- Усачи (жуки)
-
Усачи 
Научная классификация промежуточные рангиДомен: Эукариоты Царство: Животные Подцарство: Эуметазои Без ранга: Двусторонне-симметричные Без ранга: Первичноротые Без ранга: Линяющие Без ранга: Panarthropoda Тип: Членистоногие Подтип: Трахейнодышащие Надкласс: Шестиногие Класс: Насекомые Подкласс: Крылатые насекомые Инфракласс: Новокрылые насекомые Надотряд: Насекомые с полным превращением Отряд: Жесткокрылые Подотряд: Разноядные жуки Инфраотряд: Кукуйиформные Надсемейство: Хризомелоидные Семейство: Усачи Международное научное название Типовой род CerambyxПодсемейства: Anoplodermatinae
Спондилидины (Spondylidinae)
Dorcasominae
Настоящие усачи (Cerambycinae)
Disteniinae
Ламиины (Lamiinae)
Усачики (Lepturinae)
Necydalinae
Парандрины (Parandrinae)
Philinae
Усачи-прионины (Prioninae)
Систематика
на Викивидах
Изображения
на ВикискладеITIS 114497 NCBI 34667 EOL 357 Усачи́, или дровосе́ки (лат. Cerambycidae) — разветвлённое и многочисленное (пятое по количеству видов) семейство жуков (Coleoptera). Одним из наиболее характерных и отличительных, хотя и условных, признаков представителей семейства являются длинные сегментированные усы, часто значительно превышающие длину тела, иногда в 2—3, и даже в 4—5 раз[1].
Содержание
Общая характеристика
По приблизительным оценкам, общая численность семейства составляет около 26 000 видов[2], тем не менее, это не окончательная цифра, поскольку ежегодно открывают и описывают новые виды из тропических и экваториальных районов, реже — из Европы, Малой Азии и Северной Америки. На территории России обитает 583 вида[3][4], для сравнения: в бывшем СССР — 880, в США — 1200, в Канаде — 350, а на Украине около 280 видов[5]. В то же время, усачи являются одним из наиболее изученных семейств жесткокрылых.
Жуки-дровосеки широко распространены по всему земному шару, в шести зоогеографических областях, и их распространение тесно взаимосвязано с кормовыми растениями, преимущественно с древесными породами[1].

Большинство усачей — жуки средней величины, но есть среди них и гиганты, относящиеся к крупнейшим жукам в мире. Самым крупным представителем семейства является встречающийся в Южной Америке дровосек-титан (Titanus giganteus), достигающий в длину 167 мм[6], а по некоторым неподтверждённым источникам даже до 210 мм[7]. Вторым по величине является также южноамериканский вид — крупнозуб оленерогий (Macrodontia cervicornis), достигающий длины 16,5 см[8]. Усач Xixuthrus heros с Фиджи также входит в тройку крупнейших жуков семейства, достигая длины 15 см[9][10]. Самым крупным европейским видом является усач-плотник (Ergates faber) длиной до 6 см[11]. К усачам относится и самый большой жук России — уссурийский реликтовый усач (Callipogon relictus), длиной до 11 см[12].
В большинстве случаев самцы отличаются от самок более длинными усиками, удлинёнными верхними челюстями, более заострённым брюшком и часто иным окрасом (половой диморфизм)[1].
Многие усачи могут издавать резкие скрипящие звуки, воспроизводимые путём трения рёбрышка на заднем крае переднегруди о шершавую поверхность среднегруди. Некоторые усачи, обитающие на Гавайских островах, способны издавать резкие звуки путём трения бёдер задних ног о надкрылья. Данные скрипящие звуки используются жуками в случае нападения хищников, и, вероятно, носят отпугивающий характер[13][14].
Непохожи на обычных усачей виды небольшого рода парандра (Parandra) — одного из наиболее примитивных в составе семейства. Плоское и широкое тело этих жуков с очень крупной головой и мощными челюстями делает их похожими на рогачей[13].
Представители семейства весьма разнообразны по форме и длине тела, окраске, скульптуре надкрылий, наличию шипов, выростов, бугров на переднеспинке и надкрыльях — всё это, делает их одним из излюбленных семейств жуков для коллекционирования энтомологами.
Морфология имаго
Морфология жука-усача на примере большого елового усача 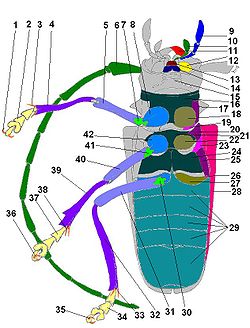
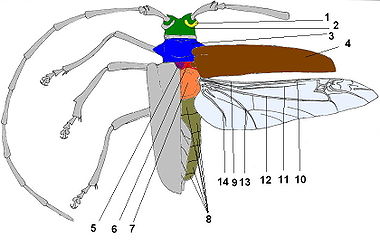
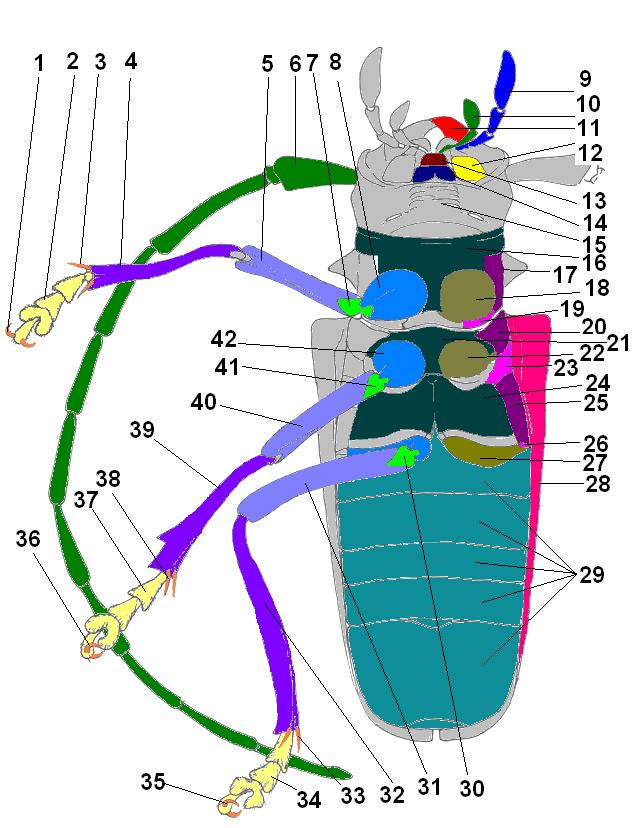
Морфология вентральной стороны жука
1, 35, 36 — коготок, 2, 34, 37 — лапка, 3, 33, 38 — шпора, 4, 32, 39 — голень, 5, 31, 40 — бедро, 6 — усик, 7, 30, 41 — вертлуг, 8, 42 — тазик, 9 — челюстное щупальце, 10 — губное щупальце, 11 — мандибула, 12 — дополнительный язычок, 13 — язычок, 14 — подбородок, 15 — горло, 16 — переднегрудь, 17, 20, 25 — эпистерны, 18, 22, 27 — тазиковые впадины, 19, 23, 26 — эпимеры, 21 — среднегрудь, 24 — заднегрудь, 28 — эпиплевры, 29 — стерниты брюшкаМорфология дорсальной стороны жука и жилкования крыла.
1 — сложный фасеточный глаз, 2 — голова, 3 — переднеспинка, 4 — надкрылье, 5 — щиток, 6 — среднеспинка, 7 — заднеспинка, 8 — тергиты брюшка, 9 — костальная жилка крыла, 10 — субкостальная жилка крыла, 11 — радиальная жилка крыла, 12 — медиальная жилка крыла, 13 — первая анальная жилка крыла, 14 — вторая анальная жилка крылаГолова
Голова обычно средних размеров, только изредка большая, благодаря сильно развитым верхним челюстям (некоторые Prioninae), округой формы, своей задней частью входит в переднегрудь. В пределах семейства, выделяют два типа головы[1]:
- первый тип — направленная вперёд или косо книзу под небольшим углом к центральной оси тела (Prioninae, Lepturinae, Cerambycinae, Aseminae, Necydalinae)
- второй тип — направленная вниз под углом 90 градусов к основной оси тела (Lamiinae).
Мандибулы хорошо развитые, у многих видов большие, особенно сильно развиты у многих Prioninae у которых более или менее резко выраженный половой диморфизм в строении верхних челюстей, у самцов они развиты значительно сильнее, чем у самок. Внутренний край мандибул гладкий, или несёт один или несколько зубцов. Форма челюстей различна — от почти прямых и коротких до длинных и сильно изогнутых (Prioninae, например, Dorysthenes paradoxus). Нижние челюсти (maxillae) состоят из двух лопастей: внутренняя лопасть короткая или отсутствует (Prioninae), у трибы Spondylini обе лопасти редуцированы и рудиментарны. Нижнечелюстные щупики состоят из четырёх члеников, иногда первый членик укорочен или редуцирован. Верхняя губа развита различно у представителей разных групп, обычно имеет форму поперечного прямоугольника или трапеции[1][14].



Derobrachus sp. Голова направлена вперёд — типично для подсемейства Prioninae Clytus arietis. Голова направлена под углом 40° к оси тела — типично для подсемейства Cerambycinae Acanthocinus aedilis. Голова направлена под углом 90° к оси тела — типично для подсемейства Lamiinae По бокам головы, охватывая основания усиков, располагаются сложные, фасеточные глаза, которые имеют более или менее выемчатую форму. Иногда, глаза целиком разделяются на две части — при этом создаётся впечатление, что у жука две пары глаз (некоторые виды Lamiinae). На верхней стороне головы различают несколько слабо разграниченных частей: шею, затылок, виски, темя, лоб и наличник. Шеей называется та часть головной капсулы, которая входит у грудь. Часть головы, примыкающая к шее, называется затылком и имеет выпуклую форму. По обе стороны от затылка, имеются характерные выступы — виски. Между глазами, выше затылка располагается темя, которое к передней части переходит в лоб[14].
 Самец и самка усача Acanthocinus aedilis. Сразу обращают на себя внимание длинные сегментированные усы самца, превышающие длину его тела в 4 раза.
Самец и самка усача Acanthocinus aedilis. Сразу обращают на себя внимание длинные сегментированные усы самца, превышающие длину его тела в 4 раза.
На лбу, между глазами, расположены усики (антенны), основания которых находятся во впадинах. Усики по большей части простые, реже пильчатые, чешуйчатые или с пластинчатыми выростами на члениках; очень редко неправильные. Состоят из 11 члеников, а у некоторых родов (Agapanthia и др.) — из 12 члеников. Иногда весь род характеризуется именно 12-члениковыми усиками (Agapanthia), иногда же, 12-члениковость свойственна только некоторым видам данного рода, наблюдается одинаково у обоих полов (Prionus insularis) или же самцы обладают 12-ти, а самки 11-члениковыми усиками (дровосек-кожевник)[1]. У представителей рода Prionus число члеников усиков может быть значительно увеличено, у некоторых видов до 25—31, членики укорочены и несут широкие и длинные пластинчатые придатки[14]. Половой диморфизм выражается в длине усиков (у самцов длиннее, чем у самок), гораздо реже в строении или количестве самих члеников[1]. Членики усиков обычно цилиндрической формы, иногда на вершине слегка утолщены или вздуты, изредка настолько сильно, что выглядят узловатыми (большинство видов Cerambyx)[1]. Первый членик, иногда (у родов Monochamus, Lamia, Morimus), имеет цикатрикс — характерный плоский выступ в форме полумесяца рядом с вершиной. Усики могут закидываться на спину и выпрямляться только вдоль верхней стороны тела, но никогда усачи не поджимают их под себя — один из признаков, отличающих семейство усачей от других семейств жуков[1].
Усики обычно длинные (за данную особенность семейство и получило своё название), обычно длиннее тела (иногда в 4-6 раз), однако имеются виды с очень короткими усиками (короткоусый сосновый усач, южноамериканский усач-землерой Hypocephalus armatus).
На передней части лба имеется шов. К наличнику прикрепляется верхняя губа, которая частично прикрывает мандибулы. На нижней части головы между шеей и ротовым аппаратом находится горло. Его передняя и самая узкая часть, прилегающая к нижней губе, называется субментумом или подбородком, а продольные бороздки возле его основы — горловыми швами[14].
Грудь
Грудь состоит из трёх сегментов, из которых наиболее развитым является первый. На дорсальной стороны они называются спинками: передне-, средне- и заднеспинка. Переднеспинка часто имеет боковой шип или шипы (дровосек-титан, Monochamus, большой дубовый усач, Rhagium и др.) или бугорок (род Carilia, Pachyta и т. п.), а у других просто округлённая (Anastrangalia, Lepturobosca, Plagionotus и др.). Сверху заметной является лишь переднеспинка, а среднеспинка, кроме щитка, и заднеспинка скрыты надкрыльями[1]. На нижней стороне грудь состоит из передне-, средне- и заднегруди. Каждый из этих сегментов, в свою очередь, состоит из центральной части — диска, отделяемых от него боковой — эпистерн, и задней — эпимер, сторон. В задней части каждого сегмента груди находятся тазиковые впадины, в которых расположены тазики — часть конечностей[1].
Надкрылья и крылья
Надкрылья обычно развиты хорошо, длинные, прикрывают нижние крылья и брюшко целиком, у самок часто из-под них выдаётся только последний сегмент брюшка[1]. Надкрылья впереди обыкновенно имеют хорошо развитые плечевые бугорки и крепятся к среднеспинке. Изредка могут быть сильно укорочены (род Necydalis, у некоторых родов трибы Molorchini, Molorchus, Leptidea, Brachypterorna). Укороченные надкрылья, то прикрывающие значительную часть брюшка, то почти совсем его не прикрывающие, наблюдаются у самок некоторых полупустынных и пустынных среднеазиатских видов Prionus и некоторых видов рода Apatophysis. Скульптура надкрылий разнообразна: от простой мелкоточечной пунктировки и сложного рисунка из морщинистых бугорков и выступов, образующих ямки и ячейки, до полностью гладкой и блестящей. Часто надкрылья несут продольные выступающие или скрытые возвышения, переходящие порой в рёбрышки[1][13]. Форма надкрылий варьирует у различных групп от узких и вытянутых (Calamobius, Theophilea) до широких и коротких (Acmaeops). У ряда видов Lepturinae надкрылья могут быть сильно суженными к концу, приобретая вид вытянутого треугольника. У видов Dorcadion они более или менее овальной формы. Вершина надкрылий или округлена, или срезана. Часть надкрылий, которая заходит на нижнюю сторону тела, называется эпиплеврами[1].
Крылья имеются у большинства родов[13]. Жилкование крыльев кантаридоидного или стафилиноидного типа. У разных групп имеются различные особенности в жилковании. Некоторые виды утратили способность к полёту, их крылья редуцированные или полностью отсутствуют, а надкрылья срослись (Lamia, Morimus, Dorcadion)[1][13].
Конечности
 Структура лапки жуков-усачей на примере Callidium violaceum. T — голень, 1 — первый членик лапки, 2 — второй членик лапки, K — коготки; выделено цветами: красным — третий членик лапки, жёлтым — четвёртый членик лапки, зелёным — пятый членик лапки).
Структура лапки жуков-усачей на примере Callidium violaceum. T — голень, 1 — первый членик лапки, 2 — второй членик лапки, K — коготки; выделено цветами: красным — третий членик лапки, жёлтым — четвёртый членик лапки, зелёным — пятый членик лапки).Конечности состоят из пяти основных частей: тазиков, вертлугов, бёдер, голеней и члеников лапок. У усачей передние тазики имеют пулеобразную или конусовидную формы, средние — имеют шаровидную форму. К вершине бедра присоединяется голень, а к ней лапка с члениками. На вершине голени расположены две шпоры, а у некоторых видов — одна. Лапка пятичлениковая, но имеет вид четырёхчлениковой, поскольку четвёртый членик является недоразвитым и обычно представлен небольшим, часто едва заметным бугорком или узелком у основания 5-го членика, несущего коготки. Третий членик разделен на две части, и является двулопастным. Последний членик лапки несёт два коготка. Все членики лапки более или менее расширены, с густой щёточкой щетинок на нижней стороне[1].
Количество члеников лапок непостоянно, например, у Parandra и некоторых других дровосеков лапки пятичлениковые[1].
Первая пара ног самая короткая, а задняя — самая длинная. Однако передние ноги самцов некоторых видов, например, Acrocinus longimanus, Telotoma hayesi, могут достигать значительной длины до 170 мм, превышая длину самого тела жука[15][16].
Брюшко
Брюшко состоит из пяти свободных колец, с верхней стороны прикрытых надкрыльями. Они называются тергитами, а с нижней стороны — стернитами. По краю тергитов расположены дыхальца. Последний видимый 9-й тергит брюшка называется пигидием. Часто виден ещё и постпигидий — лучший и самый верный признак самца. Брюшко самок обычно массивнее и шире, чем у самцов, что связано с яйцекладущей функцией. Самки ряда видов имеют длинный хитиновый яйцеклад (Acanthocinus, Leiopus), а у некоторых видов брюшко самок может вытягиваться, образуя яйцеклад, превосходящий длину тела жука (Prionus kornarovi)[1].
Окраска
Окраска представителей семейства бывает самой разнообразной и состоит из окраски самого хитина и распределения волосков по телу, если такие имеется. Может быть любого цвета: от бледно-жёлтого и белого до смоляно-чёрного, и различных металлических оттенков. Часто может образоваться тот или иной рисунок, состоящий из пятен, полос и перевязей различной формы, размера и цвета[1]. Рисунок, образованный пятнами и полосами, часто бывает весьма изменчивым — полосы могут разбиваться на отдельные пятна, частично исчезать, или, наоборот, сливаться. Некоторые виды отличаются большим процентом вариабельных форм окраски. Меланизм широко распространён среди групп с надкрыльями, не несущими тот или иной рисунок и встречается преимущественно в горах[1]. Среди усачей он редок, так мало горных представителей этого семейства. Наоборот, меланизм можно встретить среди равнинных форм, например, степные Dorcadion очень часто бывают абсолютно чёрные.
 Усач-плотник. Самка (слева) и самец. Половой диморфизм выражен не только в размерах жуков и длине усиков, но и скульптуре переднеспинки самки.
Усач-плотник. Самка (слева) и самец. Половой диморфизм выражен не только в размерах жуков и длине усиков, но и скульптуре переднеспинки самки.Половой диморфизм
Половой диморфизм резко выражен, иногда настолько, что самец и самка одного вида мало похожи друг на друга. Самки отличаются от самца более крупным и массивным телом. Величина тела при этом не всегда связана с половой принадлежностью — самцы и самки могут не отличаться по размеру тела, иногда самцы крупнее самки благодаря развитой переднегруди (род Ergates) или мандибулам (Macrodontia), но в большинстве случаев самки крупнее, а в исключительных случаях, значительно крупнее самцов и обладают более широким телом, с более параллельными, менее суженными к вершине надкрыльями[1]. Половой диморфизм выражается в длине усиков (у самцов длиннее, чем у самок), гораздо реже в строении или количестве самих члеников[1].
У Prioninae имеет место различия в форме, размере и скульптуре переднеспинки (Ergates, Macrotoma, Rhesus). Также у самцов представителей этой группы мандибулы развиты значительно сильнее, чем самок. Также, периодически попадаются особи самцов т. н. «женственного типа», со слаборазвитыми челюстями, приближающимися по своему строению к челюстям самок[1].
Одним из проявлений полового диморфизма, у представителей семейства, являются различия в окраске особей различного пола — например, среди видов рода Anastrangalia имеются отличия в цвете надкрылий.
Особенности биологии
Питание
Имаго могут питаться пыльцой, листьями, хвоей, реже объедать кору на молодых веточках. Это питание является дополнительным, и нередко предшествует спариванию и является обязательным процессом, предшествующим созреванию половых клеток Особенно резко выражено у видов рода Monochamus — жуки объедают тонкие веточки, у представителей рода Пахиты|пахит Pachyta, у большого осинового скрипуна (Saperda carcharias), но не всегда резко выражено. Многие виды охотно прилетают на вытекающий древесный сок. Большинство Prioninae не питаются[1].
Сезонная и суточная активность
Время лёта и его продолжительность весьма разнообразны и ограничены тёплым временем года в умеренных широтах. Наиболее ранними формами на юге средней полосы России являются виды Dorcadion — преимущественно весенние формы[14]. Наибольшее число видов появляется в средней полосе в июне — июле, в южных областях — в мае — июне. В Средней Азии имеется несколько видов, лёт которых приходится на конец лета — начало осени. Жуки, питающиеся на цветах активны днём, и наиболее подвижны в жаркие часы дня (Lepturinae). Большинство Prioninae активны вечером и в сумерках, а днём прячутся в различных укромных местах. Вечером более подвижны и представители Cerambyx. Бескрылые Dorcadion активны днем, а на ночь прячутся в укрытия[1]. У одних видов более активными и склонными к перелётам являются самцы (например, дровосек-кожевник[14]), у других — наоборот, самки (например, реликтовый дровосек[17]).
Полёт
Полёт жуков довольно тяжёлый, особенно у крупных видов. Только немногие виды способны быстро и легко летать, взлетая с большой скоростью, например, Eutetrapha metallescens, которая в отличие от близких к ней видов Saperda очень подвижна и часто летает[1].
Межвидовое скрещивание
Межвидовое скрещивание у усачей не является редкостью. Для видов индо-малайского рода Batocera зарегистрирован ряд гибридов, также известны межвидовые гибриды и в роде Monochamus. По наблюдениям, в Малой Азии имеет место спаривание между различными видами рода Dorcadion и среди представителей подрода Compsodorcadion[1].
Морфология преимагиальных стадий
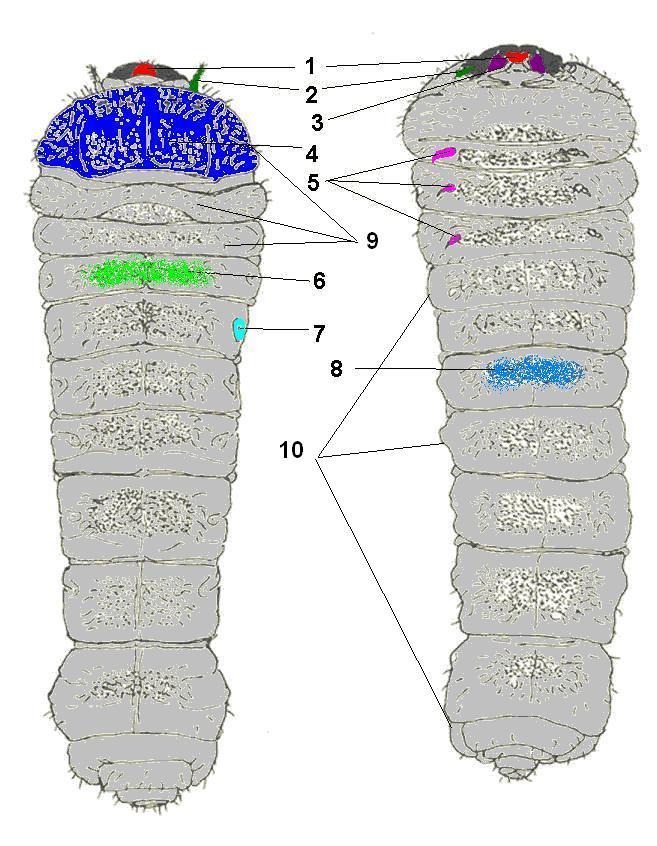
 Морфология личинки жука-усача на примере Phymatodes testaceus Обозначения: 1 — верхняя губа, 2 — усики (антенны), 3 — мандибулы (жвалы), 4 — пронотум (переднеспинка), 5 — ноги, 6 — дорсальные мозоли, 7 — дыхальца, 8 — вентральные мозоли, 9 — грудные сегменты, 10 — брюшные сегменты
Морфология личинки жука-усача на примере Phymatodes testaceus Обозначения: 1 — верхняя губа, 2 — усики (антенны), 3 — мандибулы (жвалы), 4 — пронотум (переднеспинка), 5 — ноги, 6 — дорсальные мозоли, 7 — дыхальца, 8 — вентральные мозоли, 9 — грудные сегменты, 10 — брюшные сегментыЛичинка
Тело личинок сплющено или более призматической формы, мясистое, белого или желтоватого цвета с более тёмной переднеспинкой и головой, последняя втянута в грудь с направленными вперёд челюстями. Переднегрудь сильно увеличена, заметно больше, чем среднегруди и заднегруди.
Изменилась форма тела у личинок стеблевых усачей (Agapanthia), обитающих в тонких стеблях и корнях травянистых растений — они длинные, тонкие, цилиндрической формы с увеличенными переднегрудью и последним сегментом тела, которые особенно густо покрыты волосками[1].
Голова представлена большой хитиновой капсулой. Простые глазки (от 1 до 5 пар) отсутствуют (Clytus, Agapanthia) или находятся у переднего края верхней стороны (Prionus, Spondylis, Callidium и др.), вблизи усиков. Усики очень короткие, трёхчлениковые, подогнутые книзу[1].
Брюшко состоит из 9 сегментов и анального соска, который можно посчитать за 10-й сегмент. Анальный сосок нередко бывает прикрыт сверху 9-м сегментом. Анальное отверстие трёхлопастное, но как исключение бывает в виде поперечной щели (Lamia iextor). Дыхательные отверстия овальные, в количестве 9 пар. Первая пара находится на среднегруди или сдвинута вперёд к месту сочленения передне- к среднегруди, несколько ниже остальных пар[1].
Грудные ноги у всех видов небольшие или рудиментарные, или отсутствуют. Все тело, за исключением головы и переднеспинки, покрыто мягкой кожей[1].
На верхней стороне сегментов тела имеются мозолистые бугорки по одному на каждом сегменте и по два овальных дыхальца. Ножки короткие, едва заметные. У эволюционно примитивных таксонов (Prionus, Rhagium, Gnathacmaeops и др.), на конце брюшка личинки, имеются крючки или шипы, которые помогают им перемещаться в выгрызенных ходах. Личинки же высокоорганизованных родов (Phytoecia, Agapanthia и т. п.) такое приспособление утратили.
В то же время в строении личинок различных групп наблюдаются очень резкие различия. Личинки Lamiinae безногие, с очень длиной и глубоко втянутой в переднегрудь головой, у личинок Lepiurttes голова очень широкая и в грудь втянута только слегка[1].
Куколка
 Куколка Rhagium mordax.
Куколка Rhagium mordax.Куколка усачей принадлежит к числу так называемых свободных куколок — все части её тела ясно различимы, зачатки крыльев, конечностей и усиков являются свободными (не сращенными), лишь прижатыми к телу[1][13]. Голова обычно подогнута под грудь, усики загнуты книзу между ногами, а если очень длинны, то загнуты ещё и вперёд. Цвет куколки беловатый или желтоватый. Характерной особенностью является наличие у большинства видов шипиков, шипов, зубчиков, либо толстых и коротких волос и щетинок на теле[1].
Окукливание у представителей различных таксономических групп имеет свои отличия и особенности. У большинства Prionus, а также других примитивных Lepturinae из трибы Rhagiini, личинка последнего возраста выходит из древесины в грунт, где сооружает камеру, в которой и окукливается. Похожее поведение характерно для Dorcadiini, но их личинки живут в грунту и там же окукливаются. Большая часть Cerambycinae, Lepturinae, Aseminae окукливаются в древесине — другие вгрызаются в толщу древесины, где из опилок строят камеру (Cerambyx cerdo, Monochamus, Hylotrupes и др.) другие окукливаются под корой (например, Rhagium). Прогрессивные трибы — Agapanthiini, Phytoeciini и т. п. — окукливаются в стеблях и корнях разнообразных трав[13].
Продолжительность стадии куколки обычно до 30—40 дней[1].
Развитие жуков-усачей
Большинство видов имеют одно- или двухлетний цикл развития. У разных групп усачей нет принципиальных отличий в их развитии, все они проходят стадии яйца, личинки, куколки и имаго. Вариации существуют лишь в брачном поведении и способах откладывания яиц, тем не менее существует несколько экологически отличных жизненных циклов. Первый — это развитие личинок в древесине, с дополнительным питанием на стадии имаго пыльцой цветов — антофилы (Nivellia, Carilia, Leptura и др.), вегетативными органами деревьев и их соком (Monochamus, Acanthocinus, Cerambyx и т. п.) или без дополнительного питания — афаги (Prionus, Hylotrupes и т. п.). Второй — это развитие личиночной стадии в стеблях травянистых растений из семейств Сложноцветные (Asteraceae) и Зонтичные (Apiaceae) с дополнительным питанием имаго вегетативными частями травянистых растений (Agapanthia, Phytoecia и др.). Третий — это есть развитие личиночной стадии в грунте с их питанием корнями злаков (Poaceae), а имаго дополнительно питаются их наземными частями (Dorcadion).
Большинство усачей развиваются в деревьях и кустарниках, но есть целые роды, где все виды связаны с травянистыми растениями[13]. Представители рода Phytoecia предпочитают небольшие травянистые растения, а рода Agapanthia — более крупные. Дальневосточный усач Thyestilla gebleri развивается в стеблях конопли[1]. Но более типично для усачей развитие личинок в древесине[13]. Существуют узкоспециализированные виды, которые заселяют деревья только одного рода (например, скрипун осиновый малый живёт только на тополях). Однако в различных географических районах предпочитаемые породы могут меняться, и в итоге для большинства видов усачей характерен довольно широкий круг кормовых пород. Широко распространена многоядность — один вид может жить на многих лиственных или хвойных породах, а также абсолютная всеядность, когда личинки одного вида могут жить и в хвойных, и в лиственных деревьях (дровосек-кожевник и чернопятнистый рагий) — но это те виды, личинки которых развиваются в сильно сгнившей древесине[13]. Личинки, живущие в деревьях, обитают сперва под корой, протачивая неправильные широкие ходы, наполняя их буровой мукой. Некоторые виды усачей-рагий (Rhagium) остаются все время под корой, другие делают многочисленные ходы в древесине (Saperda) или в сердцевине (Oberea oculata)[13]. Форма ходов различна у разных видов. Общим признаком является округлённо-овальная форма в поперечном сечении. Обычное направление ходов — более или менее продольное. У большинства личинок, ходы набиты буровой мукой, перемешанной с испражнениями личинок, и немногие виды делают по длине хода вентиляционные отверстия, через которые выбрасывают муку наружу[1].
В первом варианте жизненного цикла, самка откладывает яйца группами на поверхность или в трещины коры дерева. Такая кладка характерная для большинства представителей подсемейств Lepturinae, Prioninae и Cerambycinae, что считается очень примитивным признаком, ведь отсутствует любая забота о потомстве. У видов из подсемейства Lamiinae процесс кладки яиц более сложный. Самка, например, большого елового чёрного усача (Monochamus sartor) выгрызает на коре дерева специальную ямку — насечку, куда опускает яйцеклад и откладывает одно яйцо. Значительно более сложное поведение у скрипуна осинового (Saperda populnea) — самка на тоненьких веточках осины делает насечку, куда откладывает одно яйцо и прикрывает его выгрызенной ею стружкой из коры.
 Личинка усача-кожевника в куколочной колыбельке.
Личинка усача-кожевника в куколочной колыбельке.После этого самка по кругу обгрызает верхушку побега так, что через несколько дней она вянет, и к моменту выхода личинки из яйца высыхает и отпадает. Такие действия самки приводят к ослаблению побега, в котором будет развиваться личинка, и препятствует выделению растениями защитных веществ — терпенов. Личинки, после выхода из яйца, питаются лубом. Личинка скрипуна осинового (Saperda populnea) отличается от большинства своеобразным образом жизни. Она сначала питается новообразовавшейся тканью возле насечки, которую подготовила самка, потом делает узкий кольцевой периферийный ход в лубе. К осени, в большинстве случаев, углубляется в сердцевину, где прокладывает вверх короткий (приблизительно 4 см) вертикальный ход. Дерево реагирует на повреждение образованием «опухоли». У Lepturinae, Prioninae личинки развиваются в мёртвом дереве, которое начало гнить, заселено бактериями и грибами. Личинки большинства видов Lepturinae ассоциированы с грибами, смесью мицелия которых и древесины они питаются[13].
У видов, которые имеют двухлетнюю генерацию, личинка, в конце лета, вгрызается в глубину древесины, где зимует, а весной возвращается под кору, где продолжает питаться и расти, окукливаясь лишь осенью. У усачей с одногодичным жизненным циклом, личинка, в конце лета, также вгрызается в глубину древесины, где выгрызает камеру и окукливается. Весной или летом следующего года, взрослый жук проходит через личиночный туннель прогрызает кору дерева и выходит наружу. У видов, которые развиваются в корнях деревьев, личинки окукливаются в грунте, например, Prionus, Pachyta, Carilia и др. Личинка большого дубового усача (Cerambyx cerdo) живёт под корой, где, питаясь, прогрызает длинные извилистые хода. Перед окукливанием, она вгрызается в древесину, прокладывая короткий крючкообразный ход, который оканчивается камерой для окукливания. В этой камере личинка создаёт своеобразную стенку из мелкой тырсы, которая закрывает личиночный ход. Такая стенка обеспечивает стабильный температурный режим в камере и защищает от внешних врагов, которые могли бы попасть в ход. Куколка формируется осенью или в конце лета, и жук зимует в камере, выходя на поверхность лишь в следующем году, в начале лета, сперва прогрызая отверстие в личиночный ход, а затем кору.
Связь личинок усачей с грибами сложна и многообразна. Для личинок наиболее дефицитными веществами являются белки, которых ничтожно мало в древесине[13]. В природе усачи ищут древесину, в которой есть необходимый минимум белковых соединений. Мицелий грибов пронизывает всё толщу древесины и разрушает её, перерабатывая в свои богатые белком гифы[13]. Такая древесина более привлекательна для личинок усачей. Многие виды вступили с грибами в ещё более тесные симбиотические связи. В стенках их кишечника или жировом теле, имеются особые органы — мицетомы, служащие своеобразной средой обитания и размножения грибков, которые усваивают азот воздуха и превращают его в белковые соединения[13]. Личинки ряда видов способны усваивать до 20 % данной трудноусваиваемой пищи, благодаря наличию фермента целлюлозы[13]. У Xystocera globosa личинки развиваются только в живой древесине, содержащей не менее 10 % легко усваиваемых крахмала и сахаров[13].
Большинство видов связано с древесной растительностью, но есть группы, перешедшие к жизни в стеблях или корнях трав. Известны случаи, когда близкие виды из одного и того же рода усачей в лесной зоне развиваются в древесине, а в степях и пустынях встречаются в почве у корней растений[13].
Обычно самцы появляются на несколько дней раньше самок[1][13]. Вскоре после спаривания они умирают, а самки живут, пока не закончат кладку яиц. Растянутость кладки яиц сильно отражается на лёте жуков, и он часто может растягиваться на два — три месяца[1].
Происхождение
Жуки-усачи являются относительно древним семейством. Его представители, в ископаемом состоянии, известные из отложений юрского период[18]. Из этого периода описаны как вымершие, так и некоторые современные таксоны усачей[3]. Например, Prionus ooliticus, Cerambycinae dubius из Золенгофенского литографического сланца[3]. Таким образом, в верхней юре развили первоначальные формы усачей, из которых сформировалась всё семейство. Данные первые формы имели много общих черт с листоедами[3]. В нижнем меловом периоде первоначальные формы усачей достигают значительного многообразия и полностью отделяются от листоедов, что приводит к началу формирования высших таксонов семейства, а уже к середине мелового периода они вымирают. Их место занимают представители древних подсемейств Прионины, Парандрины, Лептурины и др. Данное события приходится на время появления первых покрытосеменных, древесину которых они заселяют. В третичном периоде усачи занимают практически все экологические ниши, которые существуют в условиях разлагающегося дерева и начинают заселять живые деревья, приобретая высокий уровень видового многообразия[3]. В третичных отложениях встречаются большей частью современные виды родов Leptura, Lamia, Astynomus и др., — найдены как имаго жуков, так и ходы их личинок в древесине породы Astynomus tertiarius[3]. С середины третичного периода появляются травянистые покрытосеменные растения, которые становятся отправной точкой к развитию многих новых таксонов, таких как Dorcadiini, Agapanthiini, Phytoeciini. Эти виды переходят к жизни в стеблях растений или грунте[3]. Середина третичного периода становится эпохой расцвета двудольных — значительного распространения приобретают зонтичные, сложноцветные и прочие, в стеблях которых поселяются усачи. Начало четвертичного периода и обледенение привели к вымиранию значительной части видов, распространённых в зонах умеренного климата. В Европе и Северной Азии исчезли третичные широколиственные леса, обусловив тем самым вымирание большинства здешних видов усачей[19]. Третичная реликтовая флора и фауна этих лесов сохранилась лишь на Дальнем Востоке и характеризуется значительным видовым многообразием[1][19].
Филогения
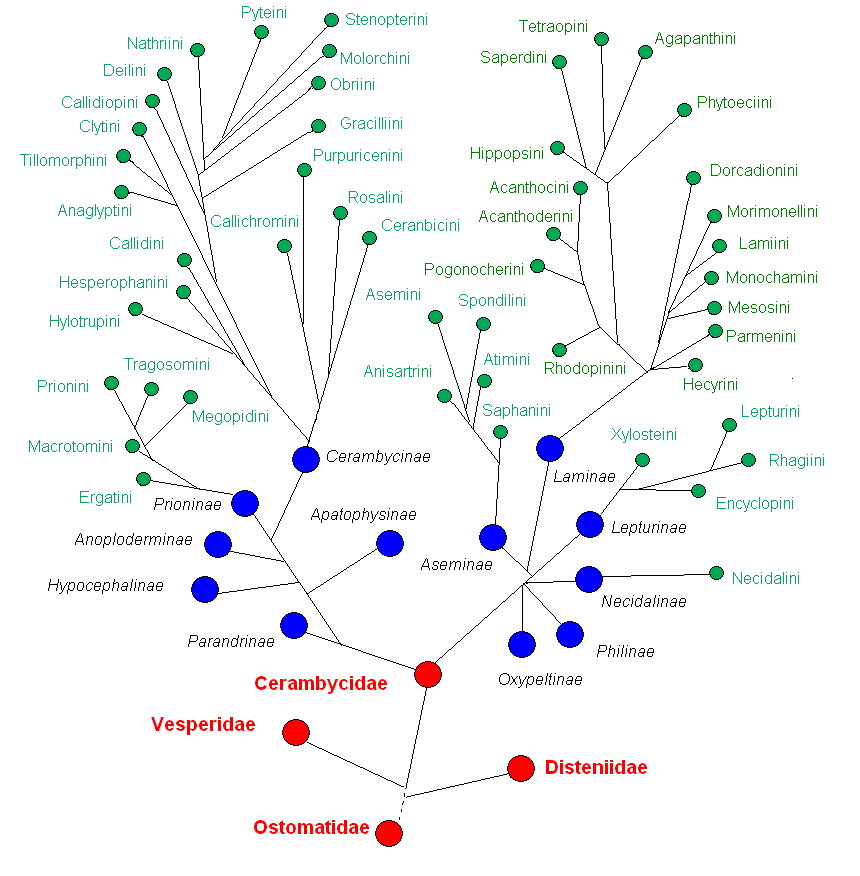
 Филогенетическое древо семейства жуков-усачей.
Филогенетическое древо семейства жуков-усачей.Жуков-усачей сближают с семействами Дистенииды (Disteniidae J. Thomson, 1860) и Vesperidae Mulsant, 1839 и включают в надсемейство Chrysomeloidea Latreille, 1802 вместе с такими семействами жуков как Chrysomelidae, Megalopodidae, Orsodacnidae и Oxypeltidae (Lawrence & Newton, 1995)[20].
Согласно современным представлениям семейство усачей включает две филогенетически отдалённые ветви[21]. Первая — это подсемейства Prioninae и Cerambycinae, которые происходят от общего предка, при этом подсемейство Prioninae является более примитивным и древним среди усачей. Как считалось ранее[3], вместе с этой ветвью развивалось подсемейства Parandrinae, Apatophysinae, Anoploderminae, и Hypocephalinae. Сегодня, подсемейство Parandrinae, рассматривают в качестве трибы Parandrini в составе подсемейства Prioninae. Подсемейство Apatophysinae переведено в ранг трибы Apatophysini и сведено в синонимы к Dorcasomini (Dorcasomini) в подсемействе Lepturinae. Anoploderminae и Hypocephalinae переведены в семейство Vesperidae. Иногда Anoploderminae выделяют в отдельное семейство[22]. Вторая филогенетическая ветвь семейства усачей включает центральный стержень Lepturinae. — Lamiinae, наряду с ними развивались небольшие подсемейства Necidalinae, Aseminae. Ранее[3], к этой группе относили подсемейства Oxypeltidae, Philinae. Сейчас Oxypeltidae относят к отдельному семейству Оксипельтины, а второе — к семейству Disteniidae[23].
Классификация
 Batocera rosembergi.
Batocera rosembergi. Самка Batocera wallacei.
Самка Batocera wallacei.Семейство усачей относят к надсемейству Хризомелоидные. Некоторые исследователи выделяют надсемейство Церамбикоидные (лат. Cerambycoidea), включая в него усачей и дистениид[24], но такое дробление не является общепринятым.
Оспаривается включение Vesperidae (Mulsant, 1839) Anoplodermatinae, Philinae, Vesperinae, как по строению личинок, так и по хромосомам[25].
- Подсемейство Aseminae (Thomson, 1860) — или в составе Spondylidinae.
- Подсемейство Cerambycinae (Latreille, 1802) — около 10 000 видов, 750 родов.
- Трибы: Achrysonini — Agallissini — Alanizini — Anaglyptini [Nomen Protectum] — Aphanasiini — Aphneopini — Auxesini — Basipterini — Bimiini — Bothriospilini — Brachypteromini — Callichromatini — Callidiini — Callidiopini — Cerambycini — Certallini — Chlidonini — Clytini — Compsocerini — Coptommatini — Curiini — Deilini — Dejanirini — Diorini — Distichocerini — Dodecosini — Eburiini — Ectenessini — Elaphidiini — Dryobiini — Eligmodermini — Erlandiini — Eroschemini — Eumichthini — Gahaniini — Glaucytini — Graciliini — Hesperophanini (подтрибы: Daramina — Hesperophanina) — Hesthesini — Heteropsini — Holopleurini — Holopterini — Hyboderini — Hylotrupini — Ibidionini — Ideratini — Lissonotini — Lygrini — Macronini — Megacoelini — Methiini — Molorchini — Mythodini — Necydalopsini — Neostenini — Obriini — Ochyrini — Oedenoderini — Oemini (подтрибы: Methioidina — Oemina) — Opsimini — Paraholopterini — Phalotini — Phlyctaenodini — Phoracanthini — Phyllarthriini — Piesarthriini — Piezocerini — Platyarthrini — Plectogastrini — Plectromerini — Pleiarthrocerini — Protaxini — Prothemini — Psebiini — Pseudocephalini — Psilomorphini — Pteroplatini — Pyrestini — Rhagiomorphini — Rhinotragini — Rhopalophorini — Rosaliini — Sestyrini — Smodicini — Spintheriini — Stenhomalini — Stenoderini — Stenopterini — Strongylurini — Tessarommatini — Thraniini — Thyrsiini — Tillomorphini — Torneutini — Trachyderini (подтрибы: Ancylocerina — Trachyderina) — Tragocerini — Trichomesiini — Tropocalymmatini — Typhocesini — Unxiini — Uracanthini — Vesperellini — Xystrocerini
- Подсемейство Disteniinae (Thomson, 1860)
- Трибы: Cyrtonopini — Disteniini — Dynamostini — Heteropalpini
- Подсемейство Dorcasominae (Lacordaire, 1868)
- Подсемейство Lamiinae (Latreille, 1802) — около 13 000 видов, более 700 родов.
- Трибы: Acanthocinini — Acanthoderini — Acmocerini — Acridocephalini — Acrocinini — Aderpasini — Aerenicini — Agapanthiini — Amphoecini — Ancitini — Ancylonotini — Anisocerini — Apomecynini — Astathini — Batocerini — Calliini — Ceroplesini (подтрибы: Ceroplesina — Crossotina) — Cloniocerini — Colobotheini — Compsosomatini — Cyrtinini — Desmiphorini — Dorcadionini — Dorcaschematini — Elytracanthinini — Enicodini — Eupromerini — Forsteriini — Gnomini — Gyaritini — Heliolini — Hemilophini — Homonoeini — Hyborhabdini — Lamiini — Laticraniini — Mauesiini — Megabasini — Mesosini — Microcymaturini — Moneilemini — Monochamini — Morimonellini — Morimopsini — Nyctimeniini — Obereini — Oculariini — Onciderini — Oncideropsidini — Onocephalini — Onychogleneini — Parmenini — Petrognathini — Phacellini — Phantasini — Phrynetini — Phymasternini — Phytoeciini — Pogonocherini — Polyrhaphidini — Pretiliini — Proctocerini — Prosopocerini — Pteropliini — Rhodopinini — Saperdini — Stenobiini — Sternotomini — Tapeinini — Tetraopini — Tetraulaxini — Theocridini — Tmesisternini — Tragocephalini — Xenicotelini — Xenofreini — Xenoleini — Xylorhizini — Zygocerini
- Подсемейство Lepturinae (Latreille, 1802) — около 200 видов.
- Трибы: Desmocerini — Encyclopini — Lepturini — (Necydalini) — Oxymirini — Rhagiini — Rhamnusiini — Teledapini — Xylosteini
- Подсемейство Necydalinae (Latreille) Pascoe, 1864 (или Necydalini в составе Lepturinae)
- Подсемейство Oxypeltinae (Lacordaire, 1869)
- Подсемейство Parandrinae (Latreille, 1802)
- Трибы: Parandrini — Erichsoniini
- Подсемейство Prioninae (Latreille, 1802) — около 1000 видов.
- Трибы: Acanthophorini — Aegosomatini — Anacolini — Cacoscelini — Callipogonini — Calocomini — Cantharocnemini — Ergatini — Eurypodini — Hopliderini — Macrodontiini — Macrotomini (подтрибы: Archetypina — Basitoxina — Macrotomina — Mallodonina — Platygnathina — Remphanina — Xixuthrina) — Mallaspini — Meroscelisini — Prionini — Solenopterini — Tereticini — Vesperoctenini
- Подсемейство Spondylidinae (Serville, 1832)
- Трибы: Anisarthronini — Asemini — Atimiini — Saphanini — Spondylidini
- Подсемейство Anoplodermatinae (Guérin-Méneville, 1840; принадлежность к Vesperidae или Prioninae окончательно не определена)
- Трибы: Anoplodermatini — Hypocephalini — Mysteriini
- Подсемейство Philinae (Thomson, 1860; принадлежность к Vesperidae или в Prioninae окончательно не определена)
- Подсемейство Vesperinae (Mulsant, 1839; принадлежность к Vesperidae или Lepturinae окончательно не определена)
Роль в экосистемах
Роль усачей в естественных экосистемах заключается в том, что, будучи, консументами первого порядка, они являются неотъемлемыми компонентами цепей питания[26]. В процессе коэволюции организмов, сформировалась трофическая сеть связей между автотрофами и гетеротрофами, в которую усачи включаются на стадиях личинки, куколки и имаго. Поскольку личинки и куколки большинства видов развиваются в такой консервативной среде как древесина, где условия развития не менялись на протяжении десятков и сотен миллионов лет, они являются звеньями в питании птиц и паразитических насекомых-энтомофагов. Имаго включаются в трофическую сеть значительно большего числа консументов второго порядка — это разнообразные птицы, пресмыкающиеся, хищники, мелкие млекопитающие и т. п.
 Личиночные ходы большого дубового усача.
Личиночные ходы большого дубового усача.Консортивные связи с древесными растениями
Как ксилофаги — личинки большинства видов усачей, за редким исключением, развиваются в древесине, являясь утилизаторами погибших хвойных и лиственных пород деревьев. Жуки-усачи, в значительной мере опосредованно, принимают участие в минеральном питании растений, которое выражается в поверхностном поступлении зольных элементов и азота из экскрементов и стимуляцией деструктивных процессов. В сущности, усачи вместе с древесными растениями, в древесине которых они развиваются, образовывают популяционные консорции, где их функциональная роль состоит в освобождении, в процессе деструкции мёртвой органики, элементов, которые будут использованы для формирования живой органической массы детерминанта консорции[27].
Санитарная роль в лесных экосистемах
В естественных и незначительной мере изменённых экосистемах, усачи выполняют санитарную роль, элиминируя мёртвую древесину и утилизируя ослабленные и придавленные конкурентной борьбой за свет, сухостои, поражённые корневым грибком (Heterobasidion annosum), трутовиками Phomitopsis pinicola, Phomitopsis chrysoloma, Polystictus circinatus, корневыми гнилями деревья, в определённой мере регулируя распространение этих грибов. Особую роль они сыграют на буреломах и вырубах, где ускоряют разложение мёртвого дерева[28]. Элиминация таких деревьев освобождает место для молодой поросли и способствует восстановлению экосистем.
 Усач мускусный (Aromia moschata), кормящийся на цветке.
Усач мускусный (Aromia moschata), кормящийся на цветке.Опыление цветковых растений
Усачи также выступают опылителями многих видов цветковых растений, ведь значительная часть этих насекомых характеризуется развитием антофилии. В частности, антофильными являются большая часть видов подсемейства Lepturinae, в меньшей степени виды из подсемейства Cerambycinae. Имаго этих видов часто встречаются на цветах, где в отличие от «классических» опылителей (пчелы, шмели, чешуекрылые, двукрылые и т. п.) они проводят значительно больше времени, и соответственно эффективность опыления оказывается выше. Также эти виды усачей являются регуляторами численности тех цветочных растений, которые они опыляют. Это связано с тем, что взрослым насекомым для жизнедеятельности и созревание яиц у самок требуется питание пыльцой, при этом жуки часто поедают гинецей и андроцей, уменьшая продукцию семян растений. Как правило, растения, которые опыляются усачами, производят большое количество семян и поедание их генеративных органов не оказывает существенного влияние на их репродукцию[14].
Экономическое значение
 Личиночные ходы и имаго вида Anoplophora glabripennis, который вместе с пиломатериалами из Китая распространился по всему миру
Личиночные ходы и имаго вида Anoplophora glabripennis, который вместе с пиломатериалами из Китая распространился по всему мируУсачей относят к одним из наиболее серьёзных вредителей древесных стройматериалов[29]. Усачей-вредителей разделяют на физиологически-технических вредителей — виды, которые повреждают живые и ослабленные деревья, и технических вредителей — виды, личинки которых заселяют уже мёртвое дерево[19]. Именно личинка у усачей является основной вредящей стадией в жизненном цикле насекомого[1][30].
Классическим примером усачей-вредителей является массовое размножение большого дубового усача (Cerambyx cerdo) в 1909—1930 гг. на Центральной и Северной Украине. В период с 1872 по 1930 года на этих местностях была внедрена вырубка леса полосами, с шириной полосы от 40 до 80 м, также оставлялись одиночные крупные дубы. Это привело к ослаблению оставленных деревьев, в виду изменения микроклимата, который под пологом леса характеризуется более постоянными показателями влажности и меньшими амплитудами суточных температур, чем на открытой местности. Ослабленные деревья становились привлекательными для заселения усачами, что и послужило причиной вспышки численности вида. Мероприятия борьбы в таких случаях являются малоэффективными[19]. На сегодняшний день большой дубовый усач считается редчайшим видом и занесён в охраняемые списки и Красные книги во всех европейских странах, в том числе и Украины[19].
Ещё одним хрестоматийным примером является вспышка численности большого елового усача в Томской области и Красноярском крае (Российская Федерация), которая наблюдалась в 1961 году[1][31]. В результате широкомасштабных лесоразработок площадью около 50 000 га наблюдалось массовое размножение этого вида усача. Оно было обусловлено неправильной хозяйственной деятельностью: одним из мероприятий, принятых для решения этой проблемы, стало ускорение вырубок и увеличение их площадей. Такая деятельность привела к быстрому росту численности данного вида и гибели леса сибирской тайги на множестве территорий. Характерными признаками было появление значительных по площади повреждённых участков леса, на которых наблюдалось пожелтение и опадание хвои сибирской лиственницы (Larix sibirica), сибирской ели (Picea sibirica) и сибирской пихты (Abies sibirica)[14]. Кроме того, взрослые насекомые откладывали яйца в ослабленные деревья, а развитие личинки служило причиной гибели последних. Эффект массового размножения большого елового чёрного усача в этом регионе наблюдался в течение 10 лет, пока не были прекращены все лесоразработки.
 Личинка домового усача (Hylotrupes bajulus), обычно обитающая в деревянных конструкциях домов.
Личинка домового усача (Hylotrupes bajulus), обычно обитающая в деревянных конструкциях домов.Некоторые виды усачей превратились в «спутников» человека, как, например, домовой усач (Hylotrupes bajulus)[14]. В естественных условиях личинки данного вида очень редки, но часто заселяют деревянные дома, перегородки, доски пола, оконные рамы, стропила потолка и крыши и даже мебель. Внешне жизнедеятельность личинок заметна только по появлению порошкообразной бурой деревянной пыли. Известны случаи, когда при массовом размножении домовой усач приводил в негодность деревянные постройки целых городских кварталов[13]. На Кавказе широко распространён усач Stromatium fulvum, от которого страдают практически все деревянные постройки[14]. На Украине, на Черноморском побережье Кавказа, в Ферганской и смежных областях сильно повреждает постройки и изделия из дерева усач Фальдермана (Chlorophorus faldermanni)[1][14].
С целью борьбы с усачами-вредителями хозяйственных растений и лесного хозяйства используется как классическая фумигация инсектицидами, так и биологические, а также биохимически-физологические методы. Одновременно, разработка таких методов предоставляет материал для научных исследований более фундаментального характера. Для разработки методов борьбы с жуком Xylotrechus pyrrhoderus, который является одним из наиболее опасных вредителей винограда в Юго-Восточной Азии, была выделена и привита к постоянному автономному существованию in vitro клеточная линия XP-1 из клеток его жирового тела, на которой потом тестировалась цитологическая чувствительность к нескольким гормонам[32]. Ещё одна стабильная клеточная линия усачей, РС-1, была получена японскими исследователями из жука Plagionotus christophi в 2008 году[33].
См. также
Литература
- Данилевский М. Л., Мирошников А. И., 1985. Жуки-дровосеки Кавказа (Coleoptera, Cerambycidae). Определитель. — Краснодар. — 419 с.
- Загайкевич И. К. Таксономия и экология усачей. — К.: Наукова думка, 1991. — 420 с.
- Лобанов А. Л., Данилевский М. Л., Мурзин С. В., 1981. Систематический список усачей (Coleoptera, Cerambycidae) фауны СССР. 1. — Энтомологическое обозрение, 60, 4: 784—803.
- Мамаев Б. М., Данилевский М. Л., 1975. Личинки жуков-дровосеков. — М.: Наука. — 282 стр.
- Плавильщиков Н. Н. Фауна СССР. Жесткокрылые. Т. XXI. Жуки-дровосеки (Cerambycidae). Часть 1. 1936. — Изд. АН СССР, М.-Л. — 612 с.
- Плавильщиков Н. Н., 1940. Жуки-дровосеки, ч. 2. — Фауна СССР, Насекомые жесткокрылые, т. 22. — Изд. АН СССР, М.-Л. — 785 с.
- Плавильщиков Н. Н., 1958. Жуки-дровосеки, ч. 3. — Фауна СССР, Насекомые жесткокрылые, т. 23, вып. 1. — Изд. АН СССР, М.-Л. — 592 с.
- Черепанов А. И., 1979. Усачи Северной Азии, том 1 (Prioninae — Aseminae). — Новосибирск: издательство «Наука», Сибирское отделение. — 470 с. 296 илл.
- Черепанов А. И., 1981. Усачи Северной Азии, том 2 (Cerambycinae, 1 часть). — Новосибирск: издательство «Наука», Сибирское отделение. — 216 с. 127 илл.
- Yves Bousquet, Daniel J. Heffern, Patrice Bouchard & Eugenio H. Nearns. Catalogue of family-group names in Cerambycidae (Coleoptera) (англ.) // Zootaxa. — Auckland, New Zealand: Magnolia Press, 2009. — В. 2321. — С. 1—80. — ISSN 1175-5326.
- Monné, M. A., 2005. Catalogue of the Cerambycidae (Coleoptera) of the Neotropical Region. Part I. Subfamily Cerambycinae. Zootaxa, 946: 1—765.
- Linsley, E. G., 1962. The Cerambycidae of North America. Part III. Taxonomy and classification of the subfamily Cerambycinae, tribes Opsimini through Megaderini. University of California Publications in Entomology, 20: 1—188.
- Linsley E. G., Chemsak J. A., 1984. Cerambycidae of North America. Part 7, No. 1. Taxonomy and Classification of the Subfamily Lamiinae, Tribes Parmenini through Acanthoderini. — Berkeley: University of California Press, 1984. Vol. 102. 1—258 p.
- Linsley E. G., Chemsak J. A., 1995. The Cerambycidae of North America. Part 7, No. 2: Taxonomy and Classification of the Subfamily Lamiinae, Tribes Acanthocinini through Hemilophini. — Berkeley: University of California Press, 1995. Vol. 114. 1—292 p.
- Sama G., 2002. Atlas of the Cerambycidae of Europe and the Mediterranean Area. Vol. 1. — Nakladatelství Kabourek. — Zlín. — 173 pp.
- Svacha P., Danilevsky M. L., 1987. Cerambycoid larvae of Europe and Soviet Union (Coleoptera, Cerambycoidea). Part I. — Acta Univ. Carolinae, 30 (1986), 1—2: 1—177.
- Svacha P., Danilevsky M. L., 1988. Cerambycoid larvae of Europe and Soviet Union (Coleoptera, Cerambycoidea). Part II. — Acta Univ. Carolinae, 31 (1987), 3—4: 121—284.
- Svacha P., Danilevsky M. L., 1989. Cerambycoid larvae of Europe and Soviet Union (Coleoptera, Cerambycoidea). Part III. — Acta Univ. Carolinae, 32 (1988), 1—2: 1—205.
Источники
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 Плавильщиков Н. Н. Жесткокрылые. Жуки-дровосеки (Cerambycidae). — М., 1936. — Т. XXI, часть 1. — (Фауна СССР).
- ↑ C. J. Micheli Studies on the longhorned woodboring beetles (Coleoptera: Cerambycidae) of the West Indies // Mast. of Scien. Thesis Univ. of Maryland — 2006. — 175 р.
- ↑ 1 2 3 4 5 6 7 8 9 Загайкевич И. К. Таксономия и экология усачей. — Киев: Наукова думка, 1991. — 420 с.
- ↑ Danilevsky M. L. Taxonomic and zoogeographical notes on the family Cerambycidae (Coleoptera) of Russia and adjacent regions. — Moscow: Moscow, 1993. — Т. vol. 1, No. 2. — P. 37—39. — (Russian entomological journal).
- ↑ Бартенев А. Ф. Обзор видов жуков-усачей (Coleoptera: Cerambycidae) фауны Украины // Вісник Харківського ентомологічного товариства. — 2003 (2004). — Т. 1—2. — № 11. — С. 24—43.
- ↑ Vanessa Heque. Longicornes de Guyane. — Cayenne, 1996.
- ↑ Mares, Lapacek, 1980
- ↑ http://www.insectworld.com — Macrodontia cervicornis M A1 16.5 cm. Архивировано из первоисточника 31 января 2012. Проверено 5 ноября 2009.
- ↑ Yanega, Douglas; David Olson, Sharon Shute, Ziro Komiya (2004). «The Xixuthrus species of Fiji (Coleoptera: Cerambycidae: Prioninae». Zootaxa (Magnolia Press) 777: 1—10. ISSN 1175-5326.
- ↑ Evenhuis, Neal L. (2007). «Note: Clarification of the Authorship of Xixuthrus heros (Coleoptera: cerambycidae)». Bishop Museum Occasional Papers 91: 39—42.
- ↑ http://www.cerambyx.uochb.cz. Архивировано из первоисточника 31 января 2012. Проверено 5 ноября 2009.
- ↑ Г.Н. Горностаев. Насекомые СССР. — Москва: Мысль, 1970. — 372 с. — (Справочники-определители географа и путешественника).
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 Жизнь животных. Энциклопедия в шести томах. Том 3. Членистоногие. Общая редакция члена-корреспондента АН СССР профессора Л. А. Зенкевича. — Москва: Просвещение, 1969. — 576 стр.
- ↑ 1 2 3 4 5 6 7 8 9 10 11 12 Шаблиовский В. В. Жуки-дровосеки лесной зоны СССР. — Л.: Наука, 1967. — 44 с.
- ↑ David W. Zeh, Jeanne A. Zeh und Melvin M. Bonilla: Phylogeography of the giant harlequin beetle (Acrocinus longimanus). Journal of Biogeography, 30, 747—753, Oxford 2003 ISSN 0305-0270
- ↑ David W. Zeh, Jeanne A. Zeh und Gerard Tavakilian: Sexual Selection and Sexual Dimorphism in the Harlequin Beetle Acrocinus longimanus. Biotropica, 24(1): 86—96, Oxford 1992 ISSN 0006-3606
- ↑ Красная книга СССР. Редкие и находящиеся под угрозой исчезновения виды животных и растений. — М.: Лесная промышленность, 1978. — 460 с.
- ↑ Ponomarenko A. G. The geological history of beetles // in: J. Pakaluk & S. A. Ślipiński (eds.). Biology, Phylogeny, and Classification of Coleoptera. Papers celebrating the 80th Birthday of Roy A. Crowson. — Warszawa: Muz. Inst. Zool. PAN. — 1995. — Vol. 1. — P. 155—172.
- ↑ 1 2 3 4 5 Шаблиовский В. В. Жуки-дровосеки лесной зоны СССР // Автореф. дисс. д-ра биол. наук: 03.00.09. — Л., 1967. — 44 с.
- ↑ Lawrence, J. F.; Newton, A. F., jr. 1995: Families and subfamilies of Coleoptera (with selected genera, notes, references and data on family-group names). Pp. 779—1006 in Pakaluk, J.; Ślipiński, S. A. (eds.) Biology, phylogeny, and classification of Coleoptera: papers celebrating the 80th birthday of Roy A. Crowson. Museum i Instytut Zoologii PAN, Warszawa
- ↑ Napp D. S. 1994. — Phylogenetic relationships among the subfamilies of Cerambycidae (Coleoptera, Chrysomelidea). — Revista Brasileira de Entomologia 28 (2): 265—419
- ↑ Jenis I., 2001. — Long-Horned Beetles Distenidae, Oxypeltidae, Vesperidae, Anoplodermatidae & Cerambycidae I, Vesperidae and Cerambycidae of Europe I, A. Regulus, Zlin, 333 pp.
- ↑ Švacha P., Wang J. J. & Chen S. C. Larval morphology and biology of Philus antennatus and Heterophilus punctulatus, and systematic position of the Philinae (Coleoptera: Cerambycidae and Vesperidae). Annales de la Societé entomologique de France (N. S.). — 1997. — 33 (3): 323—369.
- ↑ Список усачей (Cerambycoidea) бывшего СССР
- ↑ Dutrillaux A. M., Moulin S. & Dutrillaux B., 2007 — Présence d’un caryotype très original a 53—54 chromosomes chez Vesperus xatarti Mulsant 1839 (Coleoptera: Cerambycidae: Vesperinae) — Annales de la Société Entomologique de France (N. S.) 43 (1): 81—86. PDF [1]
- ↑ Сезонні флуктуації видового багатства жуків-вусачів (Coleoptera: Cerambycidae) в лісових екосистемах північно-східного макросхилу Українських Карпат та Передкарпаття // Питання біоіндикації та екології. — Запоріжжя: ЗНУ, 2007. — В. 12. — С. 125—133.
- ↑ Царик Й. В., Царик І. Й. Консорція як один із базових рівнів біологічного різноманіття // Карпатський регіон і проблеми сталого розвитку. — Рахів, 1998. — С. 303—304.
- ↑ Wermelinger B., Duelli P. Obrist M. K. Dynamics of saproxylic beetles (Coleoptera) in windthrow areas in alpine spruce forests // For. Snow Landsc. Res. — 2002. — Vol. 77, ½. — pp. 133—148
- ↑ Шаблиовский В. В. Жуки-дровосеки Приморского края. — Л., 1950. — С. 15.
- ↑ Данилевский М. Л. Морфо-экологические закономерности эволюции личинок жуков-дровосеков. — Автореф. дисс. канд. биол. наук, 1967. — С. 17.
- ↑ Рожков А. А. Про большого елового усача // Химия и жизнь : журнал. — 1976. — № 12. — С. 88—92.
- ↑ K. Iwabuchi. An Established Cell Line from the Beetle, Xylotrechus Pyrrhoderus (Coleoptera: Cerambycidae) // In Vitro Cell. Dev. Biol.-Animal. — 1999. — Vol. 35. — pp. 612—615
- ↑ K. Hoshino & M. Hirose & K. Iwabuchi. A new insect cell line from the longicorn beetle Plagionotus christophi (Coleoptera: Cerambycidae) // In Vitro Cell. Dev. Biol.-Animal. — 2008. DOI 10.1007/s11626-008-9152-7
Ссылки
- Усачи (Cerambycidae) или дровосеки (включая Philinae, Vesperinae; но исключая Disteniidae)
- Иллюстрированный указатель триб к атласу жуков-усачей (Cerambycidae) России
- Иллюстрированный указатель родов жуков-усачей (Cerambycidae) фауны России и близлежащих стран
- Усачи Урала
- Усачи на сайте coleop-terra.com (англ.)
- Усачи Палеарктики (англ.)
- Усач западной Палеарктики (англ.)
- Фотогалерея Станислава Крейчика (англ.)
- Фотогалерея усачей мира (англ.)
- Личинки и куколки усачей (польск.)
- Tree of life Cerambycidae (англ.)


Эта статья входит в число хороших статей русскоязычного раздела Википедии. Категории:- Животные по алфавиту
- Усачи
- Семейства насекомых




















Wikimedia Foundation. 2010.