- Протоплазма или саркода
-
Содержание статьи: Определение и история теории П. — Физические и морфологические свойства П. — Тончайшее строение П. и главнейшие теории. — Химические свойства П. — Физиологические свойства П.: движение, раздражимость, формирующая деятельность, размножение.Определение и история теории П. — П. есть особое органическое вещество, от которого исключительно зависит жизнедеятельность клетки. Так как все растения и животные состоят из одной или чаще — многих клеток, то и все жизненные явления как животных, так и растений должны быть сведены к жизненным проявлениям этого вещества. Обладая, таким образом, всеми свойствами, характеристичными для живых существ, П. в собственном смысле слова может быть названа живой организованной материей и представляет выдающийся интерес в современной биологии, стремящейся объяснить связь и причину всех жизненных явлений. Теория П. находится в тесной связи с клеточной теорией, созданной в тридцатых годах этого столетия Шлейденом для растений и перенесенной Шванном на животное царство. Клеточная теория была возведена в общий принцип и получила название Шванн-Шлейденовской теории, которая со временем претерпела совершенное преобразование и на ее место выступила теория П. Название П. (πρώτος —первое, πλάσμα — организованное вещество) было впервые введено в науку Пуркинье в 1840 г. для обозначения живого образовательного вещества самых молодых животных зародышей, а затем в 1846 году распространено Молем (Hugo von Mohl) на азотистое полужидкое вещество, заключающееся в полости растительных клеток, которое уже было наблюдаемо Шлейденом и названо, в отличие от клеточного сока, растительной слизью. Моль утверждал, что П. совершенно заполняет внутренность молодых растительных клеток, а у более старых принимает в себя жидкость, собирающуюся в так называемых вакуолях; далее, он доказал, что П. обнаруживает чрезвычайно своеобразные явления движения, которые, хотя были открыты еще в конце прошлого столетия Б. Корти на хлорофилловых зернах растений, но истолкованы неверно, как активные движения этих зерен. К этому открытию присоединились еще другие наблюдения. Кон (Cohn) нашел, что у низших водорослей, при размножении, П. отстает от клеточной оболочки, собирается в овальный комочек, который вскоре, при разрыве оболочки, покидает клетку и начинает свободно плавать при помощи появившихся ресничек. Так как из этого тельца, по успокоении его, образовывалось новое растение, а оболочка клетки не представляла никаких жизненных явлений и погибала, то было несомненно доказано, что П., а не оболочка (как утверждала клеточная теория), составляет самую существенную часть клетки. Аналогичные наблюдения в животном царстве показали, благодаря Келликеру (Kölliker) и Бишофу, значение П. в животных тканях и отсутствие во многих клетках особой оболочки, которая является, таким образом, несущественной. Далее в П. некоторых животных клеток (белых кровяных и лимфатических тельцах) были открыты подобные же движения, как в П. растений, и Ремак (Remak) перенес введенное Молем название П. и на основное вещество животных клеток. Наконец, весьма важными оказались еще раньше сделанные (1835) наблюдения Дюжардэна (Dujardin) над низшими одноклеточными организмами (Protozoa), которые показали несомненно, что эти организмы, лишенные большей частью оболочки, состоят из однородного, зернистого вещества, названного им саркодой. Эти организмы, будучи лишены особых органов, тем не менее передвигаются, захватывают пищу, усваивают ее и размножаются, так что у них все жизненные отправления очевидно должна совершать саркода. Более точные исследования жизненных отправлений этих существ привели ученых к заключению, что саркода низших организмов тожественна с П. растений и животных. На основании приведенного ряда наблюдений, а также и своих собственных, Макс Шульце (Мах Schultze) в 1863 г. подверг строгой критике Шванн-Шлейденевскую клеточную теорию, основав вместо нее новую теорию П., которая, разработанная особенно тщательно в последнее двадцатилетие, стала краеугольным камнем современной зоологии и ботаники. Преобразовав совершенно старую теорию, Шульце сохранил, однако, введенное в науку название клетки, но определил ее как комочек П., одаренный жизненными свойствами. Шульце вполне сознавал, что этот комочек П., который является носителем столь сложных жизненных свойств, совсем не такое простое образование, как выражается этим словом, а должен иметь чрезвычайно сложное строение, которое являлось тогда недоступным вследствие недостаточности оптических средств, что и было подтверждено исследованиями последних лет. Но несмотря на то, что теперь известно ее тончайшее строение, а также и ее различные физические, химические и физиологические свойства, мы и в настоящее время все еще не в состоянии дать точного определения слова П., оттого, что функциональные свойства П., при кажущейся ее однородности, чрезвычайно разнородны. Шульце, доказав тожественность П. растений и животных, имел в виду, конечно, только одинаковость их морфологических и общих физиологических свойств, но отнюдь не тождественность их строения. В действительности, существует столько протоплазм, сколько есть различных существ и органов, выполняющих различные функции. Подобно тому как в органической химии существуют изомерные соединения, возможно, что и в биологии есть изомерия П., обуславливающая, вследствие разнообразия группировки атомов, число и взаимное отношение которых одинаковы, различные свойства и разную жизнедеятельность П. Единственное определение П., которое выдерживает критику, это — основывающееся на отправлениях П., т. е. физиологическое определение, которое и было дано выше. Действительно, самый существенный признак П. — это "быть живой" и утрачивать важнейшие свойства свои вместе с потерей жизни. П. мертвая — больше не П. При настоящем состоянии наших знаний мы должны рассматривать П., невзирая на сложность и разнообразие ее строения, как простое тело, синтез и происхождение которого нам неизвестны.Физические и морфологические свойства. П. представляет вязко-жидкое, почти всегда бесцветное, в больших массах серовато-голубоватое вещество, нерастворимое в воде, преломляющее свет сильнее последней и снабженное подчас двоякой светопреломляемостью. Вопрос, в каком состоянии находится живая П., представляется еще спорным, хотя большинство ученых склонно думать, что состав П. приближается скорее к жидкому, чем к твердому состоянию. Будучи в состоянии покоя предоставлена самой себе, П. принимает форму с поверхностью наименьшего протяжения, что свойственно жидким телам; точно так же и внутренние токи, наблюдаемые в живой П., предполагают подвижность молекул, возможную лишь в жидких веществах. П. обладает довольно большой силой сцепления; опыты над небольшим комочком П. плазмодия миксомицета (см.) показали, что плазмодий выдерживает давление до 80 мг. на квадратный мм., а плазматические нити этого плазмодия оказывают сопротивление разрыву при нагрузке в 120—300 мг. на квадратный мм. П. ресничных инфузорий обладает еще большим сцеплением. В морфологическом отношении П., как важнейшая составная часть клетки, т. е. морфологической единицы живой материи, представляется неразрывной с некоторыми образованиями или включениями, являющимися или продуктами видоизменения, или продуктами формирующей деятельности П. К таким составным частям П. принадлежат: оболочка, ядро, центрозома и различные включения. Под именем оболочки в биологии часто подразумевают два совершенно различных образования: 1) особо видоизмененную (отчасти химически) и уплотненную часть наружного слоя П. или пелликулу (см. Простейшие), имеющую иногда вид тонкой и прозрачной оболочки и встречающуюся в П. каждой клетки, свободно живущей (одноклеточные организмы) или изолированной (яйца, кровяные шарики и т. п.), а также и у большинства входящих в состав многоклеточного (растительного и животного) организма, и 2) настоящую клеточную оболочку, представляющую продукт выделения П. и состоящую из клетчатки, хитина или другого органического вещества; она отсутствует у так называемых голых клеток и достигает особого развития в растительных тканях, где бывает весьма разнообразного строения (см. Клеточка, фиг. 1).Ядро имеет особенно важное значение: находясь в тесной связи и взаимодействии с П. и являясь, как полагают, носителем наследственных качеств, оно представляет необходимую составную часть П., без которой существование живой П. является невозможным (см. Простейшие и Протисты). Относительно величины, числа, формы и химического состава — см. Клеточка. Что же касается его тончайшего строения, то ядру присуще в большинстве случаев такое же, как и П., т. е. мелкоячеистое (см. ниже); лишь в очень немногих случаях оно представляется своеобразным, как, например, пузыревидным (см. Простейшие) или вполне однородным (см. Ресничные инфузории). Ввиду того, что ядро состоит из аналогичного органического вещества (с примесью нуклеинов, пара-нуклеинов и др.), как и П., Страсбургер (Strasburger) предложил различать собственно П. или цитоплазму (κύτος — клетка), т. е. плазматическое содержимое клетки, от нуклеоплазмы или вернее кариоплазмы, (κάρυον — ядро), представляющую плазматическое содержимое ядра.Центрозома или центральное, или полярное тельце, найденное и исследованное точнее в самое последнее время, представляет по своей функции весьма важное образование и по новейшим исследованиям составляет такую же неотъемлемую принадлежность П., как и ядро. Оно предоставляет, так сказать, динамический центр клетки, выступая отчетливо при размножении (делении) клетки и обуславливая изменение строения цитоплазмы, а также и ядра. В состоянии покоя клетки центрозома едва отличима от многочисленных микрозом (см. ниже); величина клетки от 0,0005 до 0,0015 мм; присутствие ее всего легче может быть обнаружено действием особых красок (кислых анилиновых), окрашивающих исключительно ее одну. Она лежит или в самой цитоплазме (всегда вблизи ядра) или в особом, небольшом углублении ядра, т. е. его оболочки или же внутри ядра. Центрозома (см. прилагаемую табл., фиг. 5) собственно состоит из трех частей: 1) самой центрозомы (ц) или маленького, сильно преломляющего свет зернышка, 2) особого участка дифференцированной П., так называемой атракционной сферы или архоплазмы (арх.), окружающей центрозому со всех сторон, и 3) отходящих от последней по всем направлениям лучей или звезды (aster), заметных только при наступлении деления и обуславливающих так называемую лучистость (л) делящейся клетки. Разные включения, встречающиеся в изобилии в П., являются результатом ее формирующей деятельности и представляют или продукт изменения, т. е. образуются в ней и за счет ее, или же продукты обмена веществ. К таким включениям принадлежат: вакуоли, заполненные жидкостью (водой, раствором сахара, соли и т. п.), пузырьки газа, хроматофоры, пиреноиды, стрекательные капсулы, пигментные зернышки, капли слизи и жира, гликогена, зерна крахмала и т. п. (см. Клеточка, Простейшие — соответственные статьи).
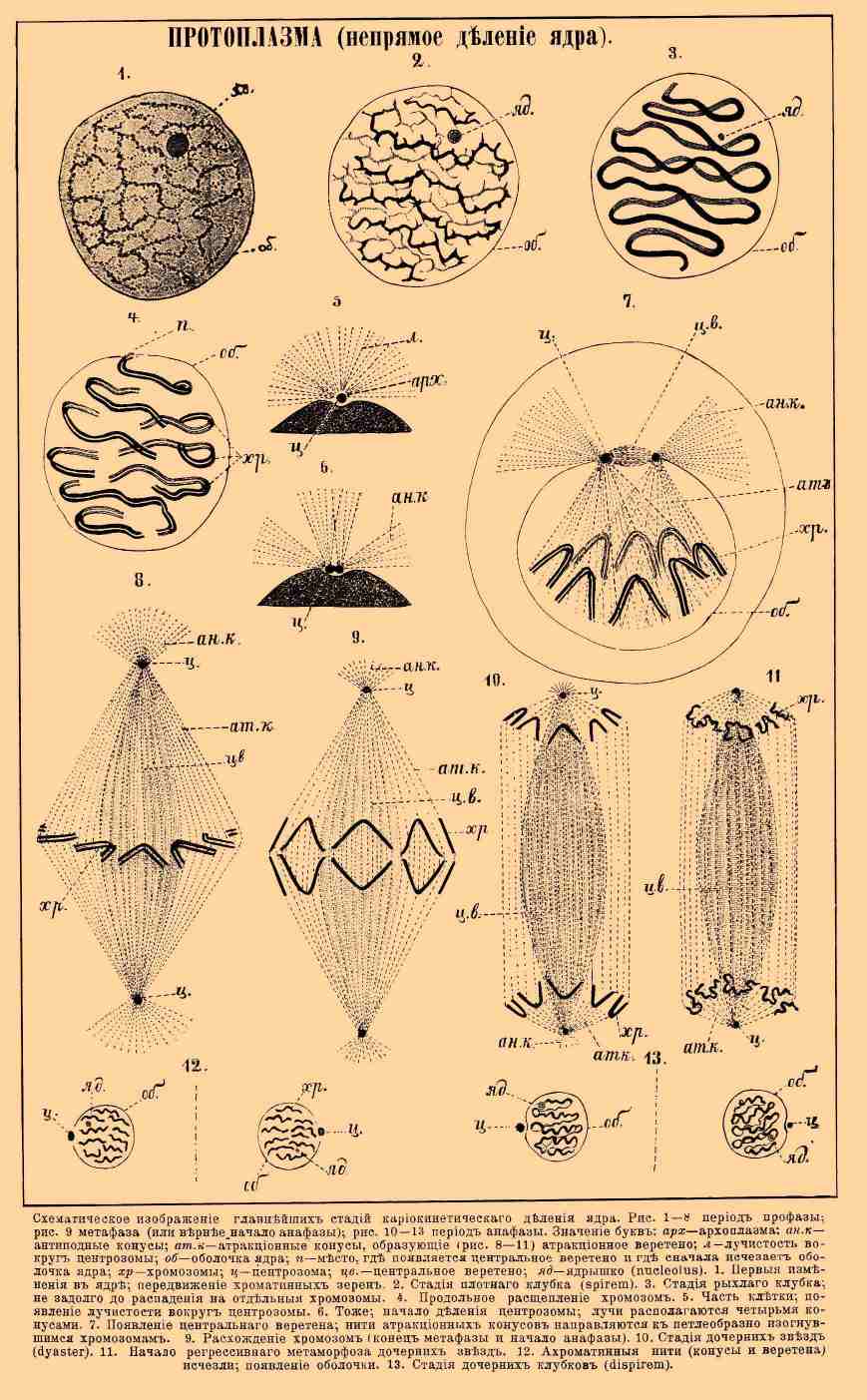 ПРОТОПЛАЗМА (непрямое деление ядра).Схематическое изображение кариокинетического деления ядра. Рис. 1—8. Период профазы; рис. 9. метафаза (или, вернее, начало анафазы); рис. 10—13 период анафазы. Значение букв: арх — архоплазма, ан.к — артиподные конусы, ат.к — атракционные конусы, образующие (рис. 8—11) атракционное веретено, л — лучистость вокруг центросомы, об — оболочка ядра, п — место, где появляется центральное веретено и где сначала исчезает оболочка ядра, хр — хромосомы, ц — центрозома, цв. — центральное веретено, яд — ядрышко (nucleolus). 1. Первые изменения в ядре; передвижение хроматинных ядер. Стадия плотного клубка (spirem). 3. Стадия рыхлого клубка; незадолго до распадения на отдельные хромосомы. 4. Продольное расщепление хромосом. 5. Часть клетки; появление лучистости вокруг центрозомы. 6. То же; начало деления центрозомы; лучи располагаются четырьмя конусами. 7. Появление центрального веретена; нити атракционных конусов направляются к петлеобразно изогнувшимся хромосомам. 9. Расхождение хромосом (конец метафазы и начало анафазы). 10. Стадия дочерних звезд (dyaster). 11. Начало регрессивного метаморфоза дочерних звезд. 12. Ахроматинные нити (конусы и веретена) исчезли; появление оболочки. 13. Стадия дочерних клубков (dispirem).Тончайшее строение П. и главнейшие теории. Биологи до 1865 г. рассматривали П. как однородное, стекловидное, прозрачное вещество, в котором заключены в большем или меньшем количестве мельчайшие зернышки, обуславливающие степень прозрачности П. По мере же усовершенствования микроскопа (в особенности, по изобретении так называемых гомогенных иммерсионных объективов, см.) ученым удалось открыть чрезвычайное сложное тончайшее строение П. Предметом исследования служила как живая П. одноклеточных организмов, яиц и клеток животных и растительных тканей, так и убитая различными реактивами, причем последняя служила в особенности для изучения мельчайших частей, выступающих отчетливо при помощи окраски. Возникшие теории тончайшего строения П. могут быть сведены к четырем основным типам. Упоминая обо всех четырех, разберем подробнее теорию ячеистого строения, так как она объясняет наибольшее число явлений.1) Теория сетчатого или губчатого строения П. была предложена Гейцманном (Heitzmann) в 1873 г.; по этой теории П. или цитоплазма состоит из двух веществ: более плотного губчатого остова, являющегося в оптических или действительных разрезах в виде тонких сетей или переплетающихся перекладин, и жидкого вещества, заполняющего промежутки остова. К остову прилегает бесчисленное количество мелких, плазматических зернышек — микрозом, не играющих никакой существенной роли. Губка с прилегающими многочисленными песчинками, напитанная водой, представляет грубую модель такого строения П. Эта теория нашла многих сторонников; мнения расходятся лишь относительно первенствующего значения той или другой составной части П. Так, Гейцманн вместе с другими приверженцами видит в губчатом остове настоящую живую и деятельную П., а в бесструктурном жидком веществе, лишь инертную, заполняющую массу, тогда как другие сторонники этой теории, с Лейдигом (Leydig) во главе, утверждают, что единственно активным, живым веществом является заполняющая губчатый остов или "спонгиоплазму" жидкость — гиалоплазма.2) Теория фибриллярного или нитчатого строения П. была предложена в 1875 г. Купфером (Kupffer) и впоследствии (1822) подробнее развита Флеммингом (Flemming); по мнению Купфера, П. представляет собой однородное жидкое вещество, в котором залегают более плотные, тонкие нити, сплетением которых обуславливается вид сети. Комок ваты или корпии, напитанный водой, дает грубое представление о таком строении П. Эта теория отличается от предыдущей только тем, что существенная составная часть П. представляется здесь независимыми, самостоятельными нитями, сокращением которых объясняется сократимость П., в общем же обе теории сходны друг с другом, и происхождение сети может легко быть объяснено разветвлением и срастанием отдельных нитей. Описанное строение, хотя и наблюдается в различных клетках животных и растительных тканей, никак не может считаться общим и основным строением П. Строение более жидкой и подвижной П. не совместимо с допущением постоянного плотного состава; П., построенная по этим схемам, не могла бы обнаружить столь распространенных амебоидных движений, встречающихся у одноклеточных организмов, зародышей растений и свободных клеток животных, а также и постоянных токов П. в растительных клетках, одноклеточных организмах и псевдоподиях простейших.3) Теория гранулярного строения П. была намечена уже Арндтом (Arndt), Маджи (Maggi) и Мартэном (Martin), и только в 1886 г. подробнее развита Альтманом (Altmann), которого многие ошибочно считают основателем этой теории. Согласно этой теории, П. представляет собой однородное, бесструктурное жидкое или полужидкое вещество, в котором заключено бесчисленное множество мельчайших крупинок более плотного вещества — гранулы или биобласты. Эти гранулы, являющиеся единственными и действительными живыми элементами П., представляют мельчайшие элементарные организмы или морфологические единицы (отсюда и название биобласты), из которых составлена сама клетка или живая материя. Они питаются, растут, размножаются делением и являются носителями всех качеств П.: в П. они лежат одиночно или скопляются в большие или меньшие комки, или же соединяются рядами в нити или даже сети, но могут вести также и самостоятельную жизнь, и тогда являются в форме бактерий. Основное, по-видимому, бесструктурное вещество, в котором распределены гранулы, по Альтману, может быть, состоит из скопления ультрамикроскопических недоступных нашему зрению гранул. Это основное вещество может быть, более жидкой или боле густой консистенции и, таким образом, вместе с количеством и расположением гранул обуславливать ту или другую консистенцию. Гранулы, плохо заметные в живой П., Альтман проявляет действием различных реактивов (консервирующих жидкостей) и окрасок. Прибегая к грубому сравнению, П., по теории Альтмана, представляет собою род кашицы более густой или более жидкой. Эта теория менее других нашла себе сторонников и хотя и объясняет некоторые жизненные явления П., но против нее можно сделать много возражений. Главный недостаток ее заключается в том, что под именем гранул или биобластов Альтман описывает самые разнообразные включения, не давая признаков, по которым можно было бы отличить биобласта от посторонних включений; другой недостаток, помимо крайней гипотетичности заключений, это самые методы исследования, которые по своей сложности и условности (исследуется не живая П., а умерщвленная определенными реактивами и окрашенная определенными красками) возбуждают часто большое недоверие, так как при таких методах легко получить явления разбухания и другие искусственные образования и осадки. Наконец, бактерии не представляют собою биобластов или отдельных гранул, а, как показали новейшие исследования, такие же клетки, снабженные ядром, как и все прочие одноклеточные организмы.4) Теория ячеистого или пенистого строения П. была впервые предложена Бючли (Bütschli) в 1889 г. и подкреплена опытами и наблюдениями, как его собственными, так и его учеников, Блохмана (Blochmann), Эрлангера (v. Erlanger) и Шевякова, опубликована в окончательном виде в 1892 г., тогда как первое предположение о ячеистом строении было высказано Бючли еще в 1878 г. Согласно этой теории, П. также образована из двух веществ: 1) более плотного, но все же жидкого, состоящего из комбинации белковых частиц и жирных кислот и построенного наподобие маленьких (до 0,001 мм в поперечнике), замкнутых со всех сторон многогранных ячеек, и 2) менее плотного, жидкого вещества, заполняющего полости ячеек. В углах ячеек, а также и вдоль их граней собраны мелкие зернышки или микрозомы, количество, величина и расположение которых весьма разнообразны (фиг. 1 и 2).
ПРОТОПЛАЗМА (непрямое деление ядра).Схематическое изображение кариокинетического деления ядра. Рис. 1—8. Период профазы; рис. 9. метафаза (или, вернее, начало анафазы); рис. 10—13 период анафазы. Значение букв: арх — архоплазма, ан.к — артиподные конусы, ат.к — атракционные конусы, образующие (рис. 8—11) атракционное веретено, л — лучистость вокруг центросомы, об — оболочка ядра, п — место, где появляется центральное веретено и где сначала исчезает оболочка ядра, хр — хромосомы, ц — центрозома, цв. — центральное веретено, яд — ядрышко (nucleolus). 1. Первые изменения в ядре; передвижение хроматинных ядер. Стадия плотного клубка (spirem). 3. Стадия рыхлого клубка; незадолго до распадения на отдельные хромосомы. 4. Продольное расщепление хромосом. 5. Часть клетки; появление лучистости вокруг центрозомы. 6. То же; начало деления центрозомы; лучи располагаются четырьмя конусами. 7. Появление центрального веретена; нити атракционных конусов направляются к петлеобразно изогнувшимся хромосомам. 9. Расхождение хромосом (конец метафазы и начало анафазы). 10. Стадия дочерних звезд (dyaster). 11. Начало регрессивного метаморфоза дочерних звезд. 12. Ахроматинные нити (конусы и веретена) исчезли; появление оболочки. 13. Стадия дочерних клубков (dispirem).Тончайшее строение П. и главнейшие теории. Биологи до 1865 г. рассматривали П. как однородное, стекловидное, прозрачное вещество, в котором заключены в большем или меньшем количестве мельчайшие зернышки, обуславливающие степень прозрачности П. По мере же усовершенствования микроскопа (в особенности, по изобретении так называемых гомогенных иммерсионных объективов, см.) ученым удалось открыть чрезвычайное сложное тончайшее строение П. Предметом исследования служила как живая П. одноклеточных организмов, яиц и клеток животных и растительных тканей, так и убитая различными реактивами, причем последняя служила в особенности для изучения мельчайших частей, выступающих отчетливо при помощи окраски. Возникшие теории тончайшего строения П. могут быть сведены к четырем основным типам. Упоминая обо всех четырех, разберем подробнее теорию ячеистого строения, так как она объясняет наибольшее число явлений.1) Теория сетчатого или губчатого строения П. была предложена Гейцманном (Heitzmann) в 1873 г.; по этой теории П. или цитоплазма состоит из двух веществ: более плотного губчатого остова, являющегося в оптических или действительных разрезах в виде тонких сетей или переплетающихся перекладин, и жидкого вещества, заполняющего промежутки остова. К остову прилегает бесчисленное количество мелких, плазматических зернышек — микрозом, не играющих никакой существенной роли. Губка с прилегающими многочисленными песчинками, напитанная водой, представляет грубую модель такого строения П. Эта теория нашла многих сторонников; мнения расходятся лишь относительно первенствующего значения той или другой составной части П. Так, Гейцманн вместе с другими приверженцами видит в губчатом остове настоящую живую и деятельную П., а в бесструктурном жидком веществе, лишь инертную, заполняющую массу, тогда как другие сторонники этой теории, с Лейдигом (Leydig) во главе, утверждают, что единственно активным, живым веществом является заполняющая губчатый остов или "спонгиоплазму" жидкость — гиалоплазма.2) Теория фибриллярного или нитчатого строения П. была предложена в 1875 г. Купфером (Kupffer) и впоследствии (1822) подробнее развита Флеммингом (Flemming); по мнению Купфера, П. представляет собой однородное жидкое вещество, в котором залегают более плотные, тонкие нити, сплетением которых обуславливается вид сети. Комок ваты или корпии, напитанный водой, дает грубое представление о таком строении П. Эта теория отличается от предыдущей только тем, что существенная составная часть П. представляется здесь независимыми, самостоятельными нитями, сокращением которых объясняется сократимость П., в общем же обе теории сходны друг с другом, и происхождение сети может легко быть объяснено разветвлением и срастанием отдельных нитей. Описанное строение, хотя и наблюдается в различных клетках животных и растительных тканей, никак не может считаться общим и основным строением П. Строение более жидкой и подвижной П. не совместимо с допущением постоянного плотного состава; П., построенная по этим схемам, не могла бы обнаружить столь распространенных амебоидных движений, встречающихся у одноклеточных организмов, зародышей растений и свободных клеток животных, а также и постоянных токов П. в растительных клетках, одноклеточных организмах и псевдоподиях простейших.3) Теория гранулярного строения П. была намечена уже Арндтом (Arndt), Маджи (Maggi) и Мартэном (Martin), и только в 1886 г. подробнее развита Альтманом (Altmann), которого многие ошибочно считают основателем этой теории. Согласно этой теории, П. представляет собой однородное, бесструктурное жидкое или полужидкое вещество, в котором заключено бесчисленное множество мельчайших крупинок более плотного вещества — гранулы или биобласты. Эти гранулы, являющиеся единственными и действительными живыми элементами П., представляют мельчайшие элементарные организмы или морфологические единицы (отсюда и название биобласты), из которых составлена сама клетка или живая материя. Они питаются, растут, размножаются делением и являются носителями всех качеств П.: в П. они лежат одиночно или скопляются в большие или меньшие комки, или же соединяются рядами в нити или даже сети, но могут вести также и самостоятельную жизнь, и тогда являются в форме бактерий. Основное, по-видимому, бесструктурное вещество, в котором распределены гранулы, по Альтману, может быть, состоит из скопления ультрамикроскопических недоступных нашему зрению гранул. Это основное вещество может быть, более жидкой или боле густой консистенции и, таким образом, вместе с количеством и расположением гранул обуславливать ту или другую консистенцию. Гранулы, плохо заметные в живой П., Альтман проявляет действием различных реактивов (консервирующих жидкостей) и окрасок. Прибегая к грубому сравнению, П., по теории Альтмана, представляет собою род кашицы более густой или более жидкой. Эта теория менее других нашла себе сторонников и хотя и объясняет некоторые жизненные явления П., но против нее можно сделать много возражений. Главный недостаток ее заключается в том, что под именем гранул или биобластов Альтман описывает самые разнообразные включения, не давая признаков, по которым можно было бы отличить биобласта от посторонних включений; другой недостаток, помимо крайней гипотетичности заключений, это самые методы исследования, которые по своей сложности и условности (исследуется не живая П., а умерщвленная определенными реактивами и окрашенная определенными красками) возбуждают часто большое недоверие, так как при таких методах легко получить явления разбухания и другие искусственные образования и осадки. Наконец, бактерии не представляют собою биобластов или отдельных гранул, а, как показали новейшие исследования, такие же клетки, снабженные ядром, как и все прочие одноклеточные организмы.4) Теория ячеистого или пенистого строения П. была впервые предложена Бючли (Bütschli) в 1889 г. и подкреплена опытами и наблюдениями, как его собственными, так и его учеников, Блохмана (Blochmann), Эрлангера (v. Erlanger) и Шевякова, опубликована в окончательном виде в 1892 г., тогда как первое предположение о ячеистом строении было высказано Бючли еще в 1878 г. Согласно этой теории, П. также образована из двух веществ: 1) более плотного, но все же жидкого, состоящего из комбинации белковых частиц и жирных кислот и построенного наподобие маленьких (до 0,001 мм в поперечнике), замкнутых со всех сторон многогранных ячеек, и 2) менее плотного, жидкого вещества, заполняющего полости ячеек. В углах ячеек, а также и вдоль их граней собраны мелкие зернышки или микрозомы, количество, величина и расположение которых весьма разнообразны (фиг. 1 и 2).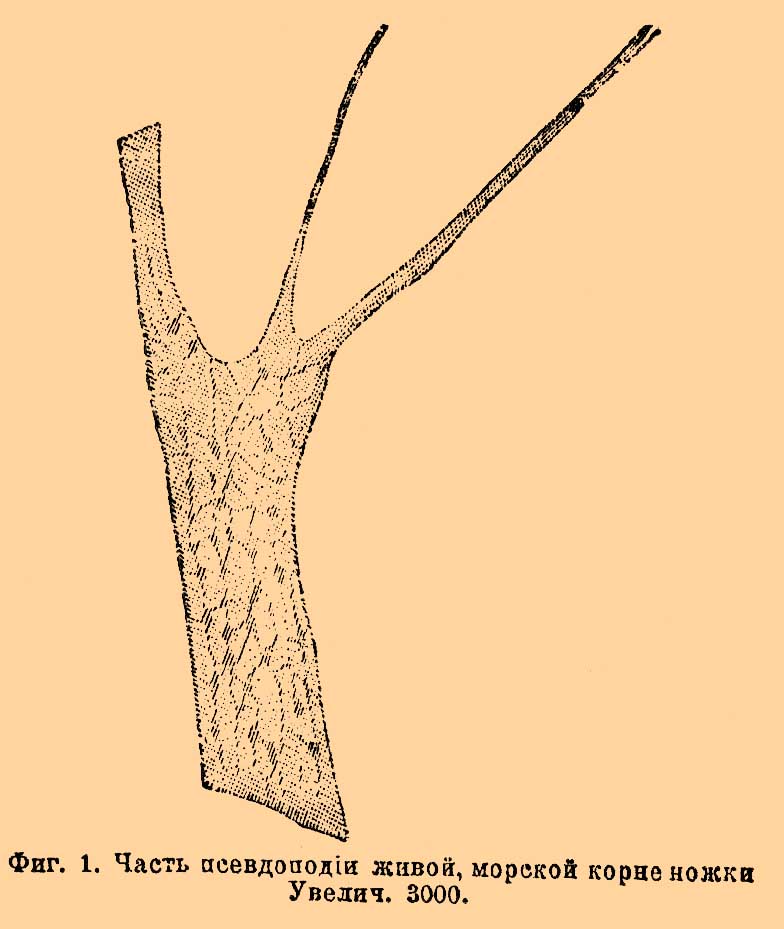 Фиг. 1. Часть псевдоподии живой, морской корненожки. Увеличение 3000.
Фиг. 1. Часть псевдоподии живой, морской корненожки. Увеличение 3000. Фиг. 2. Два живых протоплазматических отростка из волосяной клетки растения мальва. Увеличение 3000. Фиг. 3. Часть поверхности тела живой ресничной инфузории в оптическом разрезе: p — пелликула; alv — альвеолярный слой: v — сократительная вакуоль. Увеличение 2500.Эти многогранные ячейки располагаются таким образом, что в одном ребре всегда сходятся три пластинки, так что в оптическом разрезе в каждой узловой точке сходятся всего только три линии. В громадном большинстве случаев в П. можно заметить поверхностный слой, в котором ячейки расположены своеобразно, так, что перегородки их стоят перпендикулярно к наружной поверхности и потому на оптическом разрезе представляются параллельными — это так называемый альвеолярный слой П. (фиг. 3 и 4).
Фиг. 2. Два живых протоплазматических отростка из волосяной клетки растения мальва. Увеличение 3000. Фиг. 3. Часть поверхности тела живой ресничной инфузории в оптическом разрезе: p — пелликула; alv — альвеолярный слой: v — сократительная вакуоль. Увеличение 2500.Эти многогранные ячейки располагаются таким образом, что в одном ребре всегда сходятся три пластинки, так что в оптическом разрезе в каждой узловой точке сходятся всего только три линии. В громадном большинстве случаев в П. можно заметить поверхностный слой, в котором ячейки расположены своеобразно, так, что перегородки их стоят перпендикулярно к наружной поверхности и потому на оптическом разрезе представляются параллельными — это так называемый альвеолярный слой П. (фиг. 3 и 4). Фиг. 4. Тонкий разрез через яйцо морского ежа. alv — альвеолярный слой. Увеличение 2500.Пчелиные соты, состоящие из восковых ячеек, заключающих мед, или лучше мыльная пена — могут служить наглядными примерами описанного строения П., только при втором примере мы должны вообразить себе, что в ячейках находится не воздух, а другая жидкость. Наблюдения производились преимущественно над живой П. одноклеточных животных, яиц, кровяных и лимфатических телец, одно- и многоклеточных растений, а также и над умерщвленной и окрашенной П. различных одноклеточных организмов и самых разнообразных животных и растительных тканей. На всех этих объектах удалось установить ячеистое строение П., причем в тех случаях, где П. на первый взгляд казалась однородной, при более внимательном изучении (с помощью самых сильных систем), а также при надлежащей консервировке и окраске, удавалось все-таки обнаружить ячеистое строение. На основании этих наблюдений, производимых в течение с лишком двадцати лет, Бючли пришел к заключению, что каждая П. имеет ячеистое строение, хотя последнее часто настолько тонко, что может быть обнаружено лишь с большим трудом, а в некоторых случаях и совершенно ускользает от наблюдения. Кажущееся нитчатое строение зависит от того, что ячейки сильно вытягиваются по одному направлению, и их продольные грани развиты сильнее. Тем же объясняется и сетчатое или губчатое строение. Следует заметить при этом, что в оптическом и действительном разрезе ячеистое строение также представляется нам в виде сетей, и часто требуется много внимания и труда, чтобы решить, имеем ли мы дело с губчатым или ячеистым строением; тем не менее это, во всяком случае, возможно. Бючли, желая подкрепить свои наблюдения опытами, задался целью приготовить микроскопически тончайшую, несомненную пену из двух жидкостей и, рассматривая ее под микроскопом, сравнить с живой П. Опыты эти дали поразительные результаты. Растирая в агатовой ступке истертый в мельчайший порошок поташ, тростниковый сахар или поваренную соль с долго стоявшим, загустевшим прованским маслом, он получал густую, гомогенную массу, из которой приготовлял очень маленькие шарики в 1/4—1 мм. в диаметре. Эти шарики помещались на покровное стеклышко, снабженное на углах восковыми ножками, которое накладывалось на каплю смеси воды и глицерина, расположенную на предметном стеклышке. Спустя некоторое время жидкость начинала проникать в шарик и, диффундируя через тонкие слои масла, растворяла мельчайшие частички твердого вещества, т. е. поташа, соли или сахара, вследствие чего гомогенные шарики наполнялись пузырьками или вакуолями, которые, благодаря взаимному давлению, принимали многогранную, ячеистую форму. Таким образом шарики, в конце концов, превращались в тончайшую пенистую массу или "пенистые капли" (Schaumtropfen), в которых стенки ячеек состояли из масла, а содержимое ячеек — из жидкого раствора поташа, соли или сахара в смеси воды с глицерином. Эти пенистые капли под микроскопом при сильных увеличениях представляют типичное ячеистое строение с явственным наружным альвеолярным слоем, характерное для живой П. Размер ячеек (до 0,001 мм) искусственной пенистой капли соответствует размеру ячеек П. Эти комочки или капли принимают, обыкновенно, неправильную форму и до такой степени похожи на некоторые живые одноклеточные организмы, что, рассматривая небольшой участок этих комочков или капель, даже опытный микроскопист и протистолог затруднится иногда отличить их от одноклеточных существ (фиг. 5).
Фиг. 4. Тонкий разрез через яйцо морского ежа. alv — альвеолярный слой. Увеличение 2500.Пчелиные соты, состоящие из восковых ячеек, заключающих мед, или лучше мыльная пена — могут служить наглядными примерами описанного строения П., только при втором примере мы должны вообразить себе, что в ячейках находится не воздух, а другая жидкость. Наблюдения производились преимущественно над живой П. одноклеточных животных, яиц, кровяных и лимфатических телец, одно- и многоклеточных растений, а также и над умерщвленной и окрашенной П. различных одноклеточных организмов и самых разнообразных животных и растительных тканей. На всех этих объектах удалось установить ячеистое строение П., причем в тех случаях, где П. на первый взгляд казалась однородной, при более внимательном изучении (с помощью самых сильных систем), а также при надлежащей консервировке и окраске, удавалось все-таки обнаружить ячеистое строение. На основании этих наблюдений, производимых в течение с лишком двадцати лет, Бючли пришел к заключению, что каждая П. имеет ячеистое строение, хотя последнее часто настолько тонко, что может быть обнаружено лишь с большим трудом, а в некоторых случаях и совершенно ускользает от наблюдения. Кажущееся нитчатое строение зависит от того, что ячейки сильно вытягиваются по одному направлению, и их продольные грани развиты сильнее. Тем же объясняется и сетчатое или губчатое строение. Следует заметить при этом, что в оптическом и действительном разрезе ячеистое строение также представляется нам в виде сетей, и часто требуется много внимания и труда, чтобы решить, имеем ли мы дело с губчатым или ячеистым строением; тем не менее это, во всяком случае, возможно. Бючли, желая подкрепить свои наблюдения опытами, задался целью приготовить микроскопически тончайшую, несомненную пену из двух жидкостей и, рассматривая ее под микроскопом, сравнить с живой П. Опыты эти дали поразительные результаты. Растирая в агатовой ступке истертый в мельчайший порошок поташ, тростниковый сахар или поваренную соль с долго стоявшим, загустевшим прованским маслом, он получал густую, гомогенную массу, из которой приготовлял очень маленькие шарики в 1/4—1 мм. в диаметре. Эти шарики помещались на покровное стеклышко, снабженное на углах восковыми ножками, которое накладывалось на каплю смеси воды и глицерина, расположенную на предметном стеклышке. Спустя некоторое время жидкость начинала проникать в шарик и, диффундируя через тонкие слои масла, растворяла мельчайшие частички твердого вещества, т. е. поташа, соли или сахара, вследствие чего гомогенные шарики наполнялись пузырьками или вакуолями, которые, благодаря взаимному давлению, принимали многогранную, ячеистую форму. Таким образом шарики, в конце концов, превращались в тончайшую пенистую массу или "пенистые капли" (Schaumtropfen), в которых стенки ячеек состояли из масла, а содержимое ячеек — из жидкого раствора поташа, соли или сахара в смеси воды с глицерином. Эти пенистые капли под микроскопом при сильных увеличениях представляют типичное ячеистое строение с явственным наружным альвеолярным слоем, характерное для живой П. Размер ячеек (до 0,001 мм) искусственной пенистой капли соответствует размеру ячеек П. Эти комочки или капли принимают, обыкновенно, неправильную форму и до такой степени похожи на некоторые живые одноклеточные организмы, что, рассматривая небольшой участок этих комочков или капель, даже опытный микроскопист и протистолог затруднится иногда отличить их от одноклеточных существ (фиг. 5). Фиг. 5. Оптический разрез через краевую часть искусственной пенистой капли (из оливкового масла и поваренной соли). alv — альвеолярный слой. Увеличение 1250.Все это Бючли справедливо считает окончательным доказательством в пользу своей теории ячеистого строения П. Примешанные к описанным комочкам, при приготовлении их, т. е. во время растирания в ступке, мельчайшие твердые частички (как, например, сажа), располагаются вдоль граней и, главным образом, в узловых точках системы ячеек пенистых капель, как и настоящие микрозомы живой П. Под микроскопом в таких искусственных пенистых каплях, особенно содержащих крупинки сажи, можно заметить восходящие и нисходящие внутренние токи, совершенно подобные тем, какие наблюдаются в живой П. некоторых одноклеточных организмов и растительных клетках, и о которых мы будем говорить ниже при разборе физиологических свойств протоплазмы. Мало того, если эти внутренние токи происходят более или менее продолжительно в одном направлении и достаточно интенсивны, то результатом их является передвижение самих искусственных пенистых капель, которые могут двигаться самопроизвольно, подобно типичным живым амебам (при помощи так называемых псевдоподий), по разным направлениям (постоянно изменяя его), более или менее продолжительное время (до 7 суток). Относительно гипотетичной причины этого движении будет сказано ниже (см. физиологические свойства П.); заметим здесь только, что аналогичные движения одноклеточных организмов (а на них сводятся вообще все движения, обнаруживаемые также и высшими животными) допускают подобное же объяснение, если ввести некоторые поправки, принимая во внимание химическую сторону явлений.Помимо изложенных теорий, построенных с помощью и на основании непосредственных микроскопических наблюдений, существуют еще теории, построенные умозрительным путем и касающиеся причины и сущности жизни, появления живого вещества, наследственности и тому подобных общих вопросов биологии — это так называемые философские теории биологии, которых мы здесь касаться не будем (см. соответственные статьи).Химические свойства П. При настоящем положении наших знаний мы даже не можем с уверенностью сказать, есть ли П. определенное химическое тело или изменяющаяся смесь различных химических тел, так как П. умирает, как только ее подвергают действию реактивов, употребляемых в аналитической химии, а, умирая, перестает быть настоящей П., теряя часть своих характерных свойств. Дальнейшее затруднение представляет еще то обстоятельство, что при анализе П. подчас невозможно отделить различные посторонние тела и включения П., представляющие продукты обмена веществ. Известно только, что в состав П. входят: углерод, водород, азот, кислород, сера, фосфор, а также фтор, хлор, кремний, натрий, калий, кальций, магний и железо. Первые из этих элементов (т. е. С, H, N, О, S и Р) образуют очень сложные и малоизученные соединения — так называемые белковые или протеиновые тела, представляющие при значительном процентном (60 до 9%) содержании воды главную составную часть П., тогда как последние входят в состав различных солей, встречающихся в П. Возникшее в последнее время с такой силой изучение морфологических свойств, т. е. тончайшего строения П., а также и изменений, происходящих в делящемся ядре и окружающей П., привели к более внимательному и точному изучению химических свойств вновь открытых морфологических частей, составляющих П. Так, в настоящее время, мы различаем очень много веществ, входящих в состав П., из коих некоторые, как, например, линин, пиренин, парануклеин и др., нам совершенно неизвестны с химической стороны: мы не знаем ни их химической формулы, ни химического состава, ни даже представляют ли они простые соединения или смесь разных соединений — нам известно только их отношение к различным красящим веществам, а также их некоторые оптические и физические свойства. Зато другие соединения удалось изолировать и изучить отчасти их химическое строение, а отчасти только их отношение к различным химическим реактивам. В общем же, белковые вещества (по Шюценбергеру формула альбумина C60H100N16O20) характеризуются очень большим молекулярным весом и очень сложным строением частицы. В состав цитоплазмы входят: 1) пластин, составляющий самую существенную часть ее и который не удалось еще изолировать; он нерастворим в поваренной соли, сернокислом магнии и главное — в пепсине и трипсине, растворим в слабой уксусной кислоте, не окрашивается основными анилиновыми красками, а только кислыми. 2) Нуклео-альбумины — белковые, содержащие фосфор вещества, растворимые в желудочном соке, разлагающем их на пептоны, остающиеся в растворе, и выпадающую нуклеинную кислоту. 3) Глобулины — белковые тела, не содержащие фосфора, нерастворимые в воде, но растворимые в разбавленных растворах (5—10%) солей. 4) Лецитины (очень сложная химическая формула) — представляют сочетание холина (C5H15NO2) с жирами, заключающими фосфорную кислоту, растворимы в алкоголе. 5) Холестерин (C26H43OH) — непредельный одноатомный алкоголь, растворимый в эфире и кипящем спирте. 6) Фосфорнокислые и хлористые соли калия, натрия, магния и кальция, а также железо в соединении с нуклео-альбуминами. Химический состав в тонких (клеточных) оболочках неизвестен, тогда как в более толстых главными составными частями являются: хитин (С15H26N2O10), кератин, клетчатка или целлюлоза (C6H10O5) и др. В состав ядра входят: хроматин, состоящий из холестерина и нуклеина, богатого фосфором и нерастворимого в желудочном соке вещества; пиренин и линин или парахроматин, по-видимому, представляют соединения нуклео-альбуминов с пластином. Из общих химических свойств живой П. следует упомянуть ее щелочную реакцию (причем сок некоторых растительных клеток может быть кислым) и способность прижизненной окраски, т. е. окрашиваться слабыми растворами нейтральных анилиновых красок.Физиологические свойства П. Самыми характерными свойствами живой П. являются способность к движению и раздражимость, которые, в свою очередь, представляют лишь приготовительные условия для питания и проистекающих из последнего явлений роста и размножения. Движение, обнаруживаемое П., бывает двоякое. 1) Движение внутри П., выражающееся в перемещении зернышек и различных других включений — это так называемые внутренние токи, и 2) движение, выражающееся в изменении наружной формы и в перемене места самой П. — это так называемые амёбоидные и локомоторные движения. Первый род движения, без сомнения, свойственен всякой живой П., но в большинстве случаев движение это происходит настолько медленно, что ускользает от нашего непосредственного наблюдения. Всего энергичнее оно происходит в клетках растений и в некоторых простейших, но было наблюдаемо также и в животных клетках. Сущность этого движения заключается в том, что от любой точки поверхности комка П. или клетки отходит ток, идущий внутрь и направляющийся по оси (совпадающей, большей частью, с продольной осью комка П. или клетки) комка П.; достигнув противоположной точки поверхности, восходящий осевой ток переходит в нисходящие токи, идущие параллельно поверхности комка и собирающиеся в точке отправления восходящего тока. Кроме того, наблюдается в некоторых случаях еще движение в стенкоположной (т. е. наружной) П. клеток, и всегда в одном направлении, так называемое вращательное движение или rotatio (см. Клеточка). Амёбоидное движение, заключающееся в образовании в любом или определенном месте поверхности П. небольших пальчатых или нитевидных псевдоподий или ложноножек, быстро выпячиваемых и втягиваемых обратно в тело, есть следствие внутренних токов, продолжающихся более или менее долго и с достаточной силой в одном направлении, и если наружный слой П. не настолько уплотнен, чтобы мог препятствовать образованию таких отростков. Результатом образования таких псевдоподий — является поступательное движение или перемещение П. организма (простейшие) или клетки (яйца многоклеточных животных, лимфатические тельца, белые кровяные шарики позвоночных и т. п.). В простейшем случае псевдоподии представляются в виде тупых, простых или разветвленных отростков и, по мере уплотнения наружного слоя (см. Простейшие) становятся тонкими, нитевидными (образующими подчас сетчатые сплетения), на морфологической подкладке которых развиваются при дальнейшем уплотнении П. жгутики и реснички, т. е. особые органы передвижения одноклеточных организмов, встречающиеся также и у клеток некоторых тканей многоклеточных животных (наружные мерцательные покровы низших червей и разных личинок, мерцающий эпителий кишечника и др.). В этих псевдоподиях, при сильных увеличениях, заметно движение заключающихся в них зернышек, которое даже в тончайших нитях происходит в двух противоположных направлениях одновременно. Причина разнообразных движений П. остается пока невыясненной; наиболее заслуживающей внимания является гипотеза Бючли. Приготовленные им искусственным путем (см. выше) пенистые капли обнаруживают не только характерные внутренние токи, но и перемещаются более или менее быстро в разных направлениях при помощи псевдоподий наподобие настоящих амёб. При нагревании эти движения становятся быстрее, а при пропускании гальванического (индукционного) тока пенистые капли направляются к катоду. Это движение Бючли объясняет следующим образом. Представим себе, что в любом месте поверхности пенистой капли разорвутся или лопнут по какой-либо причине несколько ячеек, вследствие чего мыльный раствор вытечет из них наружу и распространится в этой точке по поверхности. Но поверхностное натяжение этой жидкости в воде меньше поверхностного натяжения масла (образующего стенки ячеек), поэтому в данном месте образуется небольшое возвышение, причем поверхностные ячейки подвигаются вперед и освобождают позади себя место, которое заполняется надвигающимися позади лежащими ячейками и так далее. На поверхности разрываются новые ячейки, и явление продолжается, вызывая полное перемещение ячеек и передвижение всей пенистой массы. Бючли полагает, что и тожественные явления движения амёбы должны быть сведены к той же причине, т. е. чисто механической. В П. наблюдаются еще особые и своеобразные движения во время размножения, направленные к разделению и новой группировке различных существенных элементов, входящих в состав П., т. е. ее цитоплазмы и, главное, — ядра. Наконец, различают еще перемещения П. и отдельных организмов в определенном направлении, обуславливаемые различными физическими, химическими или физиологическими агентами, как то: тяжестью, светом, теплотой, кислородом, углекислотой, питательными веществами и т. п.; это так называемый геотаксис, фототаксис, термотаксис, хемотаксис и т. д. (см. Тропизм и Таксис), направляющие движение или к источнику раздражения (положительный таксис), или в противоположную сторону (отрицательный).Раздражимость. — Особое свойство, присущее лишь живой П., отвечать тем или иным способом на самые разнообразные влияния окружающей среды. Эта раздражимость проявляется специфическими реакциями в зависимости от самой природы П., а способ, которым последняя реагирует, называется эффектом раздражения. Одно и то же раздражение может вызвать разные эффекты в различных П. или организмах, так, например, свет притягивает одни и отталкивает другие организмы. Весьма различные раздражители могут также вызывать в той же П. одинаковый эффект раздражения, что называется специфической энергией; так, например, мускульная клетка на всякое раздражение отвечает сокращением. Эти явления зависят от особого и разнообразного строения самой П. Таким образом, хотя раздражимость и есть основное свойство живой П., но проявляется она сообразно с особенностями строения протоплазмы. Все раздражители могут быть сведены к трем категориям. 1) Механические, как то: прикосновение, сотрясение, сжатие, удар и колебания; 2) физические, как то: тяжесть, свет, теплота и электричество; 3) химические, т. е. всякие химические вещества. Механические раздражения заставляют П. сокращаться, втягивать псевдоподии, останавливаться и принимать шаровидную форму (например, амёбу), причем слабые раздражения действуют только на непосредственно затронутое место, тогда как более сильные распространяются на большие расстояния. Световые раздражения вызывают характерные изменения формы в П., а также и движения в определенном направлении у свободно подвижных одноклеточных организмов. При действии света псевдоподии некоторых корненожек и миксомицет втягиваются и вытягиваются вновь в темноте, а из многих одноклеточных организмов, снабженных ресничками и жгутиками, одни собираются на обращенной к свету стороне сосуда, а другие на противоположной (положительная и отрицательная фототаксия). Не меньшее действие оказывает свет на П. хроматофоров или зерен хлорофилла растений, заставляя их вращаться или перемещаться или даже менять свою форму. Наконец, пигментные клетки и хроматофоры животных весьма сильно реагируют на свет и, изменяя свою величину и форму, обуславливают изменение окраски рыб, амфибий, головоногих моллюсков и др. Термические раздражения играют еще более заметную роль в жизни П. Для всякой П. существуют пределы температурные (вне которых наступает смерть П.), весьма разнообразные, в особенности же верхний (40°—130° Ц.). До достижения предельных температур наступает явление, называемое оцепенением; П. при известной температуре не обнаруживает более жизненных явлений (например, движения), но проявляет их, как только температура изменится. Термические раздражения резче всего вызываются в возрастании скорости внутренних токов или перемещений по мере возвышения температуры; причем — максимум скорости составляет оптимум температуры, лежащий несколькими градусами ниже предельной температуры, при которой наступает оцепенение от тепла. Электрические раздражения проявляются при действии гальванических токов, индукционных и постоянных, на П. лишь в том случае, если непосредственно проходят по ней. При действии слабых индукционных ударов движение зернышек (внутренние токи) в П. останавливается, а амёбы и белые кровяные шарики на короткое время прекращают свое движение. От сильных ударов псевдоподии втягиваются, движения прекращаются, организмы съеживаются в шар, лопаются и разрушаются. Постоянный ток (слабый), пропускаемый через одноклеточные организмы, побуждает их к движению в определенном направлении (гальванотропизм), причем некоторые во время замыкания тока покидают анод и направляются к катоду, где и скопляются, а по размыкании тока направляются к аноду, тогда как другие, обратно, при замыкании тока направляются к аноду, а при размыкании — к катоду (отрицательный и положительный гальванотропизм). Наконец, химические раздражители весьма разнообразны как по своему составу, так и действию, оказывая или всестороннее влияние на П. (анестезирующие вещества), или в одном определенном направлении (см. Хемотаксис и Хемотропизм). В общем же следует заметить, что П. может до известной степени приспособляться к химическим изменениям окружающей среды (соленость воды, растворы, присутствие и обилие кислорода, углекислоты, сероводорода и т. д.), если только эти изменения совершаются постепенно, причем П. претерпевает некоторые изменения в своем строении и жизненной деятельности.Формирующая деятельность П. является следствием раздражимости и способности к движению. В этой деятельности следует различать три стадии: поглощение, ассимиляцию или превращение и выделение. Посторонние вещества могут быть поглощаемы П. извне в газообразном, жидком и твердом состояниях: в первых двух — всякой П., а в последнем — лишь П. одноклеточных организмов и некоторых клеток многоклеточных животных. Из газообразных веществ наиболее важны кислород и углекислота. Поглощение кислорода, называемое дыханием, имеет особо важное значение для процессов обмена веществ, так как благодаря ему в П. образуются соединения более богатые кислородом, дальнейшим расщеплением и окислением которых получаются вода и углекислота. При этих химических реакциях, составляющих "регрессивный метаморфоз" или разрушение органического вещества, развивается теплота, являющаяся источником живых сил. Поглощение углекислоты, присущее только растительному царству и некоторым простейшим (см.), ведет к так называемому прогрессивному метаморфозу, т. е. к синтезу органических соединений с высокой частицей. При этом процессе восстановления углекислоты потребляется теплота, которая связывается во вновь образованном веществе в виде потенциальной энергии. Поглощение П. жидких веществ представляет главный источник питания как для растений, так и для животных. Первые заимствуют питательные вещества в растворенном виде или из воды, или же из влажной почвы при помощи корней, тогда как клетки животных получают их из жидкостей (пищевая кашица, хилус, лимфа, кровь), приготовляемых сложным путем в теле самих животных. При таком поглощении П. различных клеток оказывает предпочтение тому или другому веществу, что и называется избирательной способностью, причина которой еще не выяснена, но, по всей вероятности, сводится к химическому сродству разных веществ, встречающихся в П. постоянно или временно возникающих при обмене веществ. Поглощение твердых тел П. присуще лишь некоторым одноклеточным организмам (см. Простейшие, питание) и некоторым тканевым клеткам многоклеточных животных (см. Фагоцитоз). Вещества, принятые П., перерабатываются в П. и превращаются в многочисленные и самые разнообразные вещества. Химическая сторона этих превращений остается еще большей частью совершенно темной. Известно, например, только, что встречающийся в растениях сахар и крахмал образуются синтетическим путем, но едва ли прямым синтезом углерода и воды, а жирные масла — превращением углеводов, возникших через посредство хлорофилла и т. д. Далее, что превращение углеводов, жиров и белков из пассивного состояния в активное происходит при помощи так называемых ферментов (см.): так, диастаза превращает крахмал в сахар, инвертин расщепляет тростниковый сахар на декстрозу и левулозу, птиалин (фермент слюны) превращает крахмал в декстрин и виноградный сахар и т. д., что белки для дальнейшего потребления пептонизируются, а жиры расщепляются на глицерин и жирные кислоты и т. д. (см. Питание растений и Пищеварение). Не более выяснена роль П. при превращении веществ: так, одни предполагают, что различные вещества получаются превращением самой П. путем химических превращений и расщеплений ее частиц, а другие, что они образуются из пластических веществ, углеводов, жиров и пр., воспринятых П. при обмене веществ. Лучше известна морфологическая сторона превращений, т. е. результаты или продукты обмена веществ, которые отлагаются внутри П. или на ее поверхности в бесформенном виде или в виде так называемых включений, отличимых оптически. Сюда относятся вакуоли, жир, гликоген, муциген, крахмал, пигментные зернышки, кристаллы гуанина и разных солей, скелеты, разные оболочки и т. п. Наконец, выделение или удаление из П. негодных продуктов обмена веществ завершает процесс формирующей деятельности П. Эти непригодные П. или организму вещества выводятся наружу также в трояком состоянии, и в твердом лишь исключительно некоторыми простейшими и клетками Metazoa. В газообразном виде выводится углекислота (при дыхательных процессах), кислород (растениями) и водяные пары, а также и другие различные газы, а в жидком состоянии — моча, мочевая кислота, гиппуровая кислота, гуанин и т. п. Главным и непосредственным результатом формирующей деятельности П. является образование новой живой П., которая, по мнению Бючли, образуется в виде маленьких ячеек, возникающих между прежними ячейками, т. е. в их узловых точках, и постепенно увеличивающихся в объеме. Конечный результат таких новообразований выражается в росте самой П.Размножение. Рост П., рассматриваемой приуроченной к клетке, не может, однако, продолжаться беспредельно. Всякая клетка имеет свою предельную величину, по достижении которой наступает размножение, которое поэтому может быть названо ростом за пределы индивидуальности. Каждая новая клетка образуется делением старой, точно так же, как каждое ядро — делением другого ядра. Деление может быть прямое или амитотическое и непрямое или кариокинетическое или митотическое. При непрямом [Хотя сущность кариокинеза изложена уже в ст. Клеточка, но за последние три года было сделано столько крупных открытий, существенно изменяющих и дополняющих общую картину этого важного для биологи процесса, что мы излагаем его вкратце вновь, согласно новейшему учению.] делении, называемом также кариокинезом (κάρυον — ядро, κίνησις — движение) или митозом (μίτος — нить) происходят крупные изменения, как в цитоплазме, так и в ядре, появляющиеся одновременно. Мы различаем, собственно, три периода: профазу — изменения в материнском ядре, метафазу — само деление и анафазу или изменения, происходящие в дочерних ядрах. Изменения, происходящие во время профазы в ядре, выражаются в том, что хроматинные зернышки, залегающие в стадии покоя ядра в узловых точках ахроматичных стенок ячеек, начинают передвигаться (табл., фиг. 1) и располагаются порядно, образуя в совокупности волнистые нити, между которыми заметны еще ахроматинные стенки ячеек (табл., фиг. 2) — стадии плотного клубка (spirem). Различают или одну сплошную хроматинную нить (табл., фиг. 3) в ядре, или несколько отрезков. Затем хроматинные нити начинают укорачиваться, причем они увеличиваются в толщине, их зазубренные края сглаживаются (табл., фиг. 3), и если была одна сложная хроматинная нить, то она вскоре распадается на несколько (всегда определенное и большей частью четное число) отрезков — стадия рыхлого клубка. Эти отрезки, называемые хромосомами (χρώμα — цвет, σώμα — тело), имеют весьма разнообразную форму (нитей, крючков, палочек и шариков) и вскоре расщепляются в продольном направлении надвое (табл., фиг. 4 — хр), причем обе половины остаются вместе. К концу этого процесса исчезает находящееся в ядре ядрышко (табл., фиг. 1—3 — яд); по мнению других, оно у некоторых клеток дает центрозому, а у большинства (за исключением всех простейших) клеток и оболочку ядра (табл., фиг. 1—4 — об). В это же время происходят изменения и в окружающей ядро цитоплазме, а именно от центрозомы (табл., фиг. 5 — ц), окруженной архоплазмой или атракционной сферой (арх) и залегающей в состоянии покоя вблизи ядра или в небольшом углублении последнего, начинают отходить по всем направлениям тонкие плазматические лучи, обуславливающие впоследствии лучистость цитоплазмы (табл., фиг. 5 — л). Центрозома вытягивается в длину, принимает бисквитообразную форму и делится на две (табл., фиг. 6 — ц), из коих каждая окружена архоплазмой. По мере расхождения обеих центрозом, между ними выступает зачаток веретена, разрастающегося впоследствии (при дальнейшем расхождении центрозом), в так называемое центральное веретено (табл., фиг. 7 — ц. в), состоящее из тонких ахроматических нитей, идущих от одной центрозомы (ц) к другой и образующих в совокупности два конуса, соединенных своими основаниями. Во время расхождения центрозом начинает растворяться оболочка ядра (табл., фиг. 4 — об) и исчезает сначала в месте ближайшем к центральному веретену (табл., фиг. 4 — п. и фиг. 7), а затем и в других местах. Центрозомы, расходясь, т. е. удаляясь друг от друга, вытягивают в длину центральное веретено и располагаются, наконец, на двух противоположных точках ядра (табл., фиг. 8 — ц). В продолжение всего описанного процесса плазматические нити или лучи, отходящие от центрозом, группируются около каждой центрозомы в виде двух расходящихся снопов лучей или конусов (табл., фиг. 7), так называемых антиподных (ан. к.) и атракционных (ат. к.) конусов. Первые впоследствии (при окончательном расхождении центрозом) направляются в сторону, диаметрально противоположную от направления ахроматинного центрального веретена (табл., фиг. 8 — ан. к.), а последние (табл., фиг. 8 — ат. к.) направляются к хромосомам (хр) ядра таким образом, что к каждой хромосоме от каждой центрозомы направляются по одной плазматической нити. Эти нити, совместно с образовавшимся расхождением центрозом центральным веретеном (ц. в.), оказывают направляющее действие на хромосомы, которые к концу этой стадии (табл., фиг. 8) располагаются в одной плоскости, перпендикулярной к оси ахроматинного центрального веретена (полюса которого составляют центрозомы), снаружи или вокруг последнего. Они перегибаются при этом наподобие петель (табл., фиг. 7 и 8 — хр) или крючков, причем вершины этих петель обращены к центру ядра, и к ним с двух сторон подходят тонкие нити, идущие от центрозом и образующие в общей сложности два конуса — атракционных конуса (табл., рис. 8 — ат. к.), соединенных своими основаниями. Эти два конуса, образуют второе ахроматинное веретено, как бы одевающее центральное, и которое в отличие от последнего называется направляющим или атракционным веретеном. Этим процессом заканчивается профаза деления, и получившаяся стадия ядра называется звездой (aster); наступающая затем метафаза продолжается очень недолго и называется еще метакинезом, представляя самый момент деления ядра. Каждая хромосома разделяется на две равные половинки, образовавшиеся еще раньше (в стадии клубка) расщеплением хроматинных отрезков (табл., фиг. 4 — хр), причем одна половинка каждой хромосомы направляется к одному полюсу или центрозоме, а другая — к противоположной (табл., фиг. 9). Расхождением половинок хромосом начинается анафаза, и, рассматривая деление на живых клетках, получается впечатление, что ахроматинные нити атракционного веретена или конусов (табл., фиг. 9 — ат. к.), прикрепленные к вершинам петлеобразных хромосом (хр), как бы сокращаются и тянут за собой по половинке хромосом к каждому полюсу (ц). Разделение хромосом начинается с вершин петель и постепенно переходит на их бока до конца петель, причем половинки петель, расходясь, описывают полуокружность, обращаясь своими вершинами к обоим полюсам. Направляясь к центрозомам (ц), хромосомы (хр) не доходят до них и, группируясь вокруг них (табл., фиг. 10), образуют стадию дочерних звезд (dyaster), которые претерпевают регрессивный метаморфоз (табл., фиг. 11, 12 и 13), т. е. те же изменения, которые во время профазы претерпевало материнское ядро, только в обратном порядке, проходя стадии рыхлого (табл., фиг. 13), а затем плотного клубка (dispirem), пока не приобретут вида двух нормальных, покоящихся ядер. Во время этого процесса (табл., фиг. 12—13) ахроматинные веретена (центральное и атракционное) исчезают (во время стадии dispirem), равно как и лучистость (табл., фиг. 11); зато появляются вновь ядрышки (табл., фиг. 12, 13 — яд) и оболочка ядра (табл., фиг. 12—13 — об), а центрозома, уменьшаясь в объеме, занимает первоначальное положение (табл., фиг. 11, 12, 13 — ц). Вместе с делением ядра происходит и разделение цитоплазмы на две части, а именно во время анафазы, т. е. при приближении дочерних хромосом к обеим центрозомам в клетке появляется небольшая кольцевая бороздка (в плоскости, перпендикулярной к оси веретен), которая, постепенно углубляясь, перетягивает к концу процесса деления цитоплазму на две половины.Значения этого чрезвычайно сложного и запутанного процесса деления, наблюдаемого у громадного большинства клеток растений и животных, а также и у простейших — следующее. Прежде всего, различные химические вещества (хроматин или нуклеин зерен, ахроматин или линин ячеек, парануклеин ядрышек, амфипиренин оболочки ядра), находящиеся в покоящемся ядре, с одной стороны резче отделяются друг от друга, а с другой, испытывая смещения, вступают по растворении оболочки ядра в более близкие взаимные соотношения с цитоплазмой. Затем, благодаря разным приспособлениям (центрозомы, веретена, расщепление хромосом надвое), достигается точное деление содержимого ядра на две равные части не только в количественном, но и в качественном отношении.Другой тип деления клеток и ядер — прямой или амитотический, значительно проще и встречается гораздо реже предыдущего. Во время деления цитоплазма и ядро вытягиваются в длину, вследствие появляющейся небольшой перетяжки принимают бисквитообразную форму и затем делятся надвое. Предполагают, что этот способ деления — наиболее примитивный и исчезающий теперь, чтобы дать место более совершенному делению — кариокинезу; тогда как другие склонны думать, что амитоз филогенетически более молодой процесс, который наблюдается только в вымирающих клетках или в клетках, дошедших почти до предела своей способности размножения.Литература. M. Schleiden, "Beiträge zur Phytogenesis" ("Müller's Archiv", 1838); Th. Schwann, "Mikroskopische Untersuchungen über die Uebereinstimmung in der Structur und dem Wachsthum der Thiere und Pflanzen" (1839); H. v. Mohl, "Ueber die Vermehrung der Pflanzenzellen durch Theilung" (Тюбинген, 1835); Max Schultze, "Das Protoplasma der Rhizopoden und der Pflanzenzelle" (Лейпциг, 1863); A. v. Kölliker, "Die Lehre von der thierischen Zelle" ("Zeitschr. f. wissensch. Botan.", 1845); F. Dujardin, "Recherches sur les organismes inférieurs" ("Annal. d. Sc. Natur. Zool.", 2 сер., т. IV, 1835); W. Kühne, "Untersuchungen über das Protoplasma und die Contractilität" (1864); F. Heitzmann, "Untersuchungen über das Protoplasma" ("Sitzungsber. d. K. Akad. d. Wissensch.", "Wien. Math.-Naturw. Cl.", т. LVII, 1873); F. Leydig, "Zelle und Gewebe" (Бонн, 1885); С. v. Kupffer, "Ueber Differenzirung des Protoplasma an den Zellen thierischer Gewebe" ("Schrift. d. Naturw. Ver. f. Schleswig-Holstein", т. I, 1875); E. Strasburger, "Zellbildung u. Zelltheilung" (Йена, 1876); его же, "Studien über das Protoplasma" ("Jen. Zeitschr.", т. X, 1876); W. Flemming, "Zellsubstanz, Kern und Zelltheilung" (Лейпциг, 1882); R. Altmann, "Die Elementarorganismen und ihre Beziehungen zu den Zellen" (Лейпциг, 1890, 2 изд., 1894); его же, "Die Granulalehre u. ihre Kritik" ("Arch. f. Anat. u. Physiol." Anat. Abth., 1893); O. Bütschli, "Untersuchungen über mikroskopische Schäume und das Protoplasma" (Лейпциг, 1892); G. Berthold, "Studien über Protoplasmamechanik" (Лейпциг, 1886); F. Schwarz, "Die morphologische und chemische Zusammensetzung des Protoplasmas" ("Beitr. z. Biol. d. Pflanz.", Breslau, т. V, 1887); M. Verworn, "Psycho-physiologische Protisten-Studien" (Йена, 1887); B. v. Erlanger, "Beiträge zur Kentniss d. Structur des Protoplasmas, der karyokinetischen Spindel und des Centrosoms" ("Arch. f. mikrosk. Anat.", т. XLIX, 1897). Учебники (в них обширный литературный список): О. Hertwig, "Die Zelle und die Gewebe" (Йена, 1893; есть русский перевод); Y. Delage, "La structure du Protoplasma et les théories sur l'héredité et les grands problèmes de la biologie générale" (Париж, 1895); F. Henneguy, "Leçons sur la cellule" (Париж, 1896); E. Wilson, "The cell in development and in heritance" (Лондон, 1896).В. Шевяков.
Фиг. 5. Оптический разрез через краевую часть искусственной пенистой капли (из оливкового масла и поваренной соли). alv — альвеолярный слой. Увеличение 1250.Все это Бючли справедливо считает окончательным доказательством в пользу своей теории ячеистого строения П. Примешанные к описанным комочкам, при приготовлении их, т. е. во время растирания в ступке, мельчайшие твердые частички (как, например, сажа), располагаются вдоль граней и, главным образом, в узловых точках системы ячеек пенистых капель, как и настоящие микрозомы живой П. Под микроскопом в таких искусственных пенистых каплях, особенно содержащих крупинки сажи, можно заметить восходящие и нисходящие внутренние токи, совершенно подобные тем, какие наблюдаются в живой П. некоторых одноклеточных организмов и растительных клетках, и о которых мы будем говорить ниже при разборе физиологических свойств протоплазмы. Мало того, если эти внутренние токи происходят более или менее продолжительно в одном направлении и достаточно интенсивны, то результатом их является передвижение самих искусственных пенистых капель, которые могут двигаться самопроизвольно, подобно типичным живым амебам (при помощи так называемых псевдоподий), по разным направлениям (постоянно изменяя его), более или менее продолжительное время (до 7 суток). Относительно гипотетичной причины этого движении будет сказано ниже (см. физиологические свойства П.); заметим здесь только, что аналогичные движения одноклеточных организмов (а на них сводятся вообще все движения, обнаруживаемые также и высшими животными) допускают подобное же объяснение, если ввести некоторые поправки, принимая во внимание химическую сторону явлений.Помимо изложенных теорий, построенных с помощью и на основании непосредственных микроскопических наблюдений, существуют еще теории, построенные умозрительным путем и касающиеся причины и сущности жизни, появления живого вещества, наследственности и тому подобных общих вопросов биологии — это так называемые философские теории биологии, которых мы здесь касаться не будем (см. соответственные статьи).Химические свойства П. При настоящем положении наших знаний мы даже не можем с уверенностью сказать, есть ли П. определенное химическое тело или изменяющаяся смесь различных химических тел, так как П. умирает, как только ее подвергают действию реактивов, употребляемых в аналитической химии, а, умирая, перестает быть настоящей П., теряя часть своих характерных свойств. Дальнейшее затруднение представляет еще то обстоятельство, что при анализе П. подчас невозможно отделить различные посторонние тела и включения П., представляющие продукты обмена веществ. Известно только, что в состав П. входят: углерод, водород, азот, кислород, сера, фосфор, а также фтор, хлор, кремний, натрий, калий, кальций, магний и железо. Первые из этих элементов (т. е. С, H, N, О, S и Р) образуют очень сложные и малоизученные соединения — так называемые белковые или протеиновые тела, представляющие при значительном процентном (60 до 9%) содержании воды главную составную часть П., тогда как последние входят в состав различных солей, встречающихся в П. Возникшее в последнее время с такой силой изучение морфологических свойств, т. е. тончайшего строения П., а также и изменений, происходящих в делящемся ядре и окружающей П., привели к более внимательному и точному изучению химических свойств вновь открытых морфологических частей, составляющих П. Так, в настоящее время, мы различаем очень много веществ, входящих в состав П., из коих некоторые, как, например, линин, пиренин, парануклеин и др., нам совершенно неизвестны с химической стороны: мы не знаем ни их химической формулы, ни химического состава, ни даже представляют ли они простые соединения или смесь разных соединений — нам известно только их отношение к различным красящим веществам, а также их некоторые оптические и физические свойства. Зато другие соединения удалось изолировать и изучить отчасти их химическое строение, а отчасти только их отношение к различным химическим реактивам. В общем же, белковые вещества (по Шюценбергеру формула альбумина C60H100N16O20) характеризуются очень большим молекулярным весом и очень сложным строением частицы. В состав цитоплазмы входят: 1) пластин, составляющий самую существенную часть ее и который не удалось еще изолировать; он нерастворим в поваренной соли, сернокислом магнии и главное — в пепсине и трипсине, растворим в слабой уксусной кислоте, не окрашивается основными анилиновыми красками, а только кислыми. 2) Нуклео-альбумины — белковые, содержащие фосфор вещества, растворимые в желудочном соке, разлагающем их на пептоны, остающиеся в растворе, и выпадающую нуклеинную кислоту. 3) Глобулины — белковые тела, не содержащие фосфора, нерастворимые в воде, но растворимые в разбавленных растворах (5—10%) солей. 4) Лецитины (очень сложная химическая формула) — представляют сочетание холина (C5H15NO2) с жирами, заключающими фосфорную кислоту, растворимы в алкоголе. 5) Холестерин (C26H43OH) — непредельный одноатомный алкоголь, растворимый в эфире и кипящем спирте. 6) Фосфорнокислые и хлористые соли калия, натрия, магния и кальция, а также железо в соединении с нуклео-альбуминами. Химический состав в тонких (клеточных) оболочках неизвестен, тогда как в более толстых главными составными частями являются: хитин (С15H26N2O10), кератин, клетчатка или целлюлоза (C6H10O5) и др. В состав ядра входят: хроматин, состоящий из холестерина и нуклеина, богатого фосфором и нерастворимого в желудочном соке вещества; пиренин и линин или парахроматин, по-видимому, представляют соединения нуклео-альбуминов с пластином. Из общих химических свойств живой П. следует упомянуть ее щелочную реакцию (причем сок некоторых растительных клеток может быть кислым) и способность прижизненной окраски, т. е. окрашиваться слабыми растворами нейтральных анилиновых красок.Физиологические свойства П. Самыми характерными свойствами живой П. являются способность к движению и раздражимость, которые, в свою очередь, представляют лишь приготовительные условия для питания и проистекающих из последнего явлений роста и размножения. Движение, обнаруживаемое П., бывает двоякое. 1) Движение внутри П., выражающееся в перемещении зернышек и различных других включений — это так называемые внутренние токи, и 2) движение, выражающееся в изменении наружной формы и в перемене места самой П. — это так называемые амёбоидные и локомоторные движения. Первый род движения, без сомнения, свойственен всякой живой П., но в большинстве случаев движение это происходит настолько медленно, что ускользает от нашего непосредственного наблюдения. Всего энергичнее оно происходит в клетках растений и в некоторых простейших, но было наблюдаемо также и в животных клетках. Сущность этого движения заключается в том, что от любой точки поверхности комка П. или клетки отходит ток, идущий внутрь и направляющийся по оси (совпадающей, большей частью, с продольной осью комка П. или клетки) комка П.; достигнув противоположной точки поверхности, восходящий осевой ток переходит в нисходящие токи, идущие параллельно поверхности комка и собирающиеся в точке отправления восходящего тока. Кроме того, наблюдается в некоторых случаях еще движение в стенкоположной (т. е. наружной) П. клеток, и всегда в одном направлении, так называемое вращательное движение или rotatio (см. Клеточка). Амёбоидное движение, заключающееся в образовании в любом или определенном месте поверхности П. небольших пальчатых или нитевидных псевдоподий или ложноножек, быстро выпячиваемых и втягиваемых обратно в тело, есть следствие внутренних токов, продолжающихся более или менее долго и с достаточной силой в одном направлении, и если наружный слой П. не настолько уплотнен, чтобы мог препятствовать образованию таких отростков. Результатом образования таких псевдоподий — является поступательное движение или перемещение П. организма (простейшие) или клетки (яйца многоклеточных животных, лимфатические тельца, белые кровяные шарики позвоночных и т. п.). В простейшем случае псевдоподии представляются в виде тупых, простых или разветвленных отростков и, по мере уплотнения наружного слоя (см. Простейшие) становятся тонкими, нитевидными (образующими подчас сетчатые сплетения), на морфологической подкладке которых развиваются при дальнейшем уплотнении П. жгутики и реснички, т. е. особые органы передвижения одноклеточных организмов, встречающиеся также и у клеток некоторых тканей многоклеточных животных (наружные мерцательные покровы низших червей и разных личинок, мерцающий эпителий кишечника и др.). В этих псевдоподиях, при сильных увеличениях, заметно движение заключающихся в них зернышек, которое даже в тончайших нитях происходит в двух противоположных направлениях одновременно. Причина разнообразных движений П. остается пока невыясненной; наиболее заслуживающей внимания является гипотеза Бючли. Приготовленные им искусственным путем (см. выше) пенистые капли обнаруживают не только характерные внутренние токи, но и перемещаются более или менее быстро в разных направлениях при помощи псевдоподий наподобие настоящих амёб. При нагревании эти движения становятся быстрее, а при пропускании гальванического (индукционного) тока пенистые капли направляются к катоду. Это движение Бючли объясняет следующим образом. Представим себе, что в любом месте поверхности пенистой капли разорвутся или лопнут по какой-либо причине несколько ячеек, вследствие чего мыльный раствор вытечет из них наружу и распространится в этой точке по поверхности. Но поверхностное натяжение этой жидкости в воде меньше поверхностного натяжения масла (образующего стенки ячеек), поэтому в данном месте образуется небольшое возвышение, причем поверхностные ячейки подвигаются вперед и освобождают позади себя место, которое заполняется надвигающимися позади лежащими ячейками и так далее. На поверхности разрываются новые ячейки, и явление продолжается, вызывая полное перемещение ячеек и передвижение всей пенистой массы. Бючли полагает, что и тожественные явления движения амёбы должны быть сведены к той же причине, т. е. чисто механической. В П. наблюдаются еще особые и своеобразные движения во время размножения, направленные к разделению и новой группировке различных существенных элементов, входящих в состав П., т. е. ее цитоплазмы и, главное, — ядра. Наконец, различают еще перемещения П. и отдельных организмов в определенном направлении, обуславливаемые различными физическими, химическими или физиологическими агентами, как то: тяжестью, светом, теплотой, кислородом, углекислотой, питательными веществами и т. п.; это так называемый геотаксис, фототаксис, термотаксис, хемотаксис и т. д. (см. Тропизм и Таксис), направляющие движение или к источнику раздражения (положительный таксис), или в противоположную сторону (отрицательный).Раздражимость. — Особое свойство, присущее лишь живой П., отвечать тем или иным способом на самые разнообразные влияния окружающей среды. Эта раздражимость проявляется специфическими реакциями в зависимости от самой природы П., а способ, которым последняя реагирует, называется эффектом раздражения. Одно и то же раздражение может вызвать разные эффекты в различных П. или организмах, так, например, свет притягивает одни и отталкивает другие организмы. Весьма различные раздражители могут также вызывать в той же П. одинаковый эффект раздражения, что называется специфической энергией; так, например, мускульная клетка на всякое раздражение отвечает сокращением. Эти явления зависят от особого и разнообразного строения самой П. Таким образом, хотя раздражимость и есть основное свойство живой П., но проявляется она сообразно с особенностями строения протоплазмы. Все раздражители могут быть сведены к трем категориям. 1) Механические, как то: прикосновение, сотрясение, сжатие, удар и колебания; 2) физические, как то: тяжесть, свет, теплота и электричество; 3) химические, т. е. всякие химические вещества. Механические раздражения заставляют П. сокращаться, втягивать псевдоподии, останавливаться и принимать шаровидную форму (например, амёбу), причем слабые раздражения действуют только на непосредственно затронутое место, тогда как более сильные распространяются на большие расстояния. Световые раздражения вызывают характерные изменения формы в П., а также и движения в определенном направлении у свободно подвижных одноклеточных организмов. При действии света псевдоподии некоторых корненожек и миксомицет втягиваются и вытягиваются вновь в темноте, а из многих одноклеточных организмов, снабженных ресничками и жгутиками, одни собираются на обращенной к свету стороне сосуда, а другие на противоположной (положительная и отрицательная фототаксия). Не меньшее действие оказывает свет на П. хроматофоров или зерен хлорофилла растений, заставляя их вращаться или перемещаться или даже менять свою форму. Наконец, пигментные клетки и хроматофоры животных весьма сильно реагируют на свет и, изменяя свою величину и форму, обуславливают изменение окраски рыб, амфибий, головоногих моллюсков и др. Термические раздражения играют еще более заметную роль в жизни П. Для всякой П. существуют пределы температурные (вне которых наступает смерть П.), весьма разнообразные, в особенности же верхний (40°—130° Ц.). До достижения предельных температур наступает явление, называемое оцепенением; П. при известной температуре не обнаруживает более жизненных явлений (например, движения), но проявляет их, как только температура изменится. Термические раздражения резче всего вызываются в возрастании скорости внутренних токов или перемещений по мере возвышения температуры; причем — максимум скорости составляет оптимум температуры, лежащий несколькими градусами ниже предельной температуры, при которой наступает оцепенение от тепла. Электрические раздражения проявляются при действии гальванических токов, индукционных и постоянных, на П. лишь в том случае, если непосредственно проходят по ней. При действии слабых индукционных ударов движение зернышек (внутренние токи) в П. останавливается, а амёбы и белые кровяные шарики на короткое время прекращают свое движение. От сильных ударов псевдоподии втягиваются, движения прекращаются, организмы съеживаются в шар, лопаются и разрушаются. Постоянный ток (слабый), пропускаемый через одноклеточные организмы, побуждает их к движению в определенном направлении (гальванотропизм), причем некоторые во время замыкания тока покидают анод и направляются к катоду, где и скопляются, а по размыкании тока направляются к аноду, тогда как другие, обратно, при замыкании тока направляются к аноду, а при размыкании — к катоду (отрицательный и положительный гальванотропизм). Наконец, химические раздражители весьма разнообразны как по своему составу, так и действию, оказывая или всестороннее влияние на П. (анестезирующие вещества), или в одном определенном направлении (см. Хемотаксис и Хемотропизм). В общем же следует заметить, что П. может до известной степени приспособляться к химическим изменениям окружающей среды (соленость воды, растворы, присутствие и обилие кислорода, углекислоты, сероводорода и т. д.), если только эти изменения совершаются постепенно, причем П. претерпевает некоторые изменения в своем строении и жизненной деятельности.Формирующая деятельность П. является следствием раздражимости и способности к движению. В этой деятельности следует различать три стадии: поглощение, ассимиляцию или превращение и выделение. Посторонние вещества могут быть поглощаемы П. извне в газообразном, жидком и твердом состояниях: в первых двух — всякой П., а в последнем — лишь П. одноклеточных организмов и некоторых клеток многоклеточных животных. Из газообразных веществ наиболее важны кислород и углекислота. Поглощение кислорода, называемое дыханием, имеет особо важное значение для процессов обмена веществ, так как благодаря ему в П. образуются соединения более богатые кислородом, дальнейшим расщеплением и окислением которых получаются вода и углекислота. При этих химических реакциях, составляющих "регрессивный метаморфоз" или разрушение органического вещества, развивается теплота, являющаяся источником живых сил. Поглощение углекислоты, присущее только растительному царству и некоторым простейшим (см.), ведет к так называемому прогрессивному метаморфозу, т. е. к синтезу органических соединений с высокой частицей. При этом процессе восстановления углекислоты потребляется теплота, которая связывается во вновь образованном веществе в виде потенциальной энергии. Поглощение П. жидких веществ представляет главный источник питания как для растений, так и для животных. Первые заимствуют питательные вещества в растворенном виде или из воды, или же из влажной почвы при помощи корней, тогда как клетки животных получают их из жидкостей (пищевая кашица, хилус, лимфа, кровь), приготовляемых сложным путем в теле самих животных. При таком поглощении П. различных клеток оказывает предпочтение тому или другому веществу, что и называется избирательной способностью, причина которой еще не выяснена, но, по всей вероятности, сводится к химическому сродству разных веществ, встречающихся в П. постоянно или временно возникающих при обмене веществ. Поглощение твердых тел П. присуще лишь некоторым одноклеточным организмам (см. Простейшие, питание) и некоторым тканевым клеткам многоклеточных животных (см. Фагоцитоз). Вещества, принятые П., перерабатываются в П. и превращаются в многочисленные и самые разнообразные вещества. Химическая сторона этих превращений остается еще большей частью совершенно темной. Известно, например, только, что встречающийся в растениях сахар и крахмал образуются синтетическим путем, но едва ли прямым синтезом углерода и воды, а жирные масла — превращением углеводов, возникших через посредство хлорофилла и т. д. Далее, что превращение углеводов, жиров и белков из пассивного состояния в активное происходит при помощи так называемых ферментов (см.): так, диастаза превращает крахмал в сахар, инвертин расщепляет тростниковый сахар на декстрозу и левулозу, птиалин (фермент слюны) превращает крахмал в декстрин и виноградный сахар и т. д., что белки для дальнейшего потребления пептонизируются, а жиры расщепляются на глицерин и жирные кислоты и т. д. (см. Питание растений и Пищеварение). Не более выяснена роль П. при превращении веществ: так, одни предполагают, что различные вещества получаются превращением самой П. путем химических превращений и расщеплений ее частиц, а другие, что они образуются из пластических веществ, углеводов, жиров и пр., воспринятых П. при обмене веществ. Лучше известна морфологическая сторона превращений, т. е. результаты или продукты обмена веществ, которые отлагаются внутри П. или на ее поверхности в бесформенном виде или в виде так называемых включений, отличимых оптически. Сюда относятся вакуоли, жир, гликоген, муциген, крахмал, пигментные зернышки, кристаллы гуанина и разных солей, скелеты, разные оболочки и т. п. Наконец, выделение или удаление из П. негодных продуктов обмена веществ завершает процесс формирующей деятельности П. Эти непригодные П. или организму вещества выводятся наружу также в трояком состоянии, и в твердом лишь исключительно некоторыми простейшими и клетками Metazoa. В газообразном виде выводится углекислота (при дыхательных процессах), кислород (растениями) и водяные пары, а также и другие различные газы, а в жидком состоянии — моча, мочевая кислота, гиппуровая кислота, гуанин и т. п. Главным и непосредственным результатом формирующей деятельности П. является образование новой живой П., которая, по мнению Бючли, образуется в виде маленьких ячеек, возникающих между прежними ячейками, т. е. в их узловых точках, и постепенно увеличивающихся в объеме. Конечный результат таких новообразований выражается в росте самой П.Размножение. Рост П., рассматриваемой приуроченной к клетке, не может, однако, продолжаться беспредельно. Всякая клетка имеет свою предельную величину, по достижении которой наступает размножение, которое поэтому может быть названо ростом за пределы индивидуальности. Каждая новая клетка образуется делением старой, точно так же, как каждое ядро — делением другого ядра. Деление может быть прямое или амитотическое и непрямое или кариокинетическое или митотическое. При непрямом [Хотя сущность кариокинеза изложена уже в ст. Клеточка, но за последние три года было сделано столько крупных открытий, существенно изменяющих и дополняющих общую картину этого важного для биологи процесса, что мы излагаем его вкратце вновь, согласно новейшему учению.] делении, называемом также кариокинезом (κάρυον — ядро, κίνησις — движение) или митозом (μίτος — нить) происходят крупные изменения, как в цитоплазме, так и в ядре, появляющиеся одновременно. Мы различаем, собственно, три периода: профазу — изменения в материнском ядре, метафазу — само деление и анафазу или изменения, происходящие в дочерних ядрах. Изменения, происходящие во время профазы в ядре, выражаются в том, что хроматинные зернышки, залегающие в стадии покоя ядра в узловых точках ахроматичных стенок ячеек, начинают передвигаться (табл., фиг. 1) и располагаются порядно, образуя в совокупности волнистые нити, между которыми заметны еще ахроматинные стенки ячеек (табл., фиг. 2) — стадии плотного клубка (spirem). Различают или одну сплошную хроматинную нить (табл., фиг. 3) в ядре, или несколько отрезков. Затем хроматинные нити начинают укорачиваться, причем они увеличиваются в толщине, их зазубренные края сглаживаются (табл., фиг. 3), и если была одна сложная хроматинная нить, то она вскоре распадается на несколько (всегда определенное и большей частью четное число) отрезков — стадия рыхлого клубка. Эти отрезки, называемые хромосомами (χρώμα — цвет, σώμα — тело), имеют весьма разнообразную форму (нитей, крючков, палочек и шариков) и вскоре расщепляются в продольном направлении надвое (табл., фиг. 4 — хр), причем обе половины остаются вместе. К концу этого процесса исчезает находящееся в ядре ядрышко (табл., фиг. 1—3 — яд); по мнению других, оно у некоторых клеток дает центрозому, а у большинства (за исключением всех простейших) клеток и оболочку ядра (табл., фиг. 1—4 — об). В это же время происходят изменения и в окружающей ядро цитоплазме, а именно от центрозомы (табл., фиг. 5 — ц), окруженной архоплазмой или атракционной сферой (арх) и залегающей в состоянии покоя вблизи ядра или в небольшом углублении последнего, начинают отходить по всем направлениям тонкие плазматические лучи, обуславливающие впоследствии лучистость цитоплазмы (табл., фиг. 5 — л). Центрозома вытягивается в длину, принимает бисквитообразную форму и делится на две (табл., фиг. 6 — ц), из коих каждая окружена архоплазмой. По мере расхождения обеих центрозом, между ними выступает зачаток веретена, разрастающегося впоследствии (при дальнейшем расхождении центрозом), в так называемое центральное веретено (табл., фиг. 7 — ц. в), состоящее из тонких ахроматических нитей, идущих от одной центрозомы (ц) к другой и образующих в совокупности два конуса, соединенных своими основаниями. Во время расхождения центрозом начинает растворяться оболочка ядра (табл., фиг. 4 — об) и исчезает сначала в месте ближайшем к центральному веретену (табл., фиг. 4 — п. и фиг. 7), а затем и в других местах. Центрозомы, расходясь, т. е. удаляясь друг от друга, вытягивают в длину центральное веретено и располагаются, наконец, на двух противоположных точках ядра (табл., фиг. 8 — ц). В продолжение всего описанного процесса плазматические нити или лучи, отходящие от центрозом, группируются около каждой центрозомы в виде двух расходящихся снопов лучей или конусов (табл., фиг. 7), так называемых антиподных (ан. к.) и атракционных (ат. к.) конусов. Первые впоследствии (при окончательном расхождении центрозом) направляются в сторону, диаметрально противоположную от направления ахроматинного центрального веретена (табл., фиг. 8 — ан. к.), а последние (табл., фиг. 8 — ат. к.) направляются к хромосомам (хр) ядра таким образом, что к каждой хромосоме от каждой центрозомы направляются по одной плазматической нити. Эти нити, совместно с образовавшимся расхождением центрозом центральным веретеном (ц. в.), оказывают направляющее действие на хромосомы, которые к концу этой стадии (табл., фиг. 8) располагаются в одной плоскости, перпендикулярной к оси ахроматинного центрального веретена (полюса которого составляют центрозомы), снаружи или вокруг последнего. Они перегибаются при этом наподобие петель (табл., фиг. 7 и 8 — хр) или крючков, причем вершины этих петель обращены к центру ядра, и к ним с двух сторон подходят тонкие нити, идущие от центрозом и образующие в общей сложности два конуса — атракционных конуса (табл., рис. 8 — ат. к.), соединенных своими основаниями. Эти два конуса, образуют второе ахроматинное веретено, как бы одевающее центральное, и которое в отличие от последнего называется направляющим или атракционным веретеном. Этим процессом заканчивается профаза деления, и получившаяся стадия ядра называется звездой (aster); наступающая затем метафаза продолжается очень недолго и называется еще метакинезом, представляя самый момент деления ядра. Каждая хромосома разделяется на две равные половинки, образовавшиеся еще раньше (в стадии клубка) расщеплением хроматинных отрезков (табл., фиг. 4 — хр), причем одна половинка каждой хромосомы направляется к одному полюсу или центрозоме, а другая — к противоположной (табл., фиг. 9). Расхождением половинок хромосом начинается анафаза, и, рассматривая деление на живых клетках, получается впечатление, что ахроматинные нити атракционного веретена или конусов (табл., фиг. 9 — ат. к.), прикрепленные к вершинам петлеобразных хромосом (хр), как бы сокращаются и тянут за собой по половинке хромосом к каждому полюсу (ц). Разделение хромосом начинается с вершин петель и постепенно переходит на их бока до конца петель, причем половинки петель, расходясь, описывают полуокружность, обращаясь своими вершинами к обоим полюсам. Направляясь к центрозомам (ц), хромосомы (хр) не доходят до них и, группируясь вокруг них (табл., фиг. 10), образуют стадию дочерних звезд (dyaster), которые претерпевают регрессивный метаморфоз (табл., фиг. 11, 12 и 13), т. е. те же изменения, которые во время профазы претерпевало материнское ядро, только в обратном порядке, проходя стадии рыхлого (табл., фиг. 13), а затем плотного клубка (dispirem), пока не приобретут вида двух нормальных, покоящихся ядер. Во время этого процесса (табл., фиг. 12—13) ахроматинные веретена (центральное и атракционное) исчезают (во время стадии dispirem), равно как и лучистость (табл., фиг. 11); зато появляются вновь ядрышки (табл., фиг. 12, 13 — яд) и оболочка ядра (табл., фиг. 12—13 — об), а центрозома, уменьшаясь в объеме, занимает первоначальное положение (табл., фиг. 11, 12, 13 — ц). Вместе с делением ядра происходит и разделение цитоплазмы на две части, а именно во время анафазы, т. е. при приближении дочерних хромосом к обеим центрозомам в клетке появляется небольшая кольцевая бороздка (в плоскости, перпендикулярной к оси веретен), которая, постепенно углубляясь, перетягивает к концу процесса деления цитоплазму на две половины.Значения этого чрезвычайно сложного и запутанного процесса деления, наблюдаемого у громадного большинства клеток растений и животных, а также и у простейших — следующее. Прежде всего, различные химические вещества (хроматин или нуклеин зерен, ахроматин или линин ячеек, парануклеин ядрышек, амфипиренин оболочки ядра), находящиеся в покоящемся ядре, с одной стороны резче отделяются друг от друга, а с другой, испытывая смещения, вступают по растворении оболочки ядра в более близкие взаимные соотношения с цитоплазмой. Затем, благодаря разным приспособлениям (центрозомы, веретена, расщепление хромосом надвое), достигается точное деление содержимого ядра на две равные части не только в количественном, но и в качественном отношении.Другой тип деления клеток и ядер — прямой или амитотический, значительно проще и встречается гораздо реже предыдущего. Во время деления цитоплазма и ядро вытягиваются в длину, вследствие появляющейся небольшой перетяжки принимают бисквитообразную форму и затем делятся надвое. Предполагают, что этот способ деления — наиболее примитивный и исчезающий теперь, чтобы дать место более совершенному делению — кариокинезу; тогда как другие склонны думать, что амитоз филогенетически более молодой процесс, который наблюдается только в вымирающих клетках или в клетках, дошедших почти до предела своей способности размножения.Литература. M. Schleiden, "Beiträge zur Phytogenesis" ("Müller's Archiv", 1838); Th. Schwann, "Mikroskopische Untersuchungen über die Uebereinstimmung in der Structur und dem Wachsthum der Thiere und Pflanzen" (1839); H. v. Mohl, "Ueber die Vermehrung der Pflanzenzellen durch Theilung" (Тюбинген, 1835); Max Schultze, "Das Protoplasma der Rhizopoden und der Pflanzenzelle" (Лейпциг, 1863); A. v. Kölliker, "Die Lehre von der thierischen Zelle" ("Zeitschr. f. wissensch. Botan.", 1845); F. Dujardin, "Recherches sur les organismes inférieurs" ("Annal. d. Sc. Natur. Zool.", 2 сер., т. IV, 1835); W. Kühne, "Untersuchungen über das Protoplasma und die Contractilität" (1864); F. Heitzmann, "Untersuchungen über das Protoplasma" ("Sitzungsber. d. K. Akad. d. Wissensch.", "Wien. Math.-Naturw. Cl.", т. LVII, 1873); F. Leydig, "Zelle und Gewebe" (Бонн, 1885); С. v. Kupffer, "Ueber Differenzirung des Protoplasma an den Zellen thierischer Gewebe" ("Schrift. d. Naturw. Ver. f. Schleswig-Holstein", т. I, 1875); E. Strasburger, "Zellbildung u. Zelltheilung" (Йена, 1876); его же, "Studien über das Protoplasma" ("Jen. Zeitschr.", т. X, 1876); W. Flemming, "Zellsubstanz, Kern und Zelltheilung" (Лейпциг, 1882); R. Altmann, "Die Elementarorganismen und ihre Beziehungen zu den Zellen" (Лейпциг, 1890, 2 изд., 1894); его же, "Die Granulalehre u. ihre Kritik" ("Arch. f. Anat. u. Physiol." Anat. Abth., 1893); O. Bütschli, "Untersuchungen über mikroskopische Schäume und das Protoplasma" (Лейпциг, 1892); G. Berthold, "Studien über Protoplasmamechanik" (Лейпциг, 1886); F. Schwarz, "Die morphologische und chemische Zusammensetzung des Protoplasmas" ("Beitr. z. Biol. d. Pflanz.", Breslau, т. V, 1887); M. Verworn, "Psycho-physiologische Protisten-Studien" (Йена, 1887); B. v. Erlanger, "Beiträge zur Kentniss d. Structur des Protoplasmas, der karyokinetischen Spindel und des Centrosoms" ("Arch. f. mikrosk. Anat.", т. XLIX, 1897). Учебники (в них обширный литературный список): О. Hertwig, "Die Zelle und die Gewebe" (Йена, 1893; есть русский перевод); Y. Delage, "La structure du Protoplasma et les théories sur l'héredité et les grands problèmes de la biologie générale" (Париж, 1895); F. Henneguy, "Leçons sur la cellule" (Париж, 1896); E. Wilson, "The cell in development and in heritance" (Лондон, 1896).В. Шевяков.
Энциклопедический словарь Ф.А. Брокгауза и И.А. Ефрона. — С.-Пб.: Брокгауз-Ефрон. 1890—1907.