полость и пищу, но не действует на последнюю химически. Появление слюнных желез типично для наземных позвоночных и служит приспособлением против иссушающего действия воздушной среды.По сравнению с рыбами у земноводных прогрессивно развивается язык и, обладая собственной мускулатурой, принимает участие в захватывании пищи.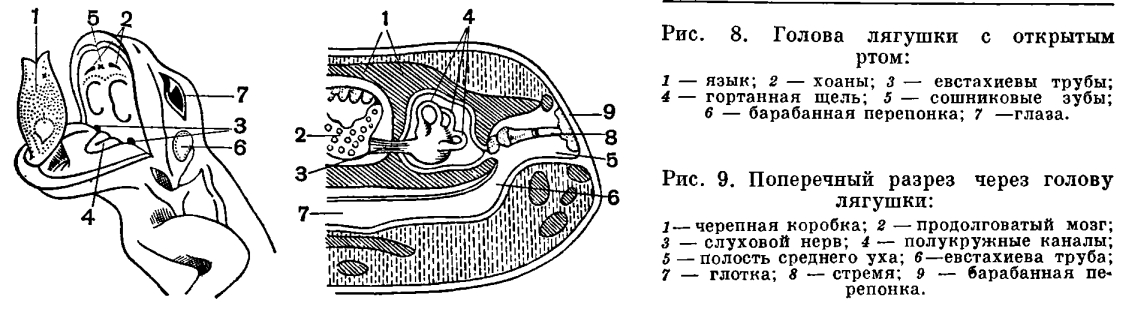 С выходом на сушу перестраиваются не только те системы органов, которые непосредственно обеспечивают обмен веществ в организме, но и органы чувств, и центральная нервная система.В водной среде меньше света, и сравнительно малая прозрачность воды не позволяет видеть далеко. В связи с этим рыбы близоруки. Воздушная среда не препятствует видимости, и земноводные приспособлены к рассматриванию предметов на более или менее далеком расстоянии. Эти приспособления выражены преимущественно в выпуклой форме роговицы и в форме хрусталика, имеющего вид двояковыпуклой линзы. Аккомодация у амфибий, так же как и у рыб, достигается лишь перемещением хрусталика, однако не при помощи серповидного отростка, а путем сокращения ресничшй мышцы, характерной для наземных позвоночных.В связи с обитанием на суше у земноводных имеются подвижные веки, защищающие глаза от высыхания и засорения.Кроме внутреннего уха, имевшегося уже у рыб, у амфибий развито среднее ухо, представляющее собой видоизмененную полость брызгальца рыб, отграниченную от окружающей среды барабанной перепонкой. В этой полости помещается впервые появившаяся у позвоночных слуховая косточка — столбик, представляющая собой видоизмененную подвесочную кость. Одним концом столбик упирается в барабанную перепонку, другим — в истонченный участок перегородки, разделяющей среднее и внутреннее ухо. Столбик передает колебания барабанной перепонки внутреннему уху. Полость среднего уха соединяется с ротовой полостью посредством узкого канала — евстахиевой трубыи оказывается сообщающейся с наружной средой, благодаря чему внутреннее и внешнее давление на барабанную перепонку уравновешивается. Это предохраняет перепонку от разрыва.Усложнение органов обоняния у земноводных выражается в изменении формы складчатости, увеличивающей обонятельную поверхность. У рыб и личинок земноводных эти складки мелкие и расположены радиально. У взрослых земноводных они крупнее и имеют вид более или менее закрученных горизонтальных выростов. Кроме того, у земноводных начинает обособляться часть обонятельного мешка, образующая самостоятельный отдел органа обоняния, характерный для наземных позвоночных — Якобсонов орган, по-видимому, служащий для обоняния пищи, находящейся во рту. Наконец, в стенках обонятельного органа земноводных, как и у всех наземных позвоночных, развиваются многочисленные железы, увлажняющие слизистую оболочку.Орган обоняния не только расположен вблизи ротовой полости, но и соединен с ней при помощи внутренних ноздрей, или хоан. Это включает орган обоняния в состав проводящих дыхательных путей. Весь воздух, используемый при дыхании, проходит через орган обоняния. Акт вдоха служит земноводным и для восприятия запахов.Приспособление к обмену воздуха в органе обоняния, с одной стороны, ведет к усложнению пути воздуха к органу дыхания. С другой, стороны, орган обоняния, приобретая новую функцию, начинает подразделяться на собственно обонятельную область, выстланную обонятельным эпителием, и дыхательную область, выстланную простым эпителием.Прогрессивное развитие всех органов чувств у земноводных в известной мере объясняется и тем, что органы боковой линии у них во взрослом состоянии, как правило, исчезают. Для водных обитателей органы боковой линии имеют главенствующее значение и до некоторой степени компенсируют функцию других органов чувств.Перестройка органов чувств, их усложнение и совершенствование, приводящее к более дифференцированной и тонкой ориентации в окружающей среде, связано с прогрессивными изменениями в головном мозге. Головной мозг земноводных отличается от мозга рыб главным образом большим развитием переднего мозга, который полностью разделен на два полушария. Развитие переднего мозга выражено также и в том, что он содержит большое число нервных клеток. Не только дно желудочков переднего мозга, но и их бока и крыша содержат нервное вещество, т. е. у земноводных появился уже первичный мозговой свод — архипаллиу м. Однако серое вещество содержится еще только в глубинных слоях крыши и отсутствует на ее поверхности.Мозжечок у земноводных недоразвит в связи с малой подвижностью и однообразным характером движений этих животных.
С выходом на сушу перестраиваются не только те системы органов, которые непосредственно обеспечивают обмен веществ в организме, но и органы чувств, и центральная нервная система.В водной среде меньше света, и сравнительно малая прозрачность воды не позволяет видеть далеко. В связи с этим рыбы близоруки. Воздушная среда не препятствует видимости, и земноводные приспособлены к рассматриванию предметов на более или менее далеком расстоянии. Эти приспособления выражены преимущественно в выпуклой форме роговицы и в форме хрусталика, имеющего вид двояковыпуклой линзы. Аккомодация у амфибий, так же как и у рыб, достигается лишь перемещением хрусталика, однако не при помощи серповидного отростка, а путем сокращения ресничшй мышцы, характерной для наземных позвоночных.В связи с обитанием на суше у земноводных имеются подвижные веки, защищающие глаза от высыхания и засорения.Кроме внутреннего уха, имевшегося уже у рыб, у амфибий развито среднее ухо, представляющее собой видоизмененную полость брызгальца рыб, отграниченную от окружающей среды барабанной перепонкой. В этой полости помещается впервые появившаяся у позвоночных слуховая косточка — столбик, представляющая собой видоизмененную подвесочную кость. Одним концом столбик упирается в барабанную перепонку, другим — в истонченный участок перегородки, разделяющей среднее и внутреннее ухо. Столбик передает колебания барабанной перепонки внутреннему уху. Полость среднего уха соединяется с ротовой полостью посредством узкого канала — евстахиевой трубыи оказывается сообщающейся с наружной средой, благодаря чему внутреннее и внешнее давление на барабанную перепонку уравновешивается. Это предохраняет перепонку от разрыва.Усложнение органов обоняния у земноводных выражается в изменении формы складчатости, увеличивающей обонятельную поверхность. У рыб и личинок земноводных эти складки мелкие и расположены радиально. У взрослых земноводных они крупнее и имеют вид более или менее закрученных горизонтальных выростов. Кроме того, у земноводных начинает обособляться часть обонятельного мешка, образующая самостоятельный отдел органа обоняния, характерный для наземных позвоночных — Якобсонов орган, по-видимому, служащий для обоняния пищи, находящейся во рту. Наконец, в стенках обонятельного органа земноводных, как и у всех наземных позвоночных, развиваются многочисленные железы, увлажняющие слизистую оболочку.Орган обоняния не только расположен вблизи ротовой полости, но и соединен с ней при помощи внутренних ноздрей, или хоан. Это включает орган обоняния в состав проводящих дыхательных путей. Весь воздух, используемый при дыхании, проходит через орган обоняния. Акт вдоха служит земноводным и для восприятия запахов.Приспособление к обмену воздуха в органе обоняния, с одной стороны, ведет к усложнению пути воздуха к органу дыхания. С другой, стороны, орган обоняния, приобретая новую функцию, начинает подразделяться на собственно обонятельную область, выстланную обонятельным эпителием, и дыхательную область, выстланную простым эпителием.Прогрессивное развитие всех органов чувств у земноводных в известной мере объясняется и тем, что органы боковой линии у них во взрослом состоянии, как правило, исчезают. Для водных обитателей органы боковой линии имеют главенствующее значение и до некоторой степени компенсируют функцию других органов чувств.Перестройка органов чувств, их усложнение и совершенствование, приводящее к более дифференцированной и тонкой ориентации в окружающей среде, связано с прогрессивными изменениями в головном мозге. Головной мозг земноводных отличается от мозга рыб главным образом большим развитием переднего мозга, который полностью разделен на два полушария. Развитие переднего мозга выражено также и в том, что он содержит большое число нервных клеток. Не только дно желудочков переднего мозга, но и их бока и крыша содержат нервное вещество, т. е. у земноводных появился уже первичный мозговой свод — архипаллиу м. Однако серое вещество содержится еще только в глубинных слоях крыши и отсутствует на ее поверхности.Мозжечок у земноводных недоразвит в связи с малой подвижностью и однообразным характером движений этих животных.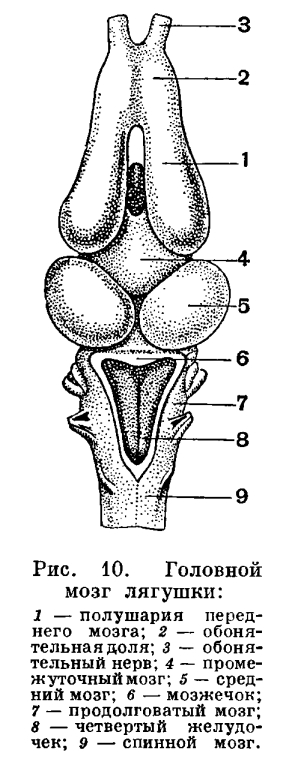 Жизнь на суше меняет характер передвижения. Изменения эти связаны с тем, что в водной среде не требуется почти никаких усилий для поддержания тела, в то время как с выходом на сушу эти усилия возрастают, поскольку удельный вес тела увеличивается во много раз. Удельный вес рыбы, имеющей плавательный пузырь, равен единице. При выходе на сушу ее удельный вес оказывается во много раз больше удельного веса воздуха. Кроме того, условия передвижения на суше и в воде отличны, потому что земля не представляет собой сколько-нибудь ровной поверхности. Органами передвижения земноводных служат передние и задние конечности пятипалого типа, характерного для всех наземных позвоночных. Принципиальное отличие в строении наземных конечностей от строения плавников заключается в том, что они удлинены и расчленены на подвижно соединенные между собой отделы. Иными словами, скелет их устроен по принципу системы рычагов, и это уменьшает усилия, необходимые для поддержания и передвижения тела в пространстве. В связи с частными приспособлениями к тому или иному типу передвижения у разных представителей земноводных строение конечностей видоизменяется.
Жизнь на суше меняет характер передвижения. Изменения эти связаны с тем, что в водной среде не требуется почти никаких усилий для поддержания тела, в то время как с выходом на сушу эти усилия возрастают, поскольку удельный вес тела увеличивается во много раз. Удельный вес рыбы, имеющей плавательный пузырь, равен единице. При выходе на сушу ее удельный вес оказывается во много раз больше удельного веса воздуха. Кроме того, условия передвижения на суше и в воде отличны, потому что земля не представляет собой сколько-нибудь ровной поверхности. Органами передвижения земноводных служат передние и задние конечности пятипалого типа, характерного для всех наземных позвоночных. Принципиальное отличие в строении наземных конечностей от строения плавников заключается в том, что они удлинены и расчленены на подвижно соединенные между собой отделы. Иными словами, скелет их устроен по принципу системы рычагов, и это уменьшает усилия, необходимые для поддержания и передвижения тела в пространстве. В связи с частными приспособлениями к тому или иному типу передвижения у разных представителей земноводных строение конечностей видоизменяется.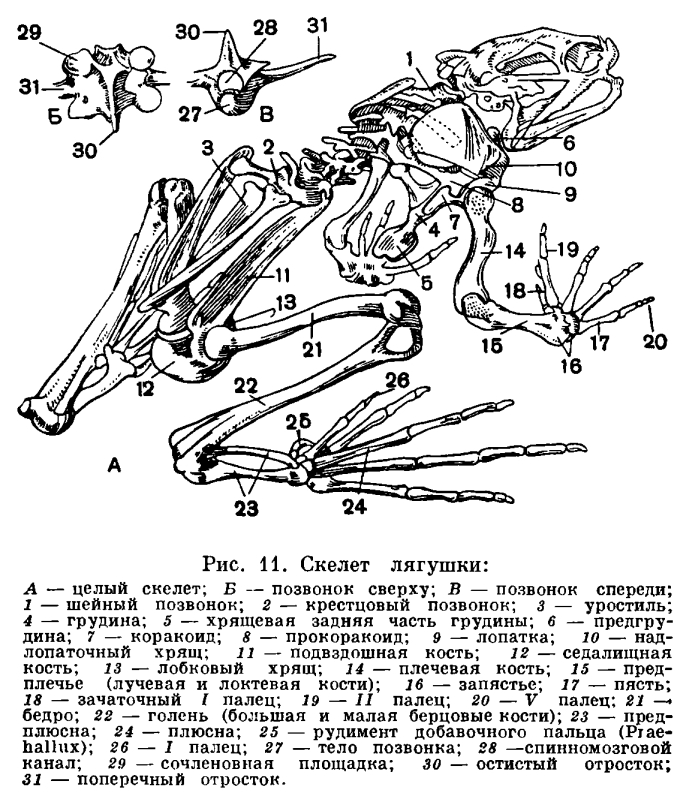 В связи с тем что у земноводных конечности несут значительно большую нагрузку, чем органы передвижения водных позвоночных, у них прогрессивно развиваются пояса конечностей, обеспечивающие им прочную опору, но не прикрепляющиеся, как у рыб, к черепу. Развитие плечевого пояса выражается в том, что увеличена ширина как спинной, так и брюшной его части. Тазовый пояс отличается главным образом сильным развитием спинного отдела, отсутствующего у рыб. Прочность тазового пояса значительно возрастает у земноводных также благодаря тому, что он прикрепляется к крестцовому позвонку, получая, таким образом, опору на осевом скелете.Строение конечностей, а также их поясов у земноводных менее совершенно, чем у других наземных позвоночных. Это выражено в том, что пояса конечностей у них не полностью окостеневают и плечевой пояс не имеет связи с осевым скелетом. Плечо и бедро у земноводных располагаются параллельно поверхности земли под прямым углом к продольной оси тела. В результате движения животного однообразны и передвигается оно медленно и неуклюже. Конечности земноводных еще относительно слабы и приподнимают тело над субстратом недостаточно» высоко. Это в полной мере применимо к современным хвостатым и особенно к древним, вымершим формам.Изменения в строении осевого скелета, а также в строении черепа связаны у земноводных с новым типом дыхания и передвижения, появляющимся у них при выходе на сушу, и с особенностями ориентации в окружающей среде. В дополнение к туловищному и хвостовому отделам позвоночника, свойственным рыбам, у амфибий появился шейный и крестцовый отделы, каждый из которых представлен всего лишь одним позвонком. Появление крестцового позвонка связано с прикреплением таза к осевому скелету, ставшему опорой для задних конечностей. Наличие шейного позвонка, подвижно сочлененного с черепом, обеспечивает поднимание и опускание головы. Подвижность головы у амфибий приобрела особое значение в связи с тем, что в их жизни возросла роль органов чувств, расположенных в голове. Кроме того, приподнимание головы способствует выполнению дыхательных движений, а также открыванию рта.
В связи с тем что у земноводных конечности несут значительно большую нагрузку, чем органы передвижения водных позвоночных, у них прогрессивно развиваются пояса конечностей, обеспечивающие им прочную опору, но не прикрепляющиеся, как у рыб, к черепу. Развитие плечевого пояса выражается в том, что увеличена ширина как спинной, так и брюшной его части. Тазовый пояс отличается главным образом сильным развитием спинного отдела, отсутствующего у рыб. Прочность тазового пояса значительно возрастает у земноводных также благодаря тому, что он прикрепляется к крестцовому позвонку, получая, таким образом, опору на осевом скелете.Строение конечностей, а также их поясов у земноводных менее совершенно, чем у других наземных позвоночных. Это выражено в том, что пояса конечностей у них не полностью окостеневают и плечевой пояс не имеет связи с осевым скелетом. Плечо и бедро у земноводных располагаются параллельно поверхности земли под прямым углом к продольной оси тела. В результате движения животного однообразны и передвигается оно медленно и неуклюже. Конечности земноводных еще относительно слабы и приподнимают тело над субстратом недостаточно» высоко. Это в полной мере применимо к современным хвостатым и особенно к древним, вымершим формам.Изменения в строении осевого скелета, а также в строении черепа связаны у земноводных с новым типом дыхания и передвижения, появляющимся у них при выходе на сушу, и с особенностями ориентации в окружающей среде. В дополнение к туловищному и хвостовому отделам позвоночника, свойственным рыбам, у амфибий появился шейный и крестцовый отделы, каждый из которых представлен всего лишь одним позвонком. Появление крестцового позвонка связано с прикреплением таза к осевому скелету, ставшему опорой для задних конечностей. Наличие шейного позвонка, подвижно сочлененного с черепом, обеспечивает поднимание и опускание головы. Подвижность головы у амфибий приобрела особое значение в связи с тем, что в их жизни возросла роль органов чувств, расположенных в голове. Кроме того, приподнимание головы способствует выполнению дыхательных движений, а также открыванию рта.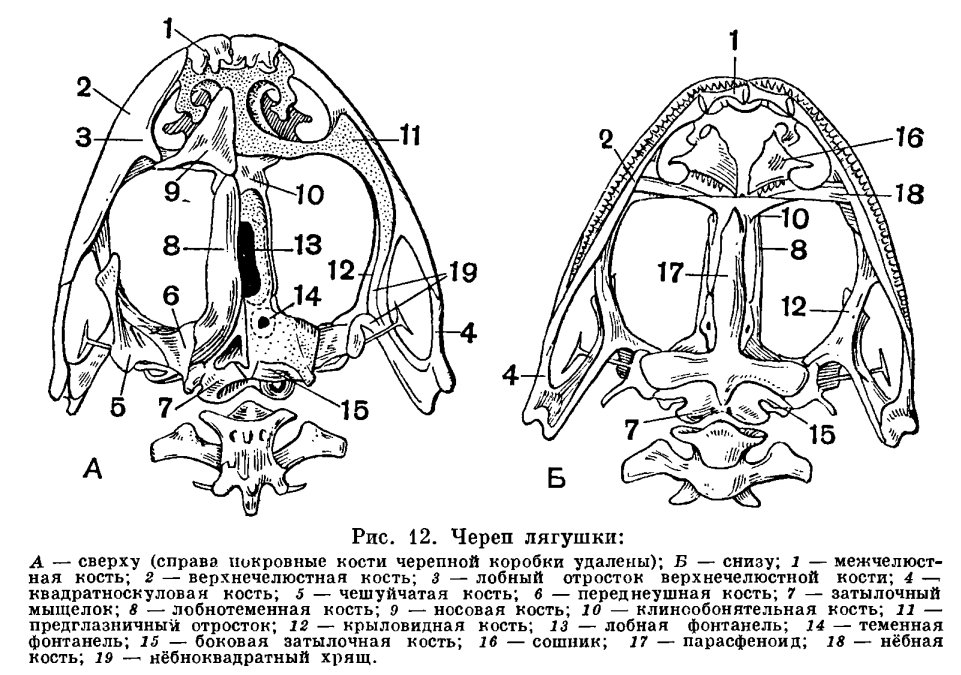 Важнейшие изменения в строении черепа в связи с выходом на сушу произошли в соответствии с изменением в системе органов дыхания. Прежде всего, вместе с исчезновением жабр редуцировались также жаберная крышка и жаберные дужки, которые частично превратились в подъязычный скелет, формирующийся и за счет нижнего элемента подъязычной дуги. Механизм жаберного дыхания у рыб связан не только с движениями жаберной крышки, но также и с движениями челюстной и подъязычной дуг. В связи с этим челюстная дуга должна обладать способностью не только к хватательным движениям — сверху вниз, но и к боковым, дыхательным движениям. Выпадение жаберного дыхания делает боковые движения челюстной дуги ненужными. С другой стороны, механизм легочного дыхания, осуществляемый при помощи опускания и поднимания дна ротовой полости, требует укрепления точек опоры мускулатуры, выполняющей эти движения. Иными словами, возникает необходимость укрепления верхнего отдела челюстной дуги. Это достигается тем, что у земноводных развилась сохраняющаяся у всех наземных позвоночных аутостилия, т. е. нёб-ноквадратный хрящ срастается с осевым черепом и лежащие над ним нёбные и крыловидные кости входят в состав черепной коробки. Аутостилия есть результат изменения механизма дыхания при переходе от дыхания жабрами к дыханию легкими.Действие механизма дыхания у земноводных оказывается тем эффективнее, чем больше расстояние между задними концами ветвей нижней челюсти. Это достигается все более широким раздвиже-нием квадратных костей, к которым при-членяется нижняя челюсть. В конечном итоге весь череп приобрел свойственную земноводным широкую и плоскую форму.В связи с развитием аутостилии и утратой жаберной крышки подъязычно-челюстная (подвесочная) кость у амфибий утрачивает роль челюстного подвеска и роль опоры для жаберной крышки. Она уменьшилась в размерах, вышла из состава челюстного аппарата и превратилась в слуховую косточку — столбик.Таким образом, почти все характерные морфо-физиологические особенности земноводных прямо или косвенно связаны с выходом их на сушу и служат приспособлением к новым условиям обитания.Среди всех систем органов единственное исключение, если не считать почки, составляют органы размножения, которых перестройка почти не коснулась.Формирование яиц у амфибий происходит в парных яичниках, размеры которых, как и у большинства других позвоночных, изменчивы в зависимости от времени года: летом они малы, а к осени и особенно весной велики и переполнены круглыми яйцами темного цвета. Созревшая яйцеклетка, одетая тонкой оболочкой, выпадает из яичника в полость тела. Из полости тела она улавливается воронками яйцеводов. В качестве яйцеводов у земноводных, так же как у большинства рыб, функционируют мюллеровы каналы. Они представляют собой парные, сильно извитые трубки, одним концом впадающие в клоаку, а на другом несущие воронки, открывающиеся в полость тела. Воронки яйцеводов у лягушек прирастают к сердечной сумке, так что при сокращении сердца они попеременно сжимаются и расправляются, захватывая яйца из полости тела. В яйцеводах яйца покрываются оболочками, слизистыми, сильно разбухающими, но не могущими в достаточной мере предохранить яйцо от высыхания. В этом отношении яйца земноводных не отличаются от икринок рыб. Развиваться вне воды, за некоторыми исключениями, они не могут.
Важнейшие изменения в строении черепа в связи с выходом на сушу произошли в соответствии с изменением в системе органов дыхания. Прежде всего, вместе с исчезновением жабр редуцировались также жаберная крышка и жаберные дужки, которые частично превратились в подъязычный скелет, формирующийся и за счет нижнего элемента подъязычной дуги. Механизм жаберного дыхания у рыб связан не только с движениями жаберной крышки, но также и с движениями челюстной и подъязычной дуг. В связи с этим челюстная дуга должна обладать способностью не только к хватательным движениям — сверху вниз, но и к боковым, дыхательным движениям. Выпадение жаберного дыхания делает боковые движения челюстной дуги ненужными. С другой стороны, механизм легочного дыхания, осуществляемый при помощи опускания и поднимания дна ротовой полости, требует укрепления точек опоры мускулатуры, выполняющей эти движения. Иными словами, возникает необходимость укрепления верхнего отдела челюстной дуги. Это достигается тем, что у земноводных развилась сохраняющаяся у всех наземных позвоночных аутостилия, т. е. нёб-ноквадратный хрящ срастается с осевым черепом и лежащие над ним нёбные и крыловидные кости входят в состав черепной коробки. Аутостилия есть результат изменения механизма дыхания при переходе от дыхания жабрами к дыханию легкими.Действие механизма дыхания у земноводных оказывается тем эффективнее, чем больше расстояние между задними концами ветвей нижней челюсти. Это достигается все более широким раздвиже-нием квадратных костей, к которым при-членяется нижняя челюсть. В конечном итоге весь череп приобрел свойственную земноводным широкую и плоскую форму.В связи с развитием аутостилии и утратой жаберной крышки подъязычно-челюстная (подвесочная) кость у амфибий утрачивает роль челюстного подвеска и роль опоры для жаберной крышки. Она уменьшилась в размерах, вышла из состава челюстного аппарата и превратилась в слуховую косточку — столбик.Таким образом, почти все характерные морфо-физиологические особенности земноводных прямо или косвенно связаны с выходом их на сушу и служат приспособлением к новым условиям обитания.Среди всех систем органов единственное исключение, если не считать почки, составляют органы размножения, которых перестройка почти не коснулась.Формирование яиц у амфибий происходит в парных яичниках, размеры которых, как и у большинства других позвоночных, изменчивы в зависимости от времени года: летом они малы, а к осени и особенно весной велики и переполнены круглыми яйцами темного цвета. Созревшая яйцеклетка, одетая тонкой оболочкой, выпадает из яичника в полость тела. Из полости тела она улавливается воронками яйцеводов. В качестве яйцеводов у земноводных, так же как у большинства рыб, функционируют мюллеровы каналы. Они представляют собой парные, сильно извитые трубки, одним концом впадающие в клоаку, а на другом несущие воронки, открывающиеся в полость тела. Воронки яйцеводов у лягушек прирастают к сердечной сумке, так что при сокращении сердца они попеременно сжимаются и расправляются, захватывая яйца из полости тела. В яйцеводах яйца покрываются оболочками, слизистыми, сильно разбухающими, но не могущими в достаточной мере предохранить яйцо от высыхания. В этом отношении яйца земноводных не отличаются от икринок рыб. Развиваться вне воды, за некоторыми исключениями, они не могут.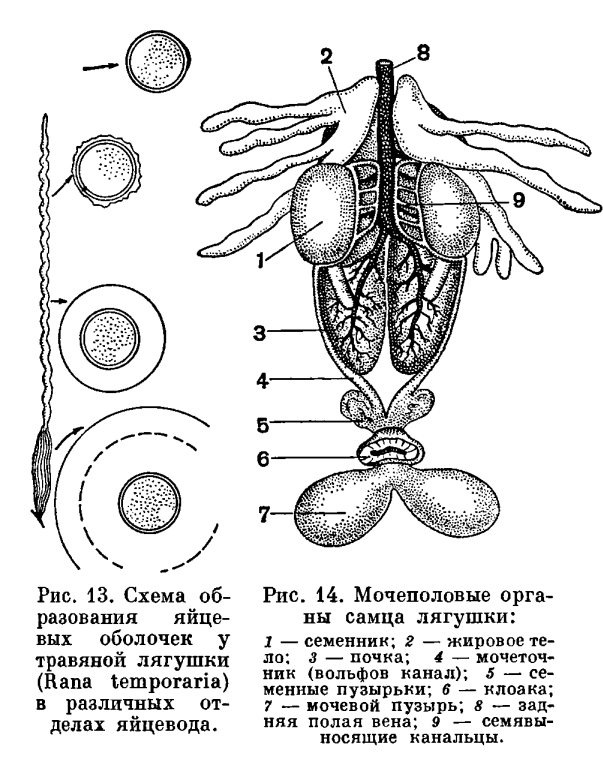 Органы размножения самцов представлены парой семенников. От них отходят многочисленные семявыносящие канальцы, которые, пройдя через почку, впадают в мочеточник.У амфибий, акуловых и двоякодышащих рыб строение органов размножения и выделения, так же как и характер их взаимосвязи, одинаково.Из яиц, обычно развивающихся в воде, у земноводных вылупляются личинки, вся организация которых приспособлена к жизни в воде. Наличие личиночной стадии расширяет возможности питания развивающегося организма, улучшает снабжение его кислородом, создает возможность перемещения в наиболее благоприятные условия существования. Перед появлением на свет головастики бесхвостых амфибий уже имеют одноклеточные железы, выделяющие фермент, который растворяет оболочки. При помощи этого фермента и происходит освобождение животного из яйца.
Органы размножения самцов представлены парой семенников. От них отходят многочисленные семявыносящие канальцы, которые, пройдя через почку, впадают в мочеточник.У амфибий, акуловых и двоякодышащих рыб строение органов размножения и выделения, так же как и характер их взаимосвязи, одинаково.Из яиц, обычно развивающихся в воде, у земноводных вылупляются личинки, вся организация которых приспособлена к жизни в воде. Наличие личиночной стадии расширяет возможности питания развивающегося организма, улучшает снабжение его кислородом, создает возможность перемещения в наиболее благоприятные условия существования. Перед появлением на свет головастики бесхвостых амфибий уже имеют одноклеточные железы, выделяющие фермент, который растворяет оболочки. При помощи этого фермента и происходит освобождение животного из яйца. У только что вылупившегося головастика травяной или остромордой лягушки части тела еще едва обозначены. Голова отделена от туловища легким перехватом, а задний конец вытянут в коротенький вырост — хвост. Хвост окружен нешироким плавником, идущим вдоль спины личинки, весь скелет которой представлен лишь хордой. В это время личинки еще не способны активно передвигаться и большими гроздьями неподвижно висят на пустых оболочках яиц, из которых вышли, удерживаясь особым личиночным органом прилипания — подковообразной присоской. На ранней стадии развития личинка питается остаточным желтком, не использованным в яйце во время развития зародыша. На нижней поверхности головы такой личинки уже ясно обозначено ротовое впячивание, прилегающее к тонкостенной широкой глотке. Однако, в связи с тем что личинка еще активно не питается, сообщения между этими двумя отделами пищеварительной системы нет и ротовое отверстие отсутствует. Широкая глотка переходит в резко суживающийся пищевод, за которым следует короткий, не разделенный на отделы кишечник. Полость пищеварительной трубки заполнена желтком, и только на спинной стороне ее йроходит узкий канал, загибающийся шз, где кишка сообщается с внешц средой через заднепроходное отверстъ . Раннее развитие анального отверстия связано с тем, что в задний отдел кишки открываются протоки выделительных органов, которые развиваются и функционируют у головастика очень рано.Органы выделения — парные округлые тела — заключены в спинном отделе стенки тела, непосредственно сзади околосердечной полости. Почки головастика — наиболее просто устроенные выделительные органы из всех известных у позвоночных животных. В зависимости от своего местоположения они носят название головных почек. Головные почки функционируют только у личинок.Развитие. У только что вылупившегося головастика процесс дыхания по сравнению с таковым у зародыша, находящегося в яйце, значительно интенсивнее, так как незадолго до вылупления у зародышей появляются наружные жабры. Жабры образуются в виде двух пар небольших отростков по бокам головы. К моменту вылупления развивается третья пара наружных жабр. В дальнейшем они разделяются на целый ряд разрастающихся и ветвящихся жаберных лепестков. Наружные жабры, специальные личиночные органы дыхания, представляют собой видоизмененные внутренние жабры, разросшиеся из жаберной, полости наружу. В отличие от внутренних жабр они постоянно омываются свежей водой и могут функционировать на очень ранних стадиях развития личинки, прежде чем появится механизм, нагнетающий воду к внутренним жабрам.Несмотря на то что на описываемой стадии функционируют только наружные жабры, у головастиков уже имеются органы дыхания, приходящие им на смену, —внутренние жабры. Жаберные мешки закладываются у зародыша на очень ранних стадиях развития. Они возникают как вертикальные кармановидные складки глотки, растущие кнаружи. В стенке глотки, лежащей между соседними жаберными мешками, развивается скелет жаберного аппарата — жаберные дужки. У только что вылупившегося головастика есть также и зачатки легких.В связи с тем что на первой стадии развития личинки еще не отыскивают активно пищу и малоподвижны, органы чувств у них развиты слабо. На переднем конце головы есть парные ямки — органы обоняния, недоразвитые глаза и зачаток органа слуха в виде слуховых пузырьков, лежащих под кожей по бокам головы. Важнейшее значение в это время имеют органы боковой линии — небольшие сосочки эпидермиса, которые расположены рядами вдоль всего тела, вокруг глаз и на других частях головы. Они воспринимают колебания воды, исходящие от различных окружающих головастиков предметов.
У только что вылупившегося головастика травяной или остромордой лягушки части тела еще едва обозначены. Голова отделена от туловища легким перехватом, а задний конец вытянут в коротенький вырост — хвост. Хвост окружен нешироким плавником, идущим вдоль спины личинки, весь скелет которой представлен лишь хордой. В это время личинки еще не способны активно передвигаться и большими гроздьями неподвижно висят на пустых оболочках яиц, из которых вышли, удерживаясь особым личиночным органом прилипания — подковообразной присоской. На ранней стадии развития личинка питается остаточным желтком, не использованным в яйце во время развития зародыша. На нижней поверхности головы такой личинки уже ясно обозначено ротовое впячивание, прилегающее к тонкостенной широкой глотке. Однако, в связи с тем что личинка еще активно не питается, сообщения между этими двумя отделами пищеварительной системы нет и ротовое отверстие отсутствует. Широкая глотка переходит в резко суживающийся пищевод, за которым следует короткий, не разделенный на отделы кишечник. Полость пищеварительной трубки заполнена желтком, и только на спинной стороне ее йроходит узкий канал, загибающийся шз, где кишка сообщается с внешц средой через заднепроходное отверстъ . Раннее развитие анального отверстия связано с тем, что в задний отдел кишки открываются протоки выделительных органов, которые развиваются и функционируют у головастика очень рано.Органы выделения — парные округлые тела — заключены в спинном отделе стенки тела, непосредственно сзади околосердечной полости. Почки головастика — наиболее просто устроенные выделительные органы из всех известных у позвоночных животных. В зависимости от своего местоположения они носят название головных почек. Головные почки функционируют только у личинок.Развитие. У только что вылупившегося головастика процесс дыхания по сравнению с таковым у зародыша, находящегося в яйце, значительно интенсивнее, так как незадолго до вылупления у зародышей появляются наружные жабры. Жабры образуются в виде двух пар небольших отростков по бокам головы. К моменту вылупления развивается третья пара наружных жабр. В дальнейшем они разделяются на целый ряд разрастающихся и ветвящихся жаберных лепестков. Наружные жабры, специальные личиночные органы дыхания, представляют собой видоизмененные внутренние жабры, разросшиеся из жаберной, полости наружу. В отличие от внутренних жабр они постоянно омываются свежей водой и могут функционировать на очень ранних стадиях развития личинки, прежде чем появится механизм, нагнетающий воду к внутренним жабрам.Несмотря на то что на описываемой стадии функционируют только наружные жабры, у головастиков уже имеются органы дыхания, приходящие им на смену, —внутренние жабры. Жаберные мешки закладываются у зародыша на очень ранних стадиях развития. Они возникают как вертикальные кармановидные складки глотки, растущие кнаружи. В стенке глотки, лежащей между соседними жаберными мешками, развивается скелет жаберного аппарата — жаберные дужки. У только что вылупившегося головастика есть также и зачатки легких.В связи с тем что на первой стадии развития личинки еще не отыскивают активно пищу и малоподвижны, органы чувств у них развиты слабо. На переднем конце головы есть парные ямки — органы обоняния, недоразвитые глаза и зачаток органа слуха в виде слуховых пузырьков, лежащих под кожей по бокам головы. Важнейшее значение в это время имеют органы боковой линии — небольшие сосочки эпидермиса, которые расположены рядами вдоль всего тела, вокруг глаз и на других частях головы. Они воспринимают колебания воды, исходящие от различных окружающих головастиков предметов.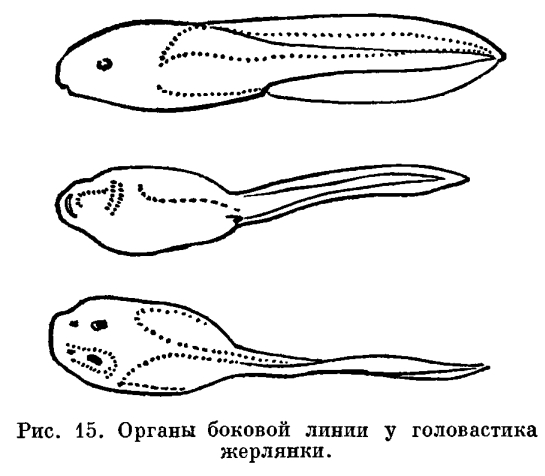 Головной мозг этого крошечного животного, не достигающего 10 мм длины, в основных чертах сформирован. Но от головного мозга взрослых форм он отличается почти полным отсутствием мозжечка и передних полушарий. Вместо полушарий мозга имеются только небольшие парные зачатки их.Крупные изменения в жизни и строении головастика происходят через несколько дней после вылупления, когда он достигает 14 мм длины. В это время желток оказывается полностью израсходованным, у личинок прорывается рот и начинают быстро расти зубы. Вскоре крошечный по сравнению с ртом лягушки рот головастика оказывается окруженным выдающимися вперед бахромчатыми губами, которые образуют небольшой конический хоботок. Верхняя и нижняя губы в углах рта без заметного перерыва соединяются между собой. Нижняя губа длиннее, шире, мягче и подвижнее, чем верхняя. По свободному краю ее идут в несколько рядов небольшие мясистые сосочки, выполняющие , по-видимому, осязательную функцию. Они скопляются преимущественно в углах рта. Перегородкой, отделяющей хоботок от ротовой полости, служит развивающийся к этому времени клюв. Он состоит из двух крепких роговых челюстей. По внутренней поверхности обеих губ образуются поперечные складки, на гребнях которых, так же как и по свободному краю губ, появляются небольшие черные роговые зубчики. Каждый из них — это одна видоизмененная эпителиальная клетка. Зубчик быстро изнашивается, и взамен немедленно возникает точно такой же.Для черепа головастика характерно раннее появление и крупная величина губных хрящей, раннее прикрепление хорошо развитой челюстной дужки к мозговой коробке, массивная подчелюстная дуга. Все эти особенности есть приспособление к укреплению скелета, поддерживающего роговые челюсти. Они связаны с развитием мускулатуры, двигающей челюсти. При метаморфозе они исчезают или существенно видоизменяются.
Головной мозг этого крошечного животного, не достигающего 10 мм длины, в основных чертах сформирован. Но от головного мозга взрослых форм он отличается почти полным отсутствием мозжечка и передних полушарий. Вместо полушарий мозга имеются только небольшие парные зачатки их.Крупные изменения в жизни и строении головастика происходят через несколько дней после вылупления, когда он достигает 14 мм длины. В это время желток оказывается полностью израсходованным, у личинок прорывается рот и начинают быстро расти зубы. Вскоре крошечный по сравнению с ртом лягушки рот головастика оказывается окруженным выдающимися вперед бахромчатыми губами, которые образуют небольшой конический хоботок. Верхняя и нижняя губы в углах рта без заметного перерыва соединяются между собой. Нижняя губа длиннее, шире, мягче и подвижнее, чем верхняя. По свободному краю ее идут в несколько рядов небольшие мясистые сосочки, выполняющие , по-видимому, осязательную функцию. Они скопляются преимущественно в углах рта. Перегородкой, отделяющей хоботок от ротовой полости, служит развивающийся к этому времени клюв. Он состоит из двух крепких роговых челюстей. По внутренней поверхности обеих губ образуются поперечные складки, на гребнях которых, так же как и по свободному краю губ, появляются небольшие черные роговые зубчики. Каждый из них — это одна видоизмененная эпителиальная клетка. Зубчик быстро изнашивается, и взамен немедленно возникает точно такой же.Для черепа головастика характерно раннее появление и крупная величина губных хрящей, раннее прикрепление хорошо развитой челюстной дужки к мозговой коробке, массивная подчелюстная дуга. Все эти особенности есть приспособление к укреплению скелета, поддерживающего роговые челюсти. Они связаны с развитием мускулатуры, двигающей челюсти. При метаморфозе они исчезают или существенно видоизменяются. Ротовой аппарат головастиков приспособлен к соскабливанию пищи с подводных предметов и водорослей. Он свойствен громадному большинству личинок бесхвостых амфибий, которые обычно питаются кормами, изобилующими в водоемах, сравнительно легко заменяя один другим. Часто образуя большие скопления, личинки иначе не могли бы найти для себя достаточного количества пищи. Начиная активно питаться, головастики поедают прежде всего яйцевые оболочки, на которых они висели первые дни жизни.Кишечник головастиков не только относительно, но и абсолютно длиннее, чем у взрослых форм. В связи с питанием резко щелочными кормами у них, как и у карповых рыб, не развивается желудок, в котором протекает кислотная фаза пищеварения, требующая затраты большого количества энергии для нейтрализации щелочной пищи.Личинка, перешедшая к активному способу питания, начинает также и активно двигаться. По форме она уже отличается от только что вылупившейся. Тело ее становится более округлым, хвост удлиняется и расширяется за счет развития вокруг него плавника, превращаясь в сильный орган движения. Однако значительно развившийся хвостовой плавник служит не только для передвижения. В нем появляется мощная сеть капиллярных сосудов, и он становится дополнительным органом дыхания. Этим не ограничив аются изменения, происходящие с органами дыхания на стадии прорыва рта. Наружные жабры к этому времени достигают максимального развития. В то же время вступают в действие постепенно заменяющие их внутренние жабры. Начинается это с того, что жаберные мешки прорываются и образуются узкие вертикальные отверстия, ведущие из глотки наружу, — жаберные щели. Тотчас же после открытия жаберных щелей начинают развиваться внутренние жабры в виде рядов небольших сосочков по краю жаберной щели. Они быстро растут и ветвятся, образуя жаберные пучки.Раньше чем откроется рот, возникают жаберные крышечки в виде складок кожи по бокам головы. Эти складки вскоре соединяются друг с другом на нижней поверхности головы. Вслед за открытием рта складки начинают быстро расти назад, как чехлом прикрывая жаберные щели. Задний край складки сливается со стенкой тела вдоль всей правой стороны, а с левой он остается свободным и вытягивается назад в виде короткой трубочки. Посредством последней заключенная под крышечками полость сообщается с внешней средой.После образования жаберных крышек, примерно на восьмой день, наружные жабры быстро исчезают, причем с левой стороны они сохраняются несколько дольше и нередко в течение некоторого времени высовываются из-под жаберной крышки наружу через имеющееся здесь отверстие.Исчезновение наружных жабр и замена их внутренними связаны, видимо, с переходом к подвижному образу жизни, при котором наружные жабры становятся более уязвимыми. Внутренние жабры, приходящие на смену наружным, имеют большую поверхность и полнее удовлетворяют требования организма в кислороде.Вскоре после прорыва рта заканчивается развитие жаберных сосудов и возникают капилляры во внутренних жабрах.Так заканчивается формирование личиночных органов, обеспечивающих головастику подвижность, максимум кислорода и возможность активного питания.Возрастающая подвижность животного и развитие мускулатуры сопровождаются развитием скелета. Вокруг хорды постепенно образуется позвоночник. Увеличивающаяся активность приводит и к некоторым усложнениям в строении органов чувств. Слуховой пузырек, например, делится на два отдела, и образуются полукружные каналы. Активное питание, движение, усиление окислительных процессов повышают обмен веществ, в связи с чем на данном этапе заканчивается формирование печени и головной почки, а также начинает закладываться выделительный орган, функционирующий во взрослом состоянии. Он возникает позади головной почки.Примерно на 20—26-й день, когда головастики достигают 24—26 мм длины, появляются одновременно передние и задние конечности. Задние конечности хорошо заметны сразу, а передние долгое время скрыты под жаберной крышкой.На стадии появления конечностей у головастика прорываются внутренние ноздри, уже есть гортанная щель, прикрытая надгортанником, гортань и парные тонкостенные сосудистые мешки — легкие, доходящие до заднего конца полости тела. Еще не скоро головастик выйдет на сушу и станет взрослой лягушкой, а система наземных органов дыхания у него уже развита.Важные изменения происходят и в органах кровообращения. В единственном до сих пор предсердии появляется перегородка, делящая его на правую и левую части. Примерно к этому моменту заканчивают развитие и вступают в связь с легкими легочные вены. Формируется малый круг кровообращения.Приблизительно на 38-й день развития в конечностях образуются суставы. К концу второго месяца наряду с жабрами начинают функционировать легкие. Наступает время метаморфоза.Нет сомнения, что головастик с первых дней своего существования находится в состоянии метаморфоза, с каждым днем приобретая все новые и новые особенности, характерные для взрослого животного, ведущего наземный образ жизни. Причем на каждом предыдущем этапе развиваются системы органов, начинающие функционировать на последующем. Однако метаморфозом обычно именуют те изменения, которые происходят в непосредственной связи с переменой среды обитания и приводят к потере личиночных органов.Метаморфические изменения происходят под воздействием гормонов щитовидной железы, которая к этому времени заканчивает свое формирование. Эти изменения касаются прежде всего органов пищеварения. Животное перестает питаться, и у него преобразуется кишечник. Затем на 50-й день освобождаются, прорывая жаберные крышки, передние конечности. Вслед за этим происходит исчезновение жабр, и весь поток венозной крови проходит теперь через легкие. Далее преобразуется ротовой аппарат. Глаза, которые до сих пор были малы и прикрыты тонкой кожей, становятся крупными и выпуклыми. Заканчивается формирование внутреннего и среднего уха, исчезают органы боковой линии. Окончательно развиваются большие полушария мозга.Заканчивается формирование и окостенение скелета, происходит половая диффе-ренцировка, исчезает головная почка, изменяется строение кожи, постепенно рассасывается и исчезает хвост.Большое количество личиночных органов у головастиков бесхвостых земноводных приводит к тому, что в период метаморфоза происходят глубокие и многочисленные, быстро следующие друг за другом изменения строения животного. Такой тип метаморфоза получил название некробиотического.
Ротовой аппарат головастиков приспособлен к соскабливанию пищи с подводных предметов и водорослей. Он свойствен громадному большинству личинок бесхвостых амфибий, которые обычно питаются кормами, изобилующими в водоемах, сравнительно легко заменяя один другим. Часто образуя большие скопления, личинки иначе не могли бы найти для себя достаточного количества пищи. Начиная активно питаться, головастики поедают прежде всего яйцевые оболочки, на которых они висели первые дни жизни.Кишечник головастиков не только относительно, но и абсолютно длиннее, чем у взрослых форм. В связи с питанием резко щелочными кормами у них, как и у карповых рыб, не развивается желудок, в котором протекает кислотная фаза пищеварения, требующая затраты большого количества энергии для нейтрализации щелочной пищи.Личинка, перешедшая к активному способу питания, начинает также и активно двигаться. По форме она уже отличается от только что вылупившейся. Тело ее становится более округлым, хвост удлиняется и расширяется за счет развития вокруг него плавника, превращаясь в сильный орган движения. Однако значительно развившийся хвостовой плавник служит не только для передвижения. В нем появляется мощная сеть капиллярных сосудов, и он становится дополнительным органом дыхания. Этим не ограничив аются изменения, происходящие с органами дыхания на стадии прорыва рта. Наружные жабры к этому времени достигают максимального развития. В то же время вступают в действие постепенно заменяющие их внутренние жабры. Начинается это с того, что жаберные мешки прорываются и образуются узкие вертикальные отверстия, ведущие из глотки наружу, — жаберные щели. Тотчас же после открытия жаберных щелей начинают развиваться внутренние жабры в виде рядов небольших сосочков по краю жаберной щели. Они быстро растут и ветвятся, образуя жаберные пучки.Раньше чем откроется рот, возникают жаберные крышечки в виде складок кожи по бокам головы. Эти складки вскоре соединяются друг с другом на нижней поверхности головы. Вслед за открытием рта складки начинают быстро расти назад, как чехлом прикрывая жаберные щели. Задний край складки сливается со стенкой тела вдоль всей правой стороны, а с левой он остается свободным и вытягивается назад в виде короткой трубочки. Посредством последней заключенная под крышечками полость сообщается с внешней средой.После образования жаберных крышек, примерно на восьмой день, наружные жабры быстро исчезают, причем с левой стороны они сохраняются несколько дольше и нередко в течение некоторого времени высовываются из-под жаберной крышки наружу через имеющееся здесь отверстие.Исчезновение наружных жабр и замена их внутренними связаны, видимо, с переходом к подвижному образу жизни, при котором наружные жабры становятся более уязвимыми. Внутренние жабры, приходящие на смену наружным, имеют большую поверхность и полнее удовлетворяют требования организма в кислороде.Вскоре после прорыва рта заканчивается развитие жаберных сосудов и возникают капилляры во внутренних жабрах.Так заканчивается формирование личиночных органов, обеспечивающих головастику подвижность, максимум кислорода и возможность активного питания.Возрастающая подвижность животного и развитие мускулатуры сопровождаются развитием скелета. Вокруг хорды постепенно образуется позвоночник. Увеличивающаяся активность приводит и к некоторым усложнениям в строении органов чувств. Слуховой пузырек, например, делится на два отдела, и образуются полукружные каналы. Активное питание, движение, усиление окислительных процессов повышают обмен веществ, в связи с чем на данном этапе заканчивается формирование печени и головной почки, а также начинает закладываться выделительный орган, функционирующий во взрослом состоянии. Он возникает позади головной почки.Примерно на 20—26-й день, когда головастики достигают 24—26 мм длины, появляются одновременно передние и задние конечности. Задние конечности хорошо заметны сразу, а передние долгое время скрыты под жаберной крышкой.На стадии появления конечностей у головастика прорываются внутренние ноздри, уже есть гортанная щель, прикрытая надгортанником, гортань и парные тонкостенные сосудистые мешки — легкие, доходящие до заднего конца полости тела. Еще не скоро головастик выйдет на сушу и станет взрослой лягушкой, а система наземных органов дыхания у него уже развита.Важные изменения происходят и в органах кровообращения. В единственном до сих пор предсердии появляется перегородка, делящая его на правую и левую части. Примерно к этому моменту заканчивают развитие и вступают в связь с легкими легочные вены. Формируется малый круг кровообращения.Приблизительно на 38-й день развития в конечностях образуются суставы. К концу второго месяца наряду с жабрами начинают функционировать легкие. Наступает время метаморфоза.Нет сомнения, что головастик с первых дней своего существования находится в состоянии метаморфоза, с каждым днем приобретая все новые и новые особенности, характерные для взрослого животного, ведущего наземный образ жизни. Причем на каждом предыдущем этапе развиваются системы органов, начинающие функционировать на последующем. Однако метаморфозом обычно именуют те изменения, которые происходят в непосредственной связи с переменой среды обитания и приводят к потере личиночных органов.Метаморфические изменения происходят под воздействием гормонов щитовидной железы, которая к этому времени заканчивает свое формирование. Эти изменения касаются прежде всего органов пищеварения. Животное перестает питаться, и у него преобразуется кишечник. Затем на 50-й день освобождаются, прорывая жаберные крышки, передние конечности. Вслед за этим происходит исчезновение жабр, и весь поток венозной крови проходит теперь через легкие. Далее преобразуется ротовой аппарат. Глаза, которые до сих пор были малы и прикрыты тонкой кожей, становятся крупными и выпуклыми. Заканчивается формирование внутреннего и среднего уха, исчезают органы боковой линии. Окончательно развиваются большие полушария мозга.Заканчивается формирование и окостенение скелета, происходит половая диффе-ренцировка, исчезает головная почка, изменяется строение кожи, постепенно рассасывается и исчезает хвост.Большое количество личиночных органов у головастиков бесхвостых земноводных приводит к тому, что в период метаморфоза происходят глубокие и многочисленные, быстро следующие друг за другом изменения строения животного. Такой тип метаморфоза получил название некробиотического. В период метаморфоза происходит резкое падение роста. Только что метаморфизировавшая сеголетка обычно по размерам значительно меньше головастика. Величина сеголеток тем больше, чем больше величина головастиков перед метаморфозом.Метаморфозом не заканчивается развитие организма. Дальнейший рост, окостенение скелета, развитие зубов и половых желез происходит уже после превращения головастика.Рост отличается значительной продолжительностью и не затормаживается после наступления половозрелости.Развитие хвостатых земноводных значительно отличается от развития бесхвостых. Личинка обыкновенного тритона, покидающая яйцо на 14—20-й день после его откладки, находится на более поздней стадии развития, чем личинка лягушки. Это связано с большим запасом желтка в яйцах хвостатых. У личинки тритона отчетливо выражен хвост, окруженный плавником, имеются зачатки передних конечностей и перисто-разветвленные наружные жабры. Она лишена присоски, но по бокам головы у личинки расположены железистые выросты — балансиры, быстро затем исчезающие. Только что вылупившаяся личинка малоподвижна, но вскоре начинает плавать при помощи хвоста и конечностей. Уже в день выклева у нее обозначена ротовая щель, а на второй день прорывается рот, полностью заканчивающий формирование на 11-й день. У двухдневной личинки вместе со ртом открываются и жаберные щели. Однако внутренние жабры, в отличие от таковых у бесхвостых, не развиваются, а наружные функционируют в течение всего личиночного периода жизни.
В период метаморфоза происходит резкое падение роста. Только что метаморфизировавшая сеголетка обычно по размерам значительно меньше головастика. Величина сеголеток тем больше, чем больше величина головастиков перед метаморфозом.Метаморфозом не заканчивается развитие организма. Дальнейший рост, окостенение скелета, развитие зубов и половых желез происходит уже после превращения головастика.Рост отличается значительной продолжительностью и не затормаживается после наступления половозрелости.Развитие хвостатых земноводных значительно отличается от развития бесхвостых. Личинка обыкновенного тритона, покидающая яйцо на 14—20-й день после его откладки, находится на более поздней стадии развития, чем личинка лягушки. Это связано с большим запасом желтка в яйцах хвостатых. У личинки тритона отчетливо выражен хвост, окруженный плавником, имеются зачатки передних конечностей и перисто-разветвленные наружные жабры. Она лишена присоски, но по бокам головы у личинки расположены железистые выросты — балансиры, быстро затем исчезающие. Только что вылупившаяся личинка малоподвижна, но вскоре начинает плавать при помощи хвоста и конечностей. Уже в день выклева у нее обозначена ротовая щель, а на второй день прорывается рот, полностью заканчивающий формирование на 11-й день. У двухдневной личинки вместе со ртом открываются и жаберные щели. Однако внутренние жабры, в отличие от таковых у бесхвостых, не развиваются, а наружные функционируют в течение всего личиночного периода жизни.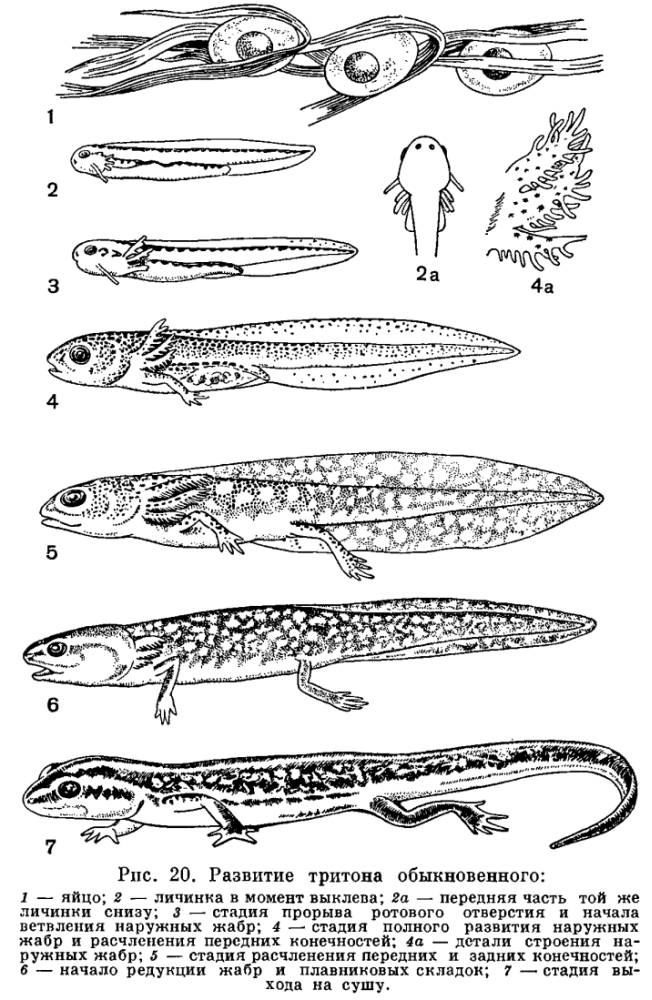 Когда на передних конечностях обозначатся пальцы, начинается развитие задних конечностей; у обыкновенного тритона это происходит примерно на 20-й день личиночной жизни. И на передних, и на задних конечностях пальцы закладываются и развиваются раньше на обращенной внутрь (радиальной) стороне. По-видимому, пальцы радиальной стороны для живущей в воде личинки функционально важнее, чем пальцы внешней (ульналь-ной) стороны. При передвижении личинки по дну водоема конечности вначале касаются дна только радиальной стороной.По характеру питания личинки хвостатых не отличаются от взрослых форм. Они тоже хищники, но нападают на более мелкие организмы. Хищничество среди молоди тритонов возможно потому, что личинки, выходящие из одиночных яиц, отложенных через большие промежутки времени на значительных пространствах, не образуют больших скоплений и могут быть обеспечены кормом. Длина кишечника у личинок хвостатых соответственно равна длине его у взрослых форм.В связи с хищничеством у молоди тритонов, в отличие от бесхвостых, крупные и хорошо развитые глаза рано становятся такими же, как у взрослых. В итоге развивающиеся тритоны имеют небольшое число личиночных органов. В связи с этим метаморфоз у них происходит постепенно, без коренной перестройки организации животного. Такой метаморфоз называют эволютивным. Прежде чем выйти на сушу, личинки переходят к легочному дыханию, теряют наружные жабры, у них зарастают жаберные щели и изменяется строение кожи. Этим и ограничивается их метаморфоз.Некоторым видам хвостатых земноводных присуща задержка в метаморфозе на большее или меньшее время. У других метаморфоз совсем не наступает, но развиваются органы размножения. Это явление получило название неотении, т. е. размножения личинок. Неотения известна у представителей почти всех семейств хвостатых земноводных. У тритонов в некоторые годы бывает много крупных личинок, у которых метаморфоз задержался на длительный период, — это частичная неотения.Наиболее широко известна неотения у американских амбистом, личинки которых называются аксолотлями. Эти личинки способны неопределенно долгое время не метаморфизировать и размножаться. Искусственно, путем воздействия гормона щитовидной железы, можно вызвать превращение аксолотля в амбистому.Неотения представляет большой теоретический интерес, так как указывает на возможность эволюционного развития не от взрослой стадии предков, а от личиночной. Вероятно, постоянножаберные хвостатые амфибии — это не что иное, как неотенические личинки, потерявшие способность к метаморфозу. Они произошли от разных групп хвостатых амфибий. Так, слепой тритон из пещер Техаса — личинка какого-то безлегочного тритона, амфиума — личинка неизвестной саламандры (см. ниже).ОБРАЗ ЖИЗНИОбраз жизни земноводных тесно связан с их строением и физиологией. Несовершенное строение наземных органов дыхания — легких не только накладывает отпечаток на строение ряда других систем органов, но и определяет основные особенности биологии земноводных. Постоянное испарение влаги с поверхности кожи делает земноводных зависимыми от влажности окружающей среды. Не меньшую роль для амфибий, не обладающих теплокровностью, имеет и температура окружающей среды.Однако у разных представителей класса земноводных легкие развиты неодинаково. Так, у лиопельмы капилляры в легких составляют всего лишь 31, 9% от всех капилляров, принимающих участие в газообмене (капилляры легких, кожи и нёба), у желтобрюхой жерлянки — 45, 2%, у обыкновенной чесночницы — 49, 3%. У лягушек легкие развиты лучше, капилляры их составляют у травяной лягушки — 62, 5% , у прудовой — 64, 8%. Особенно велика сеть капилляров в легких у жаб, на долю которых у разных видов приходится от 63, 0 до 77, 8%, а также у обыкновенной квакши (74, 7%) и у гладкой шпорцевой лягушки (65, 9%). Из жаб, обитающих в нашей стране, менее всего развита сеть капилляров в легких у камышовой жабы (63%), но значительно лучше у обыкновенной жабы (71, 5%). У тритонов легочные капилляры составляют всего 24, 5—22, 8%, у мексиканской амбистомы — 48, 9%, у европейской саламандры — 57, 3%, у сирена — 58, 16% и у угревидной амфиумы — 68, 2%. Соответственно меньшему развитию легких у всех исследованных земноводных сильнее развиваются капилляры кожи. Доля капилляров нёба не велика и изменяется у разных видов от 0, 2% до 3%. Большее развитие нёбных капилляров (от 4, 53% до 10, 85%), дополняющих кожное дыхание, наблюдается в семействе безлегочных саламандр. Ротовая полость этих животных усиленно вентилируется. Число колебаний горла достигает 120—180 в минуту.Степень развития той или иной системы органов дыхания меняется с возрастом животного. По мере роста озерных и прудовых лягушек общая длина капилляров кожи у них увеличивается, но количество их на 1 г веса падает, так как лишь небольшая часть этих сосудов формируется вновь. У сеголеток травяных и остромордых лягушек сильно возрастает общая протяженность дыхательных капилляров на 1 г веса.В процессе индивидуального развития у земноводных изменяются и свойства гемоглобина.Разнообразию особенностей строения соответствуют и различия в образе жизни представителей этого класса.Численность и разнообразие земноводных особенно велики в тропиках, отличающихся высокой влажностью и относительно постоянной высокой температурой. По направлению к полюсам число видов земноводных уменьшается. Сухие и особенно сухие и холодные области земного шара бедны амфибиями. Земноводных нет там, где нет хотя бы временных водоемов, так как их яйца, как правило, не могут развиваться вне воды. Благодаря особенностям строения кожи амфибии не могут поддерживать осмотическое давление в гипертонической среде и полностью отсутствуют в соленых водоемах. В соленых водоемах не может идти и развитие яиц этих животных.По характеру местообитания земноводные делятся на две группы: наземных и водных. Первые вне периода размножения далеко уходят от водоемов, вторые всю жизнь проводят в воде или в непосредственной близости от нее. Водные формы господствуют среди хвостатых, к ним относятся и некоторые бесхвостые; в нашей стране, например, озерные и прудовые лягушки, жерлянки. Среди наземных широко представлены древесные обитатели — квакши, веслоногие лягушки, представители семейства короткоголовых и узкоротых. Часть наземных амфибий ведет роющий образ жизни, например почти все безногие и целый ряд бесхвостых.Как правило, наземные обитатели характеризуются лучшим развитием легких и меньшим значением кожи в процессе дыхания. Это делает их менее зависимыми от влажности окружающей среды. Однако из этого правила есть исключения, чаще всего сводящиеся к тому, что у ряда водных форм легкие оказываются так же хорошо развитыми, как и у наземных. Это наблюдается у гладкой шпорцевой лягушки, прудовой лягушки, угревидной амфиумы и сирена. Быть может, эти вторичноводные формы исходно были хорошо приспособлены к наземному способу дыхания, но в дальнейшем были вытеснены в водоемы какими-то иными факторами. Среди наземных обитателей относительно слабо развитыми легкими отличается чесночница. Это легко согласуется с тем, что она большую часть времени проводит зарывшись в землю. Исключительное положение занимает и семейство безлегочных саламандр, половина родов которого — водные животные. Потеря наземных органов дыхания произошла у них, вероятно, в связи с обитанием в горных ручьях. Однако и те виды, которые вторично перешли к жизни во влажных местах на суше, тоже не имеют легких и дышат главным образом через кожу и слизистую ротовой полости.Проникновение земноводных в зоны, неблагоприятные для них, происходит за счет видов, или максимально приспособленных к жизни на суше, или ведущих водный образ жизни. Так, в пределах нашей страны в пустыни проникают жабы и озерные лягушки. Американская жаба-ага — единственный вид среди земноводных, обитающий в предустьевых солоноватых водах, имеет наиболее ороговевшую кожу и наиболее совершенное легкое. В местах с неблагоприятными условиями существования наземные формы могут переходить к водному образу жизни (остромордая лягушка — в засушливых зонах, травяная и остромордая лягушки — в тундре).
Когда на передних конечностях обозначатся пальцы, начинается развитие задних конечностей; у обыкновенного тритона это происходит примерно на 20-й день личиночной жизни. И на передних, и на задних конечностях пальцы закладываются и развиваются раньше на обращенной внутрь (радиальной) стороне. По-видимому, пальцы радиальной стороны для живущей в воде личинки функционально важнее, чем пальцы внешней (ульналь-ной) стороны. При передвижении личинки по дну водоема конечности вначале касаются дна только радиальной стороной.По характеру питания личинки хвостатых не отличаются от взрослых форм. Они тоже хищники, но нападают на более мелкие организмы. Хищничество среди молоди тритонов возможно потому, что личинки, выходящие из одиночных яиц, отложенных через большие промежутки времени на значительных пространствах, не образуют больших скоплений и могут быть обеспечены кормом. Длина кишечника у личинок хвостатых соответственно равна длине его у взрослых форм.В связи с хищничеством у молоди тритонов, в отличие от бесхвостых, крупные и хорошо развитые глаза рано становятся такими же, как у взрослых. В итоге развивающиеся тритоны имеют небольшое число личиночных органов. В связи с этим метаморфоз у них происходит постепенно, без коренной перестройки организации животного. Такой метаморфоз называют эволютивным. Прежде чем выйти на сушу, личинки переходят к легочному дыханию, теряют наружные жабры, у них зарастают жаберные щели и изменяется строение кожи. Этим и ограничивается их метаморфоз.Некоторым видам хвостатых земноводных присуща задержка в метаморфозе на большее или меньшее время. У других метаморфоз совсем не наступает, но развиваются органы размножения. Это явление получило название неотении, т. е. размножения личинок. Неотения известна у представителей почти всех семейств хвостатых земноводных. У тритонов в некоторые годы бывает много крупных личинок, у которых метаморфоз задержался на длительный период, — это частичная неотения.Наиболее широко известна неотения у американских амбистом, личинки которых называются аксолотлями. Эти личинки способны неопределенно долгое время не метаморфизировать и размножаться. Искусственно, путем воздействия гормона щитовидной железы, можно вызвать превращение аксолотля в амбистому.Неотения представляет большой теоретический интерес, так как указывает на возможность эволюционного развития не от взрослой стадии предков, а от личиночной. Вероятно, постоянножаберные хвостатые амфибии — это не что иное, как неотенические личинки, потерявшие способность к метаморфозу. Они произошли от разных групп хвостатых амфибий. Так, слепой тритон из пещер Техаса — личинка какого-то безлегочного тритона, амфиума — личинка неизвестной саламандры (см. ниже).ОБРАЗ ЖИЗНИОбраз жизни земноводных тесно связан с их строением и физиологией. Несовершенное строение наземных органов дыхания — легких не только накладывает отпечаток на строение ряда других систем органов, но и определяет основные особенности биологии земноводных. Постоянное испарение влаги с поверхности кожи делает земноводных зависимыми от влажности окружающей среды. Не меньшую роль для амфибий, не обладающих теплокровностью, имеет и температура окружающей среды.Однако у разных представителей класса земноводных легкие развиты неодинаково. Так, у лиопельмы капилляры в легких составляют всего лишь 31, 9% от всех капилляров, принимающих участие в газообмене (капилляры легких, кожи и нёба), у желтобрюхой жерлянки — 45, 2%, у обыкновенной чесночницы — 49, 3%. У лягушек легкие развиты лучше, капилляры их составляют у травяной лягушки — 62, 5% , у прудовой — 64, 8%. Особенно велика сеть капилляров в легких у жаб, на долю которых у разных видов приходится от 63, 0 до 77, 8%, а также у обыкновенной квакши (74, 7%) и у гладкой шпорцевой лягушки (65, 9%). Из жаб, обитающих в нашей стране, менее всего развита сеть капилляров в легких у камышовой жабы (63%), но значительно лучше у обыкновенной жабы (71, 5%). У тритонов легочные капилляры составляют всего 24, 5—22, 8%, у мексиканской амбистомы — 48, 9%, у европейской саламандры — 57, 3%, у сирена — 58, 16% и у угревидной амфиумы — 68, 2%. Соответственно меньшему развитию легких у всех исследованных земноводных сильнее развиваются капилляры кожи. Доля капилляров нёба не велика и изменяется у разных видов от 0, 2% до 3%. Большее развитие нёбных капилляров (от 4, 53% до 10, 85%), дополняющих кожное дыхание, наблюдается в семействе безлегочных саламандр. Ротовая полость этих животных усиленно вентилируется. Число колебаний горла достигает 120—180 в минуту.Степень развития той или иной системы органов дыхания меняется с возрастом животного. По мере роста озерных и прудовых лягушек общая длина капилляров кожи у них увеличивается, но количество их на 1 г веса падает, так как лишь небольшая часть этих сосудов формируется вновь. У сеголеток травяных и остромордых лягушек сильно возрастает общая протяженность дыхательных капилляров на 1 г веса.В процессе индивидуального развития у земноводных изменяются и свойства гемоглобина.Разнообразию особенностей строения соответствуют и различия в образе жизни представителей этого класса.Численность и разнообразие земноводных особенно велики в тропиках, отличающихся высокой влажностью и относительно постоянной высокой температурой. По направлению к полюсам число видов земноводных уменьшается. Сухие и особенно сухие и холодные области земного шара бедны амфибиями. Земноводных нет там, где нет хотя бы временных водоемов, так как их яйца, как правило, не могут развиваться вне воды. Благодаря особенностям строения кожи амфибии не могут поддерживать осмотическое давление в гипертонической среде и полностью отсутствуют в соленых водоемах. В соленых водоемах не может идти и развитие яиц этих животных.По характеру местообитания земноводные делятся на две группы: наземных и водных. Первые вне периода размножения далеко уходят от водоемов, вторые всю жизнь проводят в воде или в непосредственной близости от нее. Водные формы господствуют среди хвостатых, к ним относятся и некоторые бесхвостые; в нашей стране, например, озерные и прудовые лягушки, жерлянки. Среди наземных широко представлены древесные обитатели — квакши, веслоногие лягушки, представители семейства короткоголовых и узкоротых. Часть наземных амфибий ведет роющий образ жизни, например почти все безногие и целый ряд бесхвостых.Как правило, наземные обитатели характеризуются лучшим развитием легких и меньшим значением кожи в процессе дыхания. Это делает их менее зависимыми от влажности окружающей среды. Однако из этого правила есть исключения, чаще всего сводящиеся к тому, что у ряда водных форм легкие оказываются так же хорошо развитыми, как и у наземных. Это наблюдается у гладкой шпорцевой лягушки, прудовой лягушки, угревидной амфиумы и сирена. Быть может, эти вторичноводные формы исходно были хорошо приспособлены к наземному способу дыхания, но в дальнейшем были вытеснены в водоемы какими-то иными факторами. Среди наземных обитателей относительно слабо развитыми легкими отличается чесночница. Это легко согласуется с тем, что она большую часть времени проводит зарывшись в землю. Исключительное положение занимает и семейство безлегочных саламандр, половина родов которого — водные животные. Потеря наземных органов дыхания произошла у них, вероятно, в связи с обитанием в горных ручьях. Однако и те виды, которые вторично перешли к жизни во влажных местах на суше, тоже не имеют легких и дышат главным образом через кожу и слизистую ротовой полости.Проникновение земноводных в зоны, неблагоприятные для них, происходит за счет видов, или максимально приспособленных к жизни на суше, или ведущих водный образ жизни. Так, в пределах нашей страны в пустыни проникают жабы и озерные лягушки. Американская жаба-ага — единственный вид среди земноводных, обитающий в предустьевых солоноватых водах, имеет наиболее ороговевшую кожу и наиболее совершенное легкое. В местах с неблагоприятными условиями существования наземные формы могут переходить к водному образу жизни (остромордая лягушка — в засушливых зонах, травяная и остромордая лягушки — в тундре).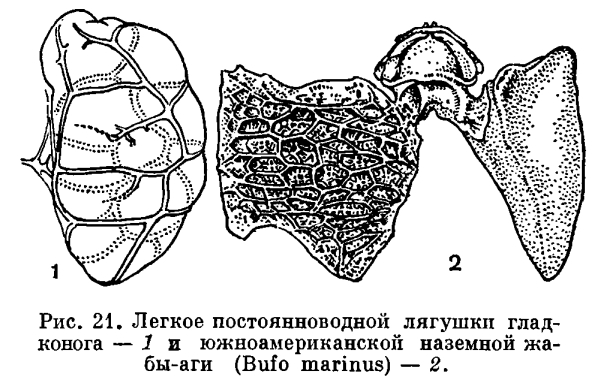 В условиях резко меняющейся температуры и влажности, что свойственно большей части суши, земноводные не в состоянии добывать пищу и вести активный образ жизни в любое время суток на протяжении всего года, Животные избирают для этого периоды с наиболее оптимальными условиями, проводя остальное время в укрытии.Суточная и сезонная цикличность в жизни земноводных — это биологическое приспособление, компенсирующее их морфо-физиологическое несовершенство как наземных обитателей.Характер суточной активности у видов, обитающих в разных биотопах, различен. Наземные виды, в связи с тем что главным ограничивающим фактором для них служит влажность, активны ночью. Следствием этого оказывается относительно короткий активный период и резко выраженный суточный цикл. Напротив, у видов, постоянно связанных с водоемами, для которых влажность перестает играть ограничивающую роль, цикличность выражена слабее. Активность у этих видов может быть либо круглосуточной, либо дневной, и основным ограничивающим фактором оказывается низкая температура. Время активности у них, как правило, значительно длиннее, чем у наземных форм.Характер суточного цикла активности вида может изменяться в зависимости от погоды, времени года и в разных частях ареала животного.Сезонная цикличность во влажных тропических лесах отсутствует. Однако эта зона охватывает лишь незначительную часть суши.В зоне тропических пустынь и саванн высокие температуры господствуют в течение круглого года, но осадки выпадают неравномерно. Кратковременные периоды обильных дождей сменяются продолжительными засушливыми периодами. В этих зонах цикличность выражена резко. Во время засухи амфибии впадают в спячку. В области тропических пустынь, где дожди непериодичны, непериодична и спячка. В саваннах, где дожди идут в определенное время года, постоянно и время спячки.Начиная с субтропических лесов и далее по направлению к полюсам, где колебания температуры по временам года достигают значительной величины, земноводные впадают в зимнюю спячку. Следовательно, здесь главный фактор, определяющий сезонную активность, — температура, а не влажность.Северные и горные по происхождению виды отличаются меньшей чувствительностью к температуре, чем южные и равнинные. Однако спячка здесь длится дольше, чем на юге и на равнинах. Водные формы, как правило, более теплолюбивы и отличаются более продолжительными сроками спячки.Примитивность земноводных как наземных животных, их крайняя зависимость от влажности и температуры, ограничивающая активность, определили и характер питания. Все амфибии питаются беспозвоночными, употребляя в пищу животных самых разнообразных видов. Земноводные схватывают все, что движется, и не берут вполне пригодный в пищу неподвижный корм. Медленное и однообразное передвижение, видимо, определило отсутствие среди земноводных настоящих хищников. Ограниченное время активности полностью исключает для амфибий питание растительными кормами, которые, будучи малопитательными, употребляются животными в больших количествах и добываются в течение большей части суток.Видовые различия в питании земноводных несущественны. Они определяются набором видов беспозвоночных в тех местах обитания, где живут амфибии, способом захватывания пищи и манерой охоты, различием в размерах земноводных и в некоторой мере характером активности охотника и добычи.Только в тропиках, где максимально благоприятны для амфибий условия влажности и температуры и велика насыщенность живыми существами, намечается развитие разных типов питания и появляются специализированные формы. Так, например, мексиканские жабы рода Rhinophrynus, мадагаскарские Rhombophryne, сиамские Glyphoglossus, африканские Nyperolius и Rhynobatrachus, как и ряд других тропических видов, питаются только муравьями и термитами. Американская Rana aesopus, видимо, поедает жаб и лягушек. Rana niedeni кормится крабами.Основные черты биологии размножения земноводных определяются в значительной мере особенностями строения их яиц, в большинстве случаев требующих для развития водной среды. Известное влияние на характер размножения оказывают и влажность, и температура окружающей среды.У громадного большинства земноводных оплодотворение, как и у рыб, наружное. Внутреннее оплодотворение свойственно только безногим и части хвостатых. У бесхвостых оно встречается крайне редко, у 2—3 видов. Однако, в отличие от рыб, даже у тех земноводных, которым свойственно наружное оплодотворение, происходит спаривание. Отсутствует оно лишь у некоторых примитивных хвостатых с наружным оплодотворением. Самцы бесхвостых земноводных крепко обхватывают самку передними лапами. Манера обхватывания отличается у представителей различных систематических групп. Без спаривания процесс икрометания нормально не идет. В искусственных условиях самки, отделенные от самцов, в большинстве случаев погибают от того, что скопившаяся в яйцеводах сильно разбухшая икра разрывает их. В других случаях такие самки по прошествии 1—2 недель откладывают икру, но процесс икрометания теряет свою специфичность. Икра течет из самки, а характерные движения и поза икрометания отсутствуют. Спаривание имеет большое биологическое значение для бесхвостых земноводных. Оно обеспечивает одновременное выделение икры и спермы. Кроме того, благодаря тому что при обхватывании клоаки самца и самки сближены, сперма выделяется прямо на яйца. Это увеличивает процент оплодотворенных яиц. Рефлекс обхватывания у самцов весной выражен очень ярко. Самец держит самку очень крепко, и оторвать его от нее можно лишь с трудом. Возбужденные самцы могут обнимать других самцов, особей другого вида, рыб и даже палки, камни и другие предметы. Однако обычно самец быстро исправляет свою ошибку. Происходит это благодаря существованию различных у разных видов механизмов распознавания пола. У одних опознавательным признаком служит объем обнимаемой особи и ее безмолвие, так как не отложившая икру самка при прикосновении к ней обычно не издает никаких звуков и отличается значительной толщиной. Объятие очень быстро прекращается, если обнимаемое животное издает предупреждающее кваканье. У других распознавание пола происходит при помощи неясного, едва уловимого вибрирующего звука, ощущаемого только в том случае, если держать животное на расстоянии нескольких сантиметров от уха. Этот звук воспринимается самцами, видимо, тактильно при соприкосновении с телом животного, издающего звук. Эта предостерегающая вибрация заставляет самца немедленно прекратить объятия. Только что отложившая икру самка зеленой жабы при попытках самца схватить ее принимает характерную позу, вытягивая вперед согнутые в локтевом суставе передние лапы, плотно прижимая распластанное тело к субстрату и производя отталкивающее движение спиной. Когда самка принимает такую позу, самец быстро оставляет ее в покое.Земноводные с внутренним оплодотворением как правило копулятивных органов не имеют. Только у безногих выворачивающаяся клоака самцов служит копулятивным органом.
В условиях резко меняющейся температуры и влажности, что свойственно большей части суши, земноводные не в состоянии добывать пищу и вести активный образ жизни в любое время суток на протяжении всего года, Животные избирают для этого периоды с наиболее оптимальными условиями, проводя остальное время в укрытии.Суточная и сезонная цикличность в жизни земноводных — это биологическое приспособление, компенсирующее их морфо-физиологическое несовершенство как наземных обитателей.Характер суточной активности у видов, обитающих в разных биотопах, различен. Наземные виды, в связи с тем что главным ограничивающим фактором для них служит влажность, активны ночью. Следствием этого оказывается относительно короткий активный период и резко выраженный суточный цикл. Напротив, у видов, постоянно связанных с водоемами, для которых влажность перестает играть ограничивающую роль, цикличность выражена слабее. Активность у этих видов может быть либо круглосуточной, либо дневной, и основным ограничивающим фактором оказывается низкая температура. Время активности у них, как правило, значительно длиннее, чем у наземных форм.Характер суточного цикла активности вида может изменяться в зависимости от погоды, времени года и в разных частях ареала животного.Сезонная цикличность во влажных тропических лесах отсутствует. Однако эта зона охватывает лишь незначительную часть суши.В зоне тропических пустынь и саванн высокие температуры господствуют в течение круглого года, но осадки выпадают неравномерно. Кратковременные периоды обильных дождей сменяются продолжительными засушливыми периодами. В этих зонах цикличность выражена резко. Во время засухи амфибии впадают в спячку. В области тропических пустынь, где дожди непериодичны, непериодична и спячка. В саваннах, где дожди идут в определенное время года, постоянно и время спячки.Начиная с субтропических лесов и далее по направлению к полюсам, где колебания температуры по временам года достигают значительной величины, земноводные впадают в зимнюю спячку. Следовательно, здесь главный фактор, определяющий сезонную активность, — температура, а не влажность.Северные и горные по происхождению виды отличаются меньшей чувствительностью к температуре, чем южные и равнинные. Однако спячка здесь длится дольше, чем на юге и на равнинах. Водные формы, как правило, более теплолюбивы и отличаются более продолжительными сроками спячки.Примитивность земноводных как наземных животных, их крайняя зависимость от влажности и температуры, ограничивающая активность, определили и характер питания. Все амфибии питаются беспозвоночными, употребляя в пищу животных самых разнообразных видов. Земноводные схватывают все, что движется, и не берут вполне пригодный в пищу неподвижный корм. Медленное и однообразное передвижение, видимо, определило отсутствие среди земноводных настоящих хищников. Ограниченное время активности полностью исключает для амфибий питание растительными кормами, которые, будучи малопитательными, употребляются животными в больших количествах и добываются в течение большей части суток.Видовые различия в питании земноводных несущественны. Они определяются набором видов беспозвоночных в тех местах обитания, где живут амфибии, способом захватывания пищи и манерой охоты, различием в размерах земноводных и в некоторой мере характером активности охотника и добычи.Только в тропиках, где максимально благоприятны для амфибий условия влажности и температуры и велика насыщенность живыми существами, намечается развитие разных типов питания и появляются специализированные формы. Так, например, мексиканские жабы рода Rhinophrynus, мадагаскарские Rhombophryne, сиамские Glyphoglossus, африканские Nyperolius и Rhynobatrachus, как и ряд других тропических видов, питаются только муравьями и термитами. Американская Rana aesopus, видимо, поедает жаб и лягушек. Rana niedeni кормится крабами.Основные черты биологии размножения земноводных определяются в значительной мере особенностями строения их яиц, в большинстве случаев требующих для развития водной среды. Известное влияние на характер размножения оказывают и влажность, и температура окружающей среды.У громадного большинства земноводных оплодотворение, как и у рыб, наружное. Внутреннее оплодотворение свойственно только безногим и части хвостатых. У бесхвостых оно встречается крайне редко, у 2—3 видов. Однако, в отличие от рыб, даже у тех земноводных, которым свойственно наружное оплодотворение, происходит спаривание. Отсутствует оно лишь у некоторых примитивных хвостатых с наружным оплодотворением. Самцы бесхвостых земноводных крепко обхватывают самку передними лапами. Манера обхватывания отличается у представителей различных систематических групп. Без спаривания процесс икрометания нормально не идет. В искусственных условиях самки, отделенные от самцов, в большинстве случаев погибают от того, что скопившаяся в яйцеводах сильно разбухшая икра разрывает их. В других случаях такие самки по прошествии 1—2 недель откладывают икру, но процесс икрометания теряет свою специфичность. Икра течет из самки, а характерные движения и поза икрометания отсутствуют. Спаривание имеет большое биологическое значение для бесхвостых земноводных. Оно обеспечивает одновременное выделение икры и спермы. Кроме того, благодаря тому что при обхватывании клоаки самца и самки сближены, сперма выделяется прямо на яйца. Это увеличивает процент оплодотворенных яиц. Рефлекс обхватывания у самцов весной выражен очень ярко. Самец держит самку очень крепко, и оторвать его от нее можно лишь с трудом. Возбужденные самцы могут обнимать других самцов, особей другого вида, рыб и даже палки, камни и другие предметы. Однако обычно самец быстро исправляет свою ошибку. Происходит это благодаря существованию различных у разных видов механизмов распознавания пола. У одних опознавательным признаком служит объем обнимаемой особи и ее безмолвие, так как не отложившая икру самка при прикосновении к ней обычно не издает никаких звуков и отличается значительной толщиной. Объятие очень быстро прекращается, если обнимаемое животное издает предупреждающее кваканье. У других распознавание пола происходит при помощи неясного, едва уловимого вибрирующего звука, ощущаемого только в том случае, если держать животное на расстоянии нескольких сантиметров от уха. Этот звук воспринимается самцами, видимо, тактильно при соприкосновении с телом животного, издающего звук. Эта предостерегающая вибрация заставляет самца немедленно прекратить объятия. Только что отложившая икру самка зеленой жабы при попытках самца схватить ее принимает характерную позу, вытягивая вперед согнутые в локтевом суставе передние лапы, плотно прижимая распластанное тело к субстрату и производя отталкивающее движение спиной. Когда самка принимает такую позу, самец быстро оставляет ее в покое.Земноводные с внутренним оплодотворением как правило копулятивных органов не имеют. Только у безногих выворачивающаяся клоака самцов служит копулятивным органом. У тритонов, как и у большинства других хвостатых, самцы откладывают студенистые комки — сперматофоры, содержащие семя. Они прикрепляются в воде к окружающим предметам или к дну, а самка схватывает их краями клоаки. Из клоаки сперматофор поступает в карманообразное углубление — сперматеку, и освобождающиеся из нее сперматофоры по мере надобности оплодотворяют постепенно проходящие мимо яйца. У наземных хвостатых амфибий самец обхватывает самку и скручивается вокруг нее так, что их клоаки соприкасаются и сперматофор переходит в клоаку самки.
У тритонов, как и у большинства других хвостатых, самцы откладывают студенистые комки — сперматофоры, содержащие семя. Они прикрепляются в воде к окружающим предметам или к дну, а самка схватывает их краями клоаки. Из клоаки сперматофор поступает в карманообразное углубление — сперматеку, и освобождающиеся из нее сперматофоры по мере надобности оплодотворяют постепенно проходящие мимо яйца. У наземных хвостатых амфибий самец обхватывает самку и скручивается вокруг нее так, что их клоаки соприкасаются и сперматофор переходит в клоаку самки. Среди земноводных встречаются формы, развитие которых связано не в полной мере или совсем не связано с водоемом. Эта особенность возникла независимо (конвергентно) во всех трех отрядах (в 14 семействах из 19) и сопровождается «заботой о потомстве». Наиболее часто наземное развитие встречается в тропиках, и причина его в особенностях кислородного режима водоемов. Мелкие пресноводные тропические водоемы крайне бедны кислородом из-за высоких температур и огромного количества гниющих органических остатков. Кроме того, они чрезмерно насыщены живыми существами. Причиной перехода к наземному развитию может служить также низкая температура воды, сильное течение и, наконец, отсутствие водоемов.Развитие может происходить вне воды полностью или только до стадии личинки. В тропиках для этого используются временные скопления дождевой воды, например в междоузлиях растений или в гнилых пнях и дуплах. Земноводные откладывают яйца и во влажных местах — на берегу водоема, на растениях у воды, в трещинах скал и других подобных местах. В ряде случаев родители сами роют норы или устраивают из пенистой массы гнезда, которые помещают как в воде, так и в почве и на растениях. Так, гнезда над водой устраивают многие квакши и филломедузы. Например, самки филломедуз устраивают гнезда, придерживая задними лапками сближенные края листьев, откладывают в образовавшуюся трубочку яйца, обо лочки которых склеивают края листьев
Среди земноводных встречаются формы, развитие которых связано не в полной мере или совсем не связано с водоемом. Эта особенность возникла независимо (конвергентно) во всех трех отрядах (в 14 семействах из 19) и сопровождается «заботой о потомстве». Наиболее часто наземное развитие встречается в тропиках, и причина его в особенностях кислородного режима водоемов. Мелкие пресноводные тропические водоемы крайне бедны кислородом из-за высоких температур и огромного количества гниющих органических остатков. Кроме того, они чрезмерно насыщены живыми существами. Причиной перехода к наземному развитию может служить также низкая температура воды, сильное течение и, наконец, отсутствие водоемов.Развитие может происходить вне воды полностью или только до стадии личинки. В тропиках для этого используются временные скопления дождевой воды, например в междоузлиях растений или в гнилых пнях и дуплах. Земноводные откладывают яйца и во влажных местах — на берегу водоема, на растениях у воды, в трещинах скал и других подобных местах. В ряде случаев родители сами роют норы или устраивают из пенистой массы гнезда, которые помещают как в воде, так и в почве и на растениях. Так, гнезда над водой устраивают многие квакши и филломедузы. Например, самки филломедуз устраивают гнезда, придерживая задними лапками сближенные края листьев, откладывают в образовавшуюся трубочку яйца, обо лочки которых склеивают края листьев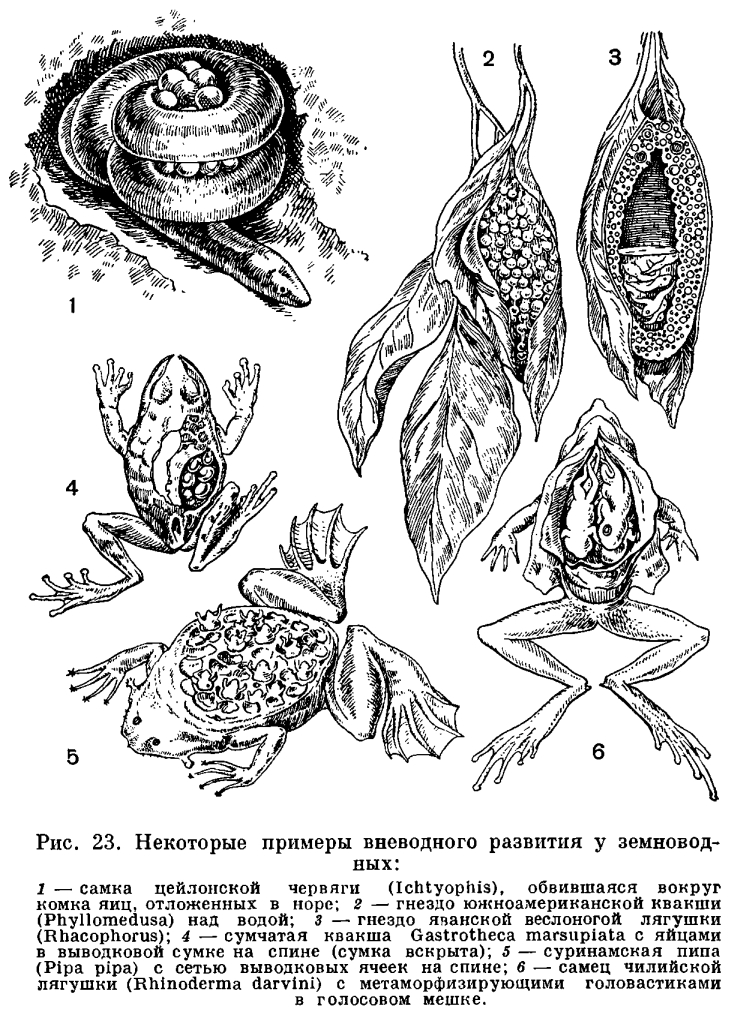
.Необходимая для развития зародыша влажность может поддерживаться и благодаря постоянному контакту кладки с телом одного из родителей. Например, цейлонская червяга обвивает кладку яиц в норе. Развивающиеся яйца могут вынашиваться в специальных образованиях на теле родителей, как у сумчатых квакш и суринамской пипы. Некоторым земноводным свойственно живорождение. Оно встречается у наиболее наземных форм — саламандр и пустынных жаб, обитающих или в засушливых областях, или в холодных высокогорьях.Земноводные, как правило, откладывают большое количество яиц, исчисляемое тысячами. Те из них, которым свойственны внутреннее оплодотворение или забота о потомстве, кладут значительно меньше яиц. Яйца их отличаются большими размерами за счет увеличения запаса питательных веществ в желтке.В период размножения у целого ряда видов амфибий меняется окраска и появляется яркий брачный наряд. Чаще всего обладателями его оказываются самцы, но у некоторых видов это свойственно самкам. У одних в период размножения происходят брачные игры, самцы других устраивают «свадебные концерты». Все это имеет значение для распознавания партнеров своего вида, привлечения и возбуждения самок.
,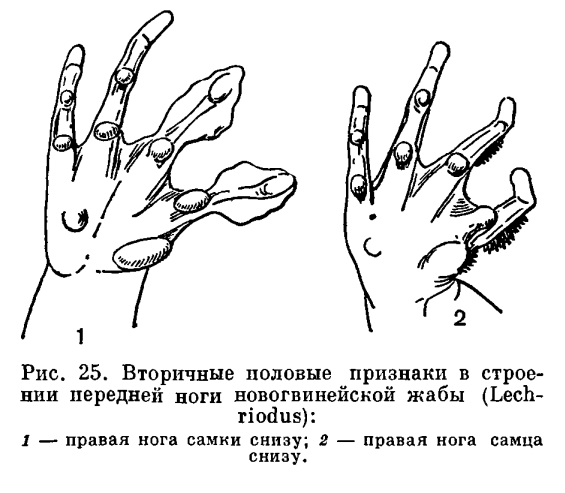 Различия между полами у земноводных не ограничиваются окраской. Как приспособление к более крепкому обхватыванию у самцов лягушек на передних конечностях развиваются шероховатые образования — брачные мозоли, которые у разных видов имеют разную форму, расположение и цвет. Передние лапы самцов по сравнению с таковыми у самок обычно отличаются более развитой мускулатурой и тяжелым скелетом. У самцов, разыскивающих самок, крупнее глаза, лучше развиты все органы чувств и соответственно больше мозг. Большинству поющих самцов свойственны особые голосовые мешки — резонаторы, усиливающие издаваемые самцом звуки. Обладающие более мощным голосом, самцы отличаются от самок и более развитыми легкими. Быть может, это связано с большей активностью и усиленным обменом веществ. Увеличение легких ведет к развитию более мощной брюшной мускулатуры.
Различия между полами у земноводных не ограничиваются окраской. Как приспособление к более крепкому обхватыванию у самцов лягушек на передних конечностях развиваются шероховатые образования — брачные мозоли, которые у разных видов имеют разную форму, расположение и цвет. Передние лапы самцов по сравнению с таковыми у самок обычно отличаются более развитой мускулатурой и тяжелым скелетом. У самцов, разыскивающих самок, крупнее глаза, лучше развиты все органы чувств и соответственно больше мозг. Большинству поющих самцов свойственны особые голосовые мешки — резонаторы, усиливающие издаваемые самцом звуки. Обладающие более мощным голосом, самцы отличаются от самок и более развитыми легкими. Быть может, это связано с большей активностью и усиленным обменом веществ. Увеличение легких ведет к развитию более мощной брюшной мускулатуры.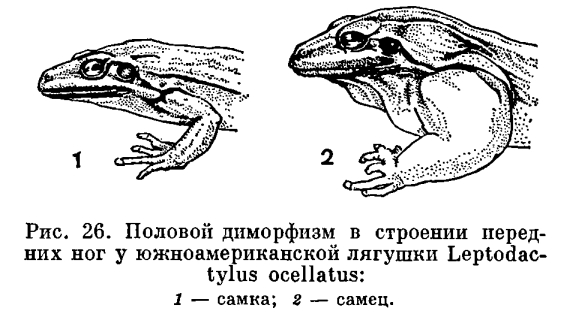 Там, где погода заметно не меняется по сезонам, размножение у земноводных продолжается круглый год. В других областях оно приурочено к наиболее благоприятному времени года, когда оптимальны условия температуры и личинки могут быть обеспечены кормом. В умеренной зоне наземные виды размножаются раньше водных, так как отличаются большей устойчивостью к низким температурам. Способность сохранять активность при относительно низких температурах развивается у наземных видов именно потому, что они вынуждены кормиться в наиболее влажное время суток, которое обычно и наиболее холодное.Помимо раннего размножения, для наземных видов характерно сокращение сроков икрометания, развития яиц и личинок. Это приводит к тому, что они ни во взрослом состоянии, ни в личиночном не сталкиваются или почти не сталкиваются в водоемах с водными видами, избегая тем самым конкуренции. Сокращение сроков развития и размножения позволяет сеголеткам расселиться по суше на большие расстояния и использовать для развития быстро пересыхающие временные водоемы. Последнее позволяет некоторым из них проникать в такие засушливые области, где не могут обитать другие виды.Продолжительность жизни земноводных невелика. Однако известны случаи, когда некоторые виды в неволе жили долго: обыкновенная жаба — 36 лет, гребенчатый тритон — 28, жерлянка — 20—29, квакша — 22, травяная лягушка — 18, камышовая жаба — 16, лягушка-бык — 16, шпорцевая лягушка — 15, чесночница — 11, пипа — 8, прудовая лягушка — 6—10 и т. д.В природе продолжительность жизни амфибий значительно меньшая. Жерлянки, по всей вероятности, в массе не живут больше 2, 5 лет, травяные лягушки — 4—6, озерные — 6—8, так же как серые жабы и многие другие виды. У обыкновенного тритона только единицы доживают до 3-летнего возраста.Гибель земноводных в умеренной зоне происходит главным образом при зимовке на суше в морозные, малоснежные зимы, при заморах, промерзании водоемов или, напротив, во время пересыхания их, когда погибают икра и личинки. Действие этих факторов определяет и колебания численности амфибий по годам, которые могут быть весьма значительными.Сравнительно немного земноводных истребляется другими животными. Чаще они становятся добычей пресмыкающихся и птиц. Так, в Волжско-Камском крае земноводные отмечены в питании 85 видов птиц, у 27 видов которых они составляют от 5 до 42% всей добычи. Обычно доля амфибий в питании животных невелика. Наиболее часто они поедаются аистами, цаплями, хищными птицами, совами, чайками и врановыми. Бесхвостые амфибии, особенно лягушки, становятся добычей птиц во много раз чаще, чем хвостатые. Птицы уничтожают не только взрослых, но и головастиков и икру лягушек. Из млекопитающих поедают земноводных енотовидная собака, норка, выдра, барсук, черный хорь. У черного хоря они составляют около 12% пищи, у норки — 20—22%, у енотовидной собаки — 18—60%, у барсука — 21, 3%. В небольших количествах земноводные входят в пищевой рацион и многих других хищников. В годы, когда основные корма этих животных, например грызуны, малочисленны, роль земноводных как объектов питания значительно повышается. Увеличивается их значение и в бедные кормами сезоны года. На зимовках взрослые и особенно сеголетки становятся жертвой рыб, особенно те виды, которые, как травяная лягушка, не закапываются в ил.Защитные приспособления земноводных от хищников в большинстве случаев имеют пассивный характер. Значительную роль играют кожные ядовитые железы. Выделения их наиболее ядовиты у сухопутных видов: жаб, квакш и наземных саламандр, а из водных — у жерлянок. Напротив, почти совершенно не ядовит секрет желез наших зеленых лягушек и в малой степени ядовит у бурых.Защитное значение имеет также и приспособительная окраска. Подавляющее большинство земноводных окрашено в тон того субстрата, на котором они постоянно держатся. У них весьма часто встречается расчленяющая окраска. Примером ее может служить пигментация задних лап травяной лягушки. Темные полосы на ней создают кажущуюся разорванность сплошной поверхности, а рисунок, составляющийся у сидящей лягушки из полос на разных участках конечности, создает впечатление непрерывности отдельных частей тела. Характер рисунка и окраска у некоторых земноводных в сочетании с поведением может достигать удивительных результатов природного камуфляжа. Так, у обитающей в Южной Африке жабы (Bufo superciliaris) верхняя поверхность окрашена в бурый цвет с темным рисунком. На боках окраска резко меняется на каштаново-коричне-вый цвет, что создает оптический эффект очень темной и резко очерченной тени. Контуры тела крупной жабы оказываются, таким образом, разорванными. Верхняя часть его выглядит как тонкий долго лежавший лист, а нижняя подобна глубокой тени, отбрасываемой этим листом. Иллюзия оказывается полной, когда жаба неподвижно сидит на земле, покрытой настоящими листьями, среди которых она прячется. Очень часто расчленяющий рисунок камуфлирует глаз — наиболее трудно маскируемый орган. Темное височное пятно, присущее всем бурым лягушкам, представляет собой пример такой маскировки.Некоторые земноводные способны в соответствии с окружающей обстановкой быстро менять свою окраску. Приспособительное значение имеет и яркая, контрастирующая, «предупредительная» окраска ядовитых видов, например ярко-оранжевое с черными пятнами брюхо жерлянок или огненной саламандры.Наружных паразитов у земноводных почти нет. Однако у зеленой жабы находили клещей, обитателей птичьих гнезд, а на некоторых южных видах паразитируют пиявки. Зараженность земноводных внутренними паразитами изучена мало. Более или менее известен видовой состав паразитических червей у отдельных видов. Например, у озерной лягушки их обнаружено около 70 видов, у прудовой — более 60, у травяной — около 50. Всего у наших амфибий найдено около 130 видов паразитирующих червей.Слабо изучены болезни земноводных.ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕПитаясь почти исключительно животной пищей, земноводные истребляют насекомых, вредящих садам, огородам, полям, лесам и лугам. Поедают они и различных переносчиков заболеваний, и промежуточных хозяев паразитических червей: комаров, москитов, оводов, слепней и их личинок, а также моллюсков и червей. В уничтожении личинок комаров особая роль в некоторых местах принадлежит жерлянкам, которые оказывают на них большее воздействие, чем гамбузия, умбра и гольян. В некоторых местностях жерлянками уничтожается до 50% личинок комаров в течение одного месяца. Уничтожают личинок и обыкновенные тритоны. Взрослых комаров интенсивно уничтожают сеголетки, особенно тех видов, которые держатся около воды. Процент вредных животных особенно велик в пище обыкновенных тритонов, квакш, жаб, менее других оказываются полезными малоазиатская и прудовая лягушки и гребенчатый тритон.Полезная деятельность земноводных колеблется в разные годы в зависимости от их численности. Неодинакова она в разных стадиях, в разное время года и при разных условиях погоды. Амфибии в больших количествах, чем птицы, поедают насекомых с неприятным запахом и вкусом, а также насекомых, обладающих покровительственной окраской. Кроме того, они охотятся ночью, когда подавляющее большинство насекомоядных птиц спит.На большую пользу земноводных как истребителей вредных насекомых в садах и огородах давно обратили внимание. Садовники Англии, Голландии, Венгрии специально завозили жаб из других стран и выпускали их в сады и оранжереи. В середине 30-х годов с Антильских островов на Гавайские было вывезено около 150 экземпляров жабы-аги. Их размножили, и более миллиона жаб было выпущено на плантации сахарного тростника. Результаты оказались очень хорошими. В этих целях использовали амфибий и у нас. Например, хорошие результаты были получены юннатами, которые выпускали жаб и лягушек на пришкольные участки.Земноводными питаются некоторые промысловые животные (см. выше). Лягушки, кормясь летом наземными беспозвоночными и собираясь на зимовку в водоем, оказываются тем промежуточным звеном, которое расширяет кормность водоемов для рыб за счет наземных беспозвоночных.С точки зрения оценки значения земноводных в производственной деятельности человека привлекают внимание озерные лягушки. Это объясняется тем, что они поедают мальков рыб. Однако выяснилось, что озерные лягушки в естественных условиях уничтожают очень незначительное количество рыбы. Их склонность к этому корму значительно возрастает там, где плотность населения мальков оказывается повышенной, например в водоемах искусственного рыборазведения и на рисовых полях, где выращивают молодь рыбы, причем и здесь сколько-нибудь значительное количество мальков поедается только в определенных местах их концентрации, например у шлюзов.В рыбхозах дельты Волги рыбоядные птицы, ужи и озерная лягушка вместе уничтожают 0, 1—1, 2% поголовья молоди. Следовательно, влияние озерных лягушек на продуктивность водоемов очень незначительно.Головастики тоже не конкурируют из-за пищи с молодью рыб. Они живут за счет диатомовых и зеленых водорослей, малодоступных другим позвоночным животным. Головастиками, в свою очередь, питаются некоторые хищные рыбы, ужи и разнообразные птицы. Так, энергия, запасенная в мельчайших обитателях водоемов — диатомовых и зеленых водорослях, через посредство головастиков используется высшими позвоночными животными.Головастики могут играть существенную роль в выкармливании домашних уток.Простота препарования лягушек, подходящие размеры и живучесть сделали их издавна излюбленными подопытными животными. Огромное количество опытов и наблюдений проводилось и проводится над этими «мучениками науки». Как первые опыты школьника, так и крупнейшие исследования медика и биолога очень часто по сей день связаны с наблюдениями над лягушкой. Большинство приборов экспериментальной медицины и биологии рассчитано на нее. На лягушках постоянно отрабатывается техника физиологического эксперимента. Расход их в лабораториях учебных и научных учреждений достигает десяти тысяч экземпляров в год. Поэтому необходимо принимать меры, чтобы не уничтожить все поголовье. В Англии, ГДР, Польше и ряде других стран лягушки находятся ныне под охраной закона и ловля их запрещена.В ряде стран Европы и Северной Америки лягушек употребляют в пищу. Их не только ловят в природе, но и разводят на специальных фермах. Лягушки являются предметом международной торговли.Земноводные приносят и вред, являясь носителями и переносчиками опасных инфекционных заболеваний, например туляремии. Многие из них — промежуточные хозяева ряда паразитических червей, живущих во взрослом состоянии в организме домашних животных, птиц, пушных зверей.ПРОИСХОЖДЕНИЕ ЗЕМНОВОДНЫХЗемноводные возникли в нижнем или среднем девоне, т. е. не меньше 300 млн. лет назад. Предками их были рыбы, обладающие легкими и такими парными плавниками, из которых могли развиться пятипалые конечности. Таким требованиям удовлетворяют древние ки-степерые рыбы. Они обладают легкими, а в скелете их парных плавников отчетливо обнаруживаются элементы, гомологичные частям скелета наземной пятипалой конечности. На то, что предками земноводных были действительно древние кистеперые рыбы, указывает также поразительное сходство покровных костей их черепа с костями черепа палеозойских земноводных. Так же как у земноводных, у кистеперых встречались как верхние, так и нижние ребра. Напротив, двоякодышащие рыбы, также обладающие легкими, по целому ряду признаков значительно отличаются от земноводных.
Там, где погода заметно не меняется по сезонам, размножение у земноводных продолжается круглый год. В других областях оно приурочено к наиболее благоприятному времени года, когда оптимальны условия температуры и личинки могут быть обеспечены кормом. В умеренной зоне наземные виды размножаются раньше водных, так как отличаются большей устойчивостью к низким температурам. Способность сохранять активность при относительно низких температурах развивается у наземных видов именно потому, что они вынуждены кормиться в наиболее влажное время суток, которое обычно и наиболее холодное.Помимо раннего размножения, для наземных видов характерно сокращение сроков икрометания, развития яиц и личинок. Это приводит к тому, что они ни во взрослом состоянии, ни в личиночном не сталкиваются или почти не сталкиваются в водоемах с водными видами, избегая тем самым конкуренции. Сокращение сроков развития и размножения позволяет сеголеткам расселиться по суше на большие расстояния и использовать для развития быстро пересыхающие временные водоемы. Последнее позволяет некоторым из них проникать в такие засушливые области, где не могут обитать другие виды.Продолжительность жизни земноводных невелика. Однако известны случаи, когда некоторые виды в неволе жили долго: обыкновенная жаба — 36 лет, гребенчатый тритон — 28, жерлянка — 20—29, квакша — 22, травяная лягушка — 18, камышовая жаба — 16, лягушка-бык — 16, шпорцевая лягушка — 15, чесночница — 11, пипа — 8, прудовая лягушка — 6—10 и т. д.В природе продолжительность жизни амфибий значительно меньшая. Жерлянки, по всей вероятности, в массе не живут больше 2, 5 лет, травяные лягушки — 4—6, озерные — 6—8, так же как серые жабы и многие другие виды. У обыкновенного тритона только единицы доживают до 3-летнего возраста.Гибель земноводных в умеренной зоне происходит главным образом при зимовке на суше в морозные, малоснежные зимы, при заморах, промерзании водоемов или, напротив, во время пересыхания их, когда погибают икра и личинки. Действие этих факторов определяет и колебания численности амфибий по годам, которые могут быть весьма значительными.Сравнительно немного земноводных истребляется другими животными. Чаще они становятся добычей пресмыкающихся и птиц. Так, в Волжско-Камском крае земноводные отмечены в питании 85 видов птиц, у 27 видов которых они составляют от 5 до 42% всей добычи. Обычно доля амфибий в питании животных невелика. Наиболее часто они поедаются аистами, цаплями, хищными птицами, совами, чайками и врановыми. Бесхвостые амфибии, особенно лягушки, становятся добычей птиц во много раз чаще, чем хвостатые. Птицы уничтожают не только взрослых, но и головастиков и икру лягушек. Из млекопитающих поедают земноводных енотовидная собака, норка, выдра, барсук, черный хорь. У черного хоря они составляют около 12% пищи, у норки — 20—22%, у енотовидной собаки — 18—60%, у барсука — 21, 3%. В небольших количествах земноводные входят в пищевой рацион и многих других хищников. В годы, когда основные корма этих животных, например грызуны, малочисленны, роль земноводных как объектов питания значительно повышается. Увеличивается их значение и в бедные кормами сезоны года. На зимовках взрослые и особенно сеголетки становятся жертвой рыб, особенно те виды, которые, как травяная лягушка, не закапываются в ил.Защитные приспособления земноводных от хищников в большинстве случаев имеют пассивный характер. Значительную роль играют кожные ядовитые железы. Выделения их наиболее ядовиты у сухопутных видов: жаб, квакш и наземных саламандр, а из водных — у жерлянок. Напротив, почти совершенно не ядовит секрет желез наших зеленых лягушек и в малой степени ядовит у бурых.Защитное значение имеет также и приспособительная окраска. Подавляющее большинство земноводных окрашено в тон того субстрата, на котором они постоянно держатся. У них весьма часто встречается расчленяющая окраска. Примером ее может служить пигментация задних лап травяной лягушки. Темные полосы на ней создают кажущуюся разорванность сплошной поверхности, а рисунок, составляющийся у сидящей лягушки из полос на разных участках конечности, создает впечатление непрерывности отдельных частей тела. Характер рисунка и окраска у некоторых земноводных в сочетании с поведением может достигать удивительных результатов природного камуфляжа. Так, у обитающей в Южной Африке жабы (Bufo superciliaris) верхняя поверхность окрашена в бурый цвет с темным рисунком. На боках окраска резко меняется на каштаново-коричне-вый цвет, что создает оптический эффект очень темной и резко очерченной тени. Контуры тела крупной жабы оказываются, таким образом, разорванными. Верхняя часть его выглядит как тонкий долго лежавший лист, а нижняя подобна глубокой тени, отбрасываемой этим листом. Иллюзия оказывается полной, когда жаба неподвижно сидит на земле, покрытой настоящими листьями, среди которых она прячется. Очень часто расчленяющий рисунок камуфлирует глаз — наиболее трудно маскируемый орган. Темное височное пятно, присущее всем бурым лягушкам, представляет собой пример такой маскировки.Некоторые земноводные способны в соответствии с окружающей обстановкой быстро менять свою окраску. Приспособительное значение имеет и яркая, контрастирующая, «предупредительная» окраска ядовитых видов, например ярко-оранжевое с черными пятнами брюхо жерлянок или огненной саламандры.Наружных паразитов у земноводных почти нет. Однако у зеленой жабы находили клещей, обитателей птичьих гнезд, а на некоторых южных видах паразитируют пиявки. Зараженность земноводных внутренними паразитами изучена мало. Более или менее известен видовой состав паразитических червей у отдельных видов. Например, у озерной лягушки их обнаружено около 70 видов, у прудовой — более 60, у травяной — около 50. Всего у наших амфибий найдено около 130 видов паразитирующих червей.Слабо изучены болезни земноводных.ХОЗЯЙСТВЕННОЕ ЗНАЧЕНИЕПитаясь почти исключительно животной пищей, земноводные истребляют насекомых, вредящих садам, огородам, полям, лесам и лугам. Поедают они и различных переносчиков заболеваний, и промежуточных хозяев паразитических червей: комаров, москитов, оводов, слепней и их личинок, а также моллюсков и червей. В уничтожении личинок комаров особая роль в некоторых местах принадлежит жерлянкам, которые оказывают на них большее воздействие, чем гамбузия, умбра и гольян. В некоторых местностях жерлянками уничтожается до 50% личинок комаров в течение одного месяца. Уничтожают личинок и обыкновенные тритоны. Взрослых комаров интенсивно уничтожают сеголетки, особенно тех видов, которые держатся около воды. Процент вредных животных особенно велик в пище обыкновенных тритонов, квакш, жаб, менее других оказываются полезными малоазиатская и прудовая лягушки и гребенчатый тритон.Полезная деятельность земноводных колеблется в разные годы в зависимости от их численности. Неодинакова она в разных стадиях, в разное время года и при разных условиях погоды. Амфибии в больших количествах, чем птицы, поедают насекомых с неприятным запахом и вкусом, а также насекомых, обладающих покровительственной окраской. Кроме того, они охотятся ночью, когда подавляющее большинство насекомоядных птиц спит.На большую пользу земноводных как истребителей вредных насекомых в садах и огородах давно обратили внимание. Садовники Англии, Голландии, Венгрии специально завозили жаб из других стран и выпускали их в сады и оранжереи. В середине 30-х годов с Антильских островов на Гавайские было вывезено около 150 экземпляров жабы-аги. Их размножили, и более миллиона жаб было выпущено на плантации сахарного тростника. Результаты оказались очень хорошими. В этих целях использовали амфибий и у нас. Например, хорошие результаты были получены юннатами, которые выпускали жаб и лягушек на пришкольные участки.Земноводными питаются некоторые промысловые животные (см. выше). Лягушки, кормясь летом наземными беспозвоночными и собираясь на зимовку в водоем, оказываются тем промежуточным звеном, которое расширяет кормность водоемов для рыб за счет наземных беспозвоночных.С точки зрения оценки значения земноводных в производственной деятельности человека привлекают внимание озерные лягушки. Это объясняется тем, что они поедают мальков рыб. Однако выяснилось, что озерные лягушки в естественных условиях уничтожают очень незначительное количество рыбы. Их склонность к этому корму значительно возрастает там, где плотность населения мальков оказывается повышенной, например в водоемах искусственного рыборазведения и на рисовых полях, где выращивают молодь рыбы, причем и здесь сколько-нибудь значительное количество мальков поедается только в определенных местах их концентрации, например у шлюзов.В рыбхозах дельты Волги рыбоядные птицы, ужи и озерная лягушка вместе уничтожают 0, 1—1, 2% поголовья молоди. Следовательно, влияние озерных лягушек на продуктивность водоемов очень незначительно.Головастики тоже не конкурируют из-за пищи с молодью рыб. Они живут за счет диатомовых и зеленых водорослей, малодоступных другим позвоночным животным. Головастиками, в свою очередь, питаются некоторые хищные рыбы, ужи и разнообразные птицы. Так, энергия, запасенная в мельчайших обитателях водоемов — диатомовых и зеленых водорослях, через посредство головастиков используется высшими позвоночными животными.Головастики могут играть существенную роль в выкармливании домашних уток.Простота препарования лягушек, подходящие размеры и живучесть сделали их издавна излюбленными подопытными животными. Огромное количество опытов и наблюдений проводилось и проводится над этими «мучениками науки». Как первые опыты школьника, так и крупнейшие исследования медика и биолога очень часто по сей день связаны с наблюдениями над лягушкой. Большинство приборов экспериментальной медицины и биологии рассчитано на нее. На лягушках постоянно отрабатывается техника физиологического эксперимента. Расход их в лабораториях учебных и научных учреждений достигает десяти тысяч экземпляров в год. Поэтому необходимо принимать меры, чтобы не уничтожить все поголовье. В Англии, ГДР, Польше и ряде других стран лягушки находятся ныне под охраной закона и ловля их запрещена.В ряде стран Европы и Северной Америки лягушек употребляют в пищу. Их не только ловят в природе, но и разводят на специальных фермах. Лягушки являются предметом международной торговли.Земноводные приносят и вред, являясь носителями и переносчиками опасных инфекционных заболеваний, например туляремии. Многие из них — промежуточные хозяева ряда паразитических червей, живущих во взрослом состоянии в организме домашних животных, птиц, пушных зверей.ПРОИСХОЖДЕНИЕ ЗЕМНОВОДНЫХЗемноводные возникли в нижнем или среднем девоне, т. е. не меньше 300 млн. лет назад. Предками их были рыбы, обладающие легкими и такими парными плавниками, из которых могли развиться пятипалые конечности. Таким требованиям удовлетворяют древние ки-степерые рыбы. Они обладают легкими, а в скелете их парных плавников отчетливо обнаруживаются элементы, гомологичные частям скелета наземной пятипалой конечности. На то, что предками земноводных были действительно древние кистеперые рыбы, указывает также поразительное сходство покровных костей их черепа с костями черепа палеозойских земноводных. Так же как у земноводных, у кистеперых встречались как верхние, так и нижние ребра. Напротив, двоякодышащие рыбы, также обладающие легкими, по целому ряду признаков значительно отличаются от земноводных.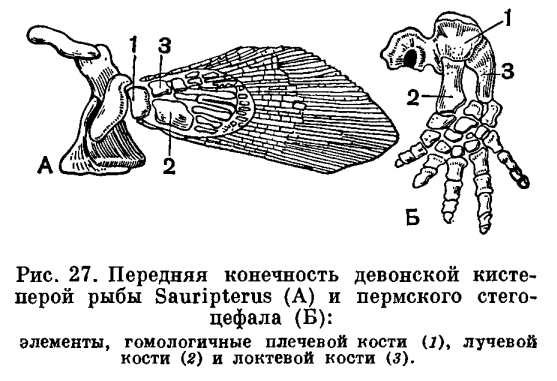 Таким образом, особенности дыхания и передвижения, обеспечивающие возможность выхода на сушу, предки земноводных приобрели, будучи еще настоящими водными позвоночными. Причиной возникновения этих приспособлений, видимо, был особый режим пресных водоемов, в которых жили некоторые кистеперые рыбы, — периодическое пересыхание их или бедность кислородом. Однако ведущим биологическим фактором, определившим разрыв предков земноводных с водоемом, закрепление их на суше, были новые кормовые возможности, найденные ими в новой среде обитания.Наибольшего разнообразия и численности земноводные достигли в каменноугольном и пермском периодах, отличавшихся ровным, влажным и теплым климатом на больших пространствах.Ископаемые палеозойские земноводные относятся к группе стегоцефалов, или панцирноголовых. Их раньше выделяли в особый подкласс стегоцефалов (Stegocephalia) в противоположность современным земноводным, объединяемым в подкласс беспанцирных (Lissamphibia). Однако выяснено, что современные отряды земноводных произошли от различных древних отрядов. Следовательно, такое разделение искусственно.
Таким образом, особенности дыхания и передвижения, обеспечивающие возможность выхода на сушу, предки земноводных приобрели, будучи еще настоящими водными позвоночными. Причиной возникновения этих приспособлений, видимо, был особый режим пресных водоемов, в которых жили некоторые кистеперые рыбы, — периодическое пересыхание их или бедность кислородом. Однако ведущим биологическим фактором, определившим разрыв предков земноводных с водоемом, закрепление их на суше, были новые кормовые возможности, найденные ими в новой среде обитания.Наибольшего разнообразия и численности земноводные достигли в каменноугольном и пермском периодах, отличавшихся ровным, влажным и теплым климатом на больших пространствах.Ископаемые палеозойские земноводные относятся к группе стегоцефалов, или панцирноголовых. Их раньше выделяли в особый подкласс стегоцефалов (Stegocephalia) в противоположность современным земноводным, объединяемым в подкласс беспанцирных (Lissamphibia). Однако выяснено, что современные отряды земноводных произошли от различных древних отрядов. Следовательно, такое разделение искусственно.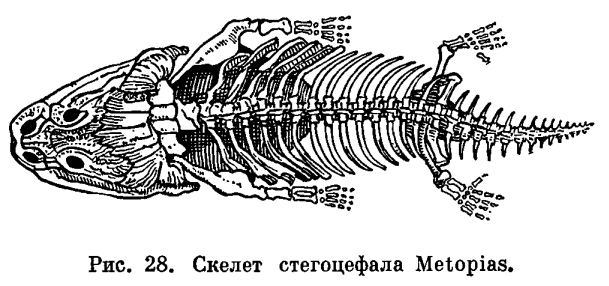 Характернейшей особенностью стегоцефалов был сплошной панцирь из кожных костей, покрывавший черепную коробку сверху и с боков (стегальный череп), так что оставались отверстия только для ноздрей, глаз и теменного органа, который у них был, по-видимому, хорошо развит. Кроме того, у большинства форм был брюшной панцирь, состоявший из налегающих друг на друга костных чешуек и покрывавший брюшную сторону животного. Этот панцирь мог иметь двоякое защитное значение: во время плавания по поверхности водоема (сверху тело не нуждалось в защите, так как тогда сухопутных позвоночных еще не существовало) и при ползании по неровной почве. Стегоцефалы отличались от современных земноводных рядом примитивных признаков (в том числе костным панцирем), унаследованных от кистеперых рыб. Кроме того, у некоторых форм, судя по капролитам (окаменевший кал), имелся спиральный клапан в кишечнике, иногда таз еще не сочленялся с позвоночником, а плечевой пояс сохранял связь с черепом, часто и передние конечности были снабжены пятью пальцами и т. д.В течение каменноугольного и пермского периодов, обозначаемых часто как век земноводных, стегоцефалы достигли большой численности и разнообразия.Учитывая ископаемые остатки стегоцефалов, палеонтологи в настоящее время разделяют класс земноводных на два подкласса: дугопозвонковых (Apsidospondyli) и тонкопозвонковых (Lepospondyli).Первый подкласс делят на два надотряда: надотряд Labyrinthodontia объединяет разнообразных лабиринтодонтов (4—5 отрядов) и надотряд прыгающих (Salientia), включающий всех многообразных современных бесхвостых амфибий (отряд Anura). Ископаемый предок бесхвостых амфибий выделен в отдельный отряд (Proanura).
Характернейшей особенностью стегоцефалов был сплошной панцирь из кожных костей, покрывавший черепную коробку сверху и с боков (стегальный череп), так что оставались отверстия только для ноздрей, глаз и теменного органа, который у них был, по-видимому, хорошо развит. Кроме того, у большинства форм был брюшной панцирь, состоявший из налегающих друг на друга костных чешуек и покрывавший брюшную сторону животного. Этот панцирь мог иметь двоякое защитное значение: во время плавания по поверхности водоема (сверху тело не нуждалось в защите, так как тогда сухопутных позвоночных еще не существовало) и при ползании по неровной почве. Стегоцефалы отличались от современных земноводных рядом примитивных признаков (в том числе костным панцирем), унаследованных от кистеперых рыб. Кроме того, у некоторых форм, судя по капролитам (окаменевший кал), имелся спиральный клапан в кишечнике, иногда таз еще не сочленялся с позвоночником, а плечевой пояс сохранял связь с черепом, часто и передние конечности были снабжены пятью пальцами и т. д.В течение каменноугольного и пермского периодов, обозначаемых часто как век земноводных, стегоцефалы достигли большой численности и разнообразия.Учитывая ископаемые остатки стегоцефалов, палеонтологи в настоящее время разделяют класс земноводных на два подкласса: дугопозвонковых (Apsidospondyli) и тонкопозвонковых (Lepospondyli).Первый подкласс делят на два надотряда: надотряд Labyrinthodontia объединяет разнообразных лабиринтодонтов (4—5 отрядов) и надотряд прыгающих (Salientia), включающий всех многообразных современных бесхвостых амфибий (отряд Anura). Ископаемый предок бесхвостых амфибий выделен в отдельный отряд (Proanura).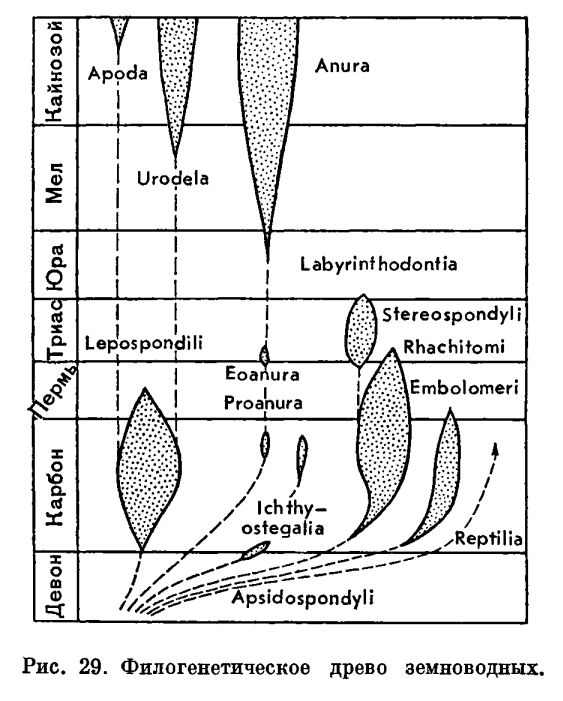 Лабиринтодонты (Labyrinthodontia) были наиболее разнообразны. У них«лаби-ринтодонтный» тип зубов, имевшийся уже у древних кистеперых рыб, достигал наибольшего развития, так что на поперечном разрезе зубов видна необычайно сложная картина разветвляющихся эмалевых петель. К лабиринтодонтам относились почти все крупные земноводные каменноугольного, пермского и триасового периодов. За это время они претерпели большие изменения: ранние формы имели умеренные размеры и рыбообразную форму тела, более поздние достигали очень крупной величины (череп до 1 м и больше), тело их было укорочено и утолщено и заканчивалось коротким толстым хвостом
Лабиринтодонты (Labyrinthodontia) были наиболее разнообразны. У них«лаби-ринтодонтный» тип зубов, имевшийся уже у древних кистеперых рыб, достигал наибольшего развития, так что на поперечном разрезе зубов видна необычайно сложная картина разветвляющихся эмалевых петель. К лабиринтодонтам относились почти все крупные земноводные каменноугольного, пермского и триасового периодов. За это время они претерпели большие изменения: ранние формы имели умеренные размеры и рыбообразную форму тела, более поздние достигали очень крупной величины (череп до 1 м и больше), тело их было укорочено и утолщено и заканчивалось коротким толстым хвостом Второй подкласс — тонкопозвонковых (Lepospondyli) включает три отряда стегоцефалов каменноугольного периода. Это были мелкие, но очень хорошо приспособленные к жизни в воде амфибии, многие из которых вторично утратили конечности. Этих лепоспондил считают исходными формами для современных отрядов хвостатых (Urodela) и безногих (Apoda) земноводных.Почти все стегоцефалы вымерли в пермский период и в течение триасового доживали свой век лишь немногочисленные, крайне специализированные лабиринтодонты.Начиная с верхней юры и нижнего мела появляются вполне типичные бесхвостые и хвостатые земноводные. Амфибии третичного периода уже мало отличаются от ныне живущих.Амфибии, включая ископаемые группы, классифицируются следующим образом.Класс Земноводные (Amphibia)Подкласс Дугопозвонковые (Apsidospondyli)Надотряд Лабиринтодонты (Labyrinthodontia)Отряд Ихтиостегалии (Ichtyostegalia)Отряд Рахитомии (Rhachitomi)Отряд Стереоспондилы (Stereospondyli)Отряд Эмболомери (Embolomeri)Надотряд Прыгающие (Salientia)Отряд Первичнобесхвостые (Proanura)Отряд Бесхвостые (Salientia, или Anura)Подкласс Тонкопозвонковые(Lepospondyli)Отряд Аистопода (Aistopoda)Отряд Нектридиа (Nectridia)Отряд Мнкрозаурии (Microsauria)Отряд Хвостатые (Urodela)Отряд Безногие (Apoda)
Второй подкласс — тонкопозвонковых (Lepospondyli) включает три отряда стегоцефалов каменноугольного периода. Это были мелкие, но очень хорошо приспособленные к жизни в воде амфибии, многие из которых вторично утратили конечности. Этих лепоспондил считают исходными формами для современных отрядов хвостатых (Urodela) и безногих (Apoda) земноводных.Почти все стегоцефалы вымерли в пермский период и в течение триасового доживали свой век лишь немногочисленные, крайне специализированные лабиринтодонты.Начиная с верхней юры и нижнего мела появляются вполне типичные бесхвостые и хвостатые земноводные. Амфибии третичного периода уже мало отличаются от ныне живущих.Амфибии, включая ископаемые группы, классифицируются следующим образом.Класс Земноводные (Amphibia)Подкласс Дугопозвонковые (Apsidospondyli)Надотряд Лабиринтодонты (Labyrinthodontia)Отряд Ихтиостегалии (Ichtyostegalia)Отряд Рахитомии (Rhachitomi)Отряд Стереоспондилы (Stereospondyli)Отряд Эмболомери (Embolomeri)Надотряд Прыгающие (Salientia)Отряд Первичнобесхвостые (Proanura)Отряд Бесхвостые (Salientia, или Anura)Подкласс Тонкопозвонковые(Lepospondyli)Отряд Аистопода (Aistopoda)Отряд Нектридиа (Nectridia)Отряд Мнкрозаурии (Microsauria)Отряд Хвостатые (Urodela)Отряд Безногие (Apoda)
Жизнь животных: в 6-ти томах. — М.: Просвещение. Под редакцией профессоров Н.А.Гладкова, А.В.Михеева. 1970.
.