- КЛАСС ИНФУЗОРИИ (INFUSORIA или CILIATA)
- КЛАСС ИНФУЗОРИИ (INFUSORIA или CILIATA)
-
Простейшие этого обширного по количеству видов — около 6 тыс.— класса широко распространены в природе. (Эта цифра приводится в сводке Корлисса, 1961 г.). К ним относятся многочисленные обитатели морских и пресных вод. Некоторые виды приспособились к жизни во влажной почве. Немалое количество видов инфузорий ведет паразитический образ жизни. Хозяевами для паразитических инфузорий являются самые разнообразные беспозвоночные и позвоночные животные до высших обезьян и человека включительно.По сравнению с другими группами простейших инфузории имеют наиболее сложное строение, что связано с разнообразием и сложностью их функций.Откуда взялось название «инфузория туфелька»? Вы не будете удивлены, если взглянете под микроскопом на живую инфузорию или даже на ее изображение (рис. 85).
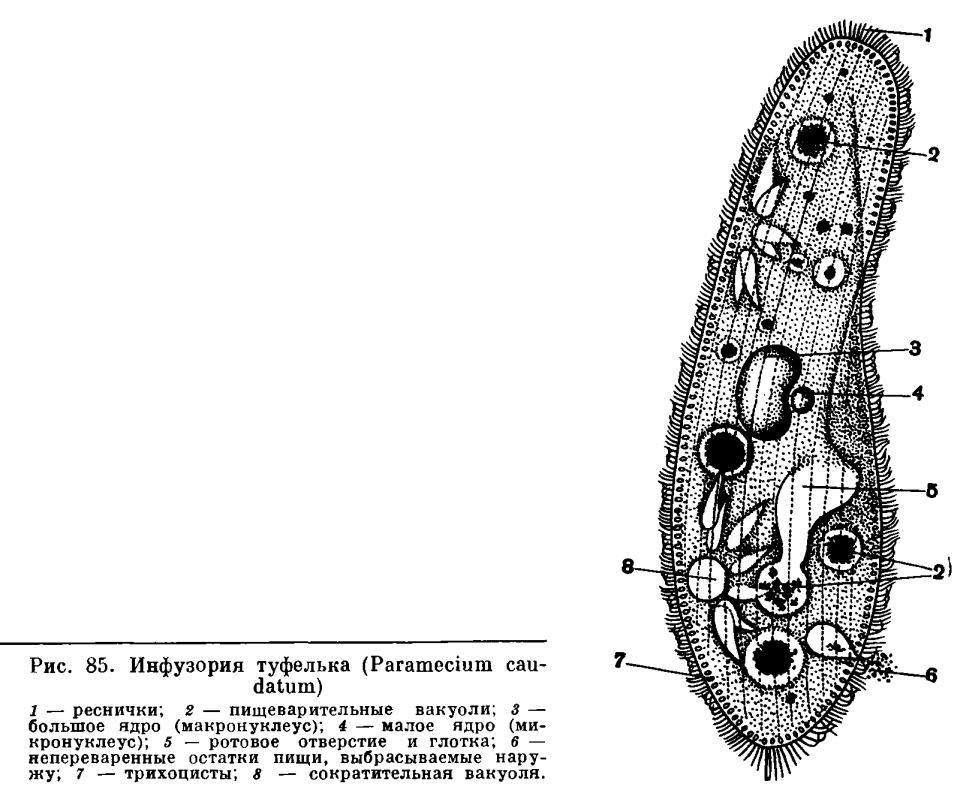 Действительно, форма тела этой инфузории напоминает изящную дамскую туфельку.Инфузория туфелька находится в непрерывном довольно быстром движении. Скорость его (при комнатной температуре) около 2, 0—2, 5 мм/сек. Для такого маленького животного это большая скорость! Ведь это означает, что за секунду туфелька пробегает расстояние, превышающее длину ее тела в 10—15 раз. Траектория движения туфелькд довольно сложна. Она движется передним концом прямо впередИНФУЗОРИЯ ТУФЕЛЬКА (PARAMECIUM CAUDATUM)Чтобы ознакомиться со строением и образом жизни этих интересных одноклеточных организмов, обратимся сначала к одному характерному примеру. Возьмем широко распространенных в мелких пресноводных водоемах инфузорий туфелек (виды рода Paramecium). Этих инфузорий очень легко развести в небольших аквариумах, если залить прудовой водой обычное луговое сено. В таких настойках развивается множество различных видов простейших и почти всегда развиваются инфузории туфельки. При помощи обычного учебного микроскопа можно рассмотреть многое из того, о чем будет дальше рассказано.Среди простейших инфузории туфельки являются довольно крупными организмами. Длина тела их около 1/6—1/3 мм. и при этом вращается вправо вдоль продольной оси тела.Столь активное движение туфельки зависит от работы большого количества тончайших волосковидных придатков — ресничек, которые покрывают все тело инфузории. Количество ресничек у одной особи инфузории туфельки равняется 10—15 тыс.!Каждая ресничка совершает очень частые веслообразные движения — при комнатной температуре до 30 биений в секунду. Во время удара назад ресничка держится в выпрямленном положении. При возвращении же ее в исходную позицию (при движении вниз) она движется в 3—5 раз медленнее и описывает полукруг.При плавании туфельки движения многочисленных покрывающих ее тело ресничек суммируются. Действия отдельных ресничек оказываются согласованными, в результате чего получаются правильные волнообразные колебания всех ресничек. Волна колебания начинается у переднего конца тела и распространяется назад. Одновременно вдоль тела туфельки проходят 2—3 волны сокращения. Таким образом, весь ресничный аппарат инфузории представляет собой как бы единое функциональное физиологическое целое, действия отдельных структурных единиц которого (ресничек) тесно связаны (координированы) между собой.Строение каждой отдельной реснички туфельки, как показали электронно микроскопические исследования, является весьма сложным.Направление и быстрота движения туфельки не являются величинами постоянными и неизменными. Туфелька, как и все живые организмы (мы видели это уже на примере амебы), реагирует на изменение внешней среды изменением направления движения.Изменение направления движения простейших под влиянием различных раздражителей называют таксисами. У инфузорий легко наблюдать различные таксисы. Если в каплю, где плавают туфельки, поместить какое-либо неблагоприятно действующее на них вещество (например, кристаллик поваренной соли), то туфельки уплывают (как бы убегают) от этого неблагоприятного для них фактора (рис. 86).
Действительно, форма тела этой инфузории напоминает изящную дамскую туфельку.Инфузория туфелька находится в непрерывном довольно быстром движении. Скорость его (при комнатной температуре) около 2, 0—2, 5 мм/сек. Для такого маленького животного это большая скорость! Ведь это означает, что за секунду туфелька пробегает расстояние, превышающее длину ее тела в 10—15 раз. Траектория движения туфелькд довольно сложна. Она движется передним концом прямо впередИНФУЗОРИЯ ТУФЕЛЬКА (PARAMECIUM CAUDATUM)Чтобы ознакомиться со строением и образом жизни этих интересных одноклеточных организмов, обратимся сначала к одному характерному примеру. Возьмем широко распространенных в мелких пресноводных водоемах инфузорий туфелек (виды рода Paramecium). Этих инфузорий очень легко развести в небольших аквариумах, если залить прудовой водой обычное луговое сено. В таких настойках развивается множество различных видов простейших и почти всегда развиваются инфузории туфельки. При помощи обычного учебного микроскопа можно рассмотреть многое из того, о чем будет дальше рассказано.Среди простейших инфузории туфельки являются довольно крупными организмами. Длина тела их около 1/6—1/3 мм. и при этом вращается вправо вдоль продольной оси тела.Столь активное движение туфельки зависит от работы большого количества тончайших волосковидных придатков — ресничек, которые покрывают все тело инфузории. Количество ресничек у одной особи инфузории туфельки равняется 10—15 тыс.!Каждая ресничка совершает очень частые веслообразные движения — при комнатной температуре до 30 биений в секунду. Во время удара назад ресничка держится в выпрямленном положении. При возвращении же ее в исходную позицию (при движении вниз) она движется в 3—5 раз медленнее и описывает полукруг.При плавании туфельки движения многочисленных покрывающих ее тело ресничек суммируются. Действия отдельных ресничек оказываются согласованными, в результате чего получаются правильные волнообразные колебания всех ресничек. Волна колебания начинается у переднего конца тела и распространяется назад. Одновременно вдоль тела туфельки проходят 2—3 волны сокращения. Таким образом, весь ресничный аппарат инфузории представляет собой как бы единое функциональное физиологическое целое, действия отдельных структурных единиц которого (ресничек) тесно связаны (координированы) между собой.Строение каждой отдельной реснички туфельки, как показали электронно микроскопические исследования, является весьма сложным.Направление и быстрота движения туфельки не являются величинами постоянными и неизменными. Туфелька, как и все живые организмы (мы видели это уже на примере амебы), реагирует на изменение внешней среды изменением направления движения.Изменение направления движения простейших под влиянием различных раздражителей называют таксисами. У инфузорий легко наблюдать различные таксисы. Если в каплю, где плавают туфельки, поместить какое-либо неблагоприятно действующее на них вещество (например, кристаллик поваренной соли), то туфельки уплывают (как бы убегают) от этого неблагоприятного для них фактора (рис. 86).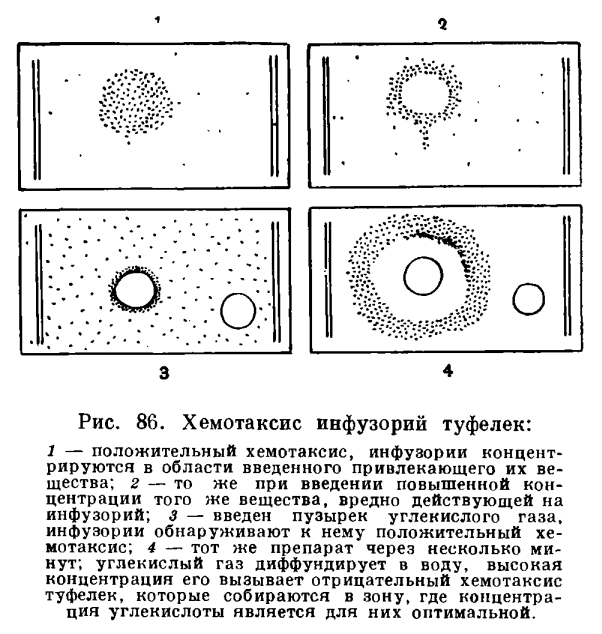 Перед нами пример отрицательного таксиса на химическое воздействие (отрицательный хемотаксис). Можно наблюдать у туфельки и положительный хемотаксис. Если, например, каплю воды, в которой плавают инфузории, прикрыть покровным стеклышком и подпустить под него пузырек углекислого газа (С02), то большая часть инфузорий направится к этому пузырьку и расположится вокруг него кольцом.Очень наглядно явление таксиса проявляется у туфелек под влиянием электрического тока. Если через жидкость, в которой плавают туфельки, пропустить слабый электрический ток, то можно наблюдать следующую картину: все инфузории ориентируют свою продольную ось параллельно линии тока, а затем, как по команде, двинутся в направлении катода, в области которого и образуют густое скопление. Движение инфузорий, определяемое направлением электрического тока, носит название гальванотаксиса. Различные таксисы у инфузорий могут быть обнаружены под влиянием самых разнообразных факторов внешней среды.Все цитоплазматическое тело инфузории отчетливо распадается на 2 слоя: наружный — более светлый (эктоплазма) и внутренний —' более темный и зернистый (эндоплазма). Самый поверхностный слой эктоплазмы образует наружную очень тонкую и вместе с тем прочную и эластичную оболочку — пелликулу, которая играет важную роль в сохранении постоянства формы тела инфузории.В наружном слое (в эктоплазме) тела живой туфельки хорошо видны многочисленные коротенькие палочки, расположенные перпендикулярно к поверхности (рис. 85, 7). Эти образования носят название трихоцисты. Функция их очень интересна и связана с защитой простейшего. При механическом, химическом или каком-либо ином сильном раздражении трихоцисты с силой выбрасываются наружу, превращаясь в тонкие длинные нити, которые поражают хищника, нападающего на туфельку. Трихоцисты представляют собой мощную защиту. Они располагаются закономерно между ресничками, так что число трихоцист приблизительно соответствует числу ресничек. На месте использованных («выстреленных») трихоцист в эктоплазме туфельки развиваются новые.На одной стороне, приблизительно по середине тела (рис. 85, 5), у туфельки имеется довольно глубокая впадина. Это ротовая впадина, или перистом. По стенкам перистома, так же как и по поверхности тела, расположены реснички. Они развиты здесь гораздо более мощно, чем на всей остальной поверхности тела. Эти тесно расположенные реснички собраны в две группы. Функция этих особо дифференцированных ресничек связана не с движением, а с питанием (рис. 87).
Перед нами пример отрицательного таксиса на химическое воздействие (отрицательный хемотаксис). Можно наблюдать у туфельки и положительный хемотаксис. Если, например, каплю воды, в которой плавают инфузории, прикрыть покровным стеклышком и подпустить под него пузырек углекислого газа (С02), то большая часть инфузорий направится к этому пузырьку и расположится вокруг него кольцом.Очень наглядно явление таксиса проявляется у туфелек под влиянием электрического тока. Если через жидкость, в которой плавают туфельки, пропустить слабый электрический ток, то можно наблюдать следующую картину: все инфузории ориентируют свою продольную ось параллельно линии тока, а затем, как по команде, двинутся в направлении катода, в области которого и образуют густое скопление. Движение инфузорий, определяемое направлением электрического тока, носит название гальванотаксиса. Различные таксисы у инфузорий могут быть обнаружены под влиянием самых разнообразных факторов внешней среды.Все цитоплазматическое тело инфузории отчетливо распадается на 2 слоя: наружный — более светлый (эктоплазма) и внутренний —' более темный и зернистый (эндоплазма). Самый поверхностный слой эктоплазмы образует наружную очень тонкую и вместе с тем прочную и эластичную оболочку — пелликулу, которая играет важную роль в сохранении постоянства формы тела инфузории.В наружном слое (в эктоплазме) тела живой туфельки хорошо видны многочисленные коротенькие палочки, расположенные перпендикулярно к поверхности (рис. 85, 7). Эти образования носят название трихоцисты. Функция их очень интересна и связана с защитой простейшего. При механическом, химическом или каком-либо ином сильном раздражении трихоцисты с силой выбрасываются наружу, превращаясь в тонкие длинные нити, которые поражают хищника, нападающего на туфельку. Трихоцисты представляют собой мощную защиту. Они располагаются закономерно между ресничками, так что число трихоцист приблизительно соответствует числу ресничек. На месте использованных («выстреленных») трихоцист в эктоплазме туфельки развиваются новые.На одной стороне, приблизительно по середине тела (рис. 85, 5), у туфельки имеется довольно глубокая впадина. Это ротовая впадина, или перистом. По стенкам перистома, так же как и по поверхности тела, расположены реснички. Они развиты здесь гораздо более мощно, чем на всей остальной поверхности тела. Эти тесно расположенные реснички собраны в две группы. Функция этих особо дифференцированных ресничек связана не с движением, а с питанием (рис. 87).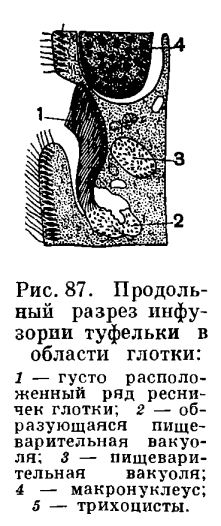 Как и чем питаются туфельки, как осуществляется у них пищеварение?Туфельки относятся к числу инфузорий, основную пищу которых составляют бактерии. Наряду с бактериями они могут заглатывать и любые другие взвешенные в воде частицы независимо от их питательности. Околоротовые реснички создают непрерывный ток воды со взвешенными в ней частицами в направлении ротового отверстия, которое расположено в глубине перистома. Мелкие пищевые частицы (чаще всего бактерии) проникают через рот в небольшую трубковидную глотку и скапливаются на дне ее, на границе с эндоплазмой. Ротовое отверстие всегда открыто. Пожалуй, не будет ошибкой сказать, что инфузория туфелька — одно из самых прожорливых животных: она непрерывно питается. Этот процесс прерывается только в определенные моменты жизни, связанные с размножением и половым процессом.
Как и чем питаются туфельки, как осуществляется у них пищеварение?Туфельки относятся к числу инфузорий, основную пищу которых составляют бактерии. Наряду с бактериями они могут заглатывать и любые другие взвешенные в воде частицы независимо от их питательности. Околоротовые реснички создают непрерывный ток воды со взвешенными в ней частицами в направлении ротового отверстия, которое расположено в глубине перистома. Мелкие пищевые частицы (чаще всего бактерии) проникают через рот в небольшую трубковидную глотку и скапливаются на дне ее, на границе с эндоплазмой. Ротовое отверстие всегда открыто. Пожалуй, не будет ошибкой сказать, что инфузория туфелька — одно из самых прожорливых животных: она непрерывно питается. Этот процесс прерывается только в определенные моменты жизни, связанные с размножением и половым процессом. Скопившийся на дне глотки пищевой комочек в дальнейшем отрывается от дна глотки и вместе с небольшим количеством жидкости поступает в эндоплазму, образуя пищеварительную вакуолю. Последняя не остается на месте своего образования, а, попадая в токи эндоплазмы, проделывает в теле туфельки довольно сложный и закономерный путь, называемый циклозом пищеварительной вакуоли (рис. 88). Во время этого довольно длительного (при комнатной температуре занимающего около часа) путешествия пищеварительной вакуоли внутри ее происходит ряд изменений, связанных с перевариванием находящейся в ней пищи.Здесь, так же как у амеб и некоторых жгутиконосцев, происходит типичное внутриклеточное пищеварение. Из окружающей пищеварительную вакуолю эндоплазмы в нее поступают пищеварительные ферменты, которые воздействуют на пищевые частицы. Продукты переваривания пищи всасываются через стгнку пищеварительной вакуоли в эндоплазму.По ходу циклоза пищеварительной вакуоли в ней сменяется несколько фаз пищеварения. В первые моменты после образования вакуоли заполняющая ее жидкость мало отличается от жидкости окружающей среды. Вскоре начинается поступление из эндоплазмы в вакуолю пищеварительных ферментов и реакция среды внутри нее становится резко кислой. Это легко обнаружить, добавляя к пище какой-либо индикатор, цвет которого меняется в зависимости от реакции (кислой, нейтральной или щелочной) среды. В этой кислой среде проходят первые фазы пищеварения. Затем картина меняется и реакция внутри пищеварительных вакуолей становится слабощелочной. В этих условиях и протекают дальнейшие этапы внутриклеточного пищеварения. Кислая фаза обычно более короткая, чем щелочная; она длится примерно 1/6—1/4 часть всего срока пребывания пищеварительной вакуоли в теле инфузории. Однако соотношение кислой и щелочной фаз может варьировать в довольно широких пределах в зависимости от характера пищи.Путь пищеварительной вакуоли в эндоплазме заканчивается тем, что она приближается к поверхности тела и через пелликулу содержимое ее, состоящее из жидкости и непереваренных остатков пищи, выбрасывается наружу — происходит дефекация. Этот процесс, в отличие от амеб, у которых дефекация может происходить в любом месте, у туфелек, как и у других инфузорий, строго приурочен к определенному участку тела, расположенному на брюшной стороне (брюшной условно называют ту поверхность животного, на которой помещается околоротовое углубление), примерно посередине между перистомом и задним концом тела.Таким образом, внутриклеточное пищеварение представляет собой сложный процесс, слагающийся из нескольких последовательно сменяющих друг друга фаз.
Скопившийся на дне глотки пищевой комочек в дальнейшем отрывается от дна глотки и вместе с небольшим количеством жидкости поступает в эндоплазму, образуя пищеварительную вакуолю. Последняя не остается на месте своего образования, а, попадая в токи эндоплазмы, проделывает в теле туфельки довольно сложный и закономерный путь, называемый циклозом пищеварительной вакуоли (рис. 88). Во время этого довольно длительного (при комнатной температуре занимающего около часа) путешествия пищеварительной вакуоли внутри ее происходит ряд изменений, связанных с перевариванием находящейся в ней пищи.Здесь, так же как у амеб и некоторых жгутиконосцев, происходит типичное внутриклеточное пищеварение. Из окружающей пищеварительную вакуолю эндоплазмы в нее поступают пищеварительные ферменты, которые воздействуют на пищевые частицы. Продукты переваривания пищи всасываются через стгнку пищеварительной вакуоли в эндоплазму.По ходу циклоза пищеварительной вакуоли в ней сменяется несколько фаз пищеварения. В первые моменты после образования вакуоли заполняющая ее жидкость мало отличается от жидкости окружающей среды. Вскоре начинается поступление из эндоплазмы в вакуолю пищеварительных ферментов и реакция среды внутри нее становится резко кислой. Это легко обнаружить, добавляя к пище какой-либо индикатор, цвет которого меняется в зависимости от реакции (кислой, нейтральной или щелочной) среды. В этой кислой среде проходят первые фазы пищеварения. Затем картина меняется и реакция внутри пищеварительных вакуолей становится слабощелочной. В этих условиях и протекают дальнейшие этапы внутриклеточного пищеварения. Кислая фаза обычно более короткая, чем щелочная; она длится примерно 1/6—1/4 часть всего срока пребывания пищеварительной вакуоли в теле инфузории. Однако соотношение кислой и щелочной фаз может варьировать в довольно широких пределах в зависимости от характера пищи.Путь пищеварительной вакуоли в эндоплазме заканчивается тем, что она приближается к поверхности тела и через пелликулу содержимое ее, состоящее из жидкости и непереваренных остатков пищи, выбрасывается наружу — происходит дефекация. Этот процесс, в отличие от амеб, у которых дефекация может происходить в любом месте, у туфелек, как и у других инфузорий, строго приурочен к определенному участку тела, расположенному на брюшной стороне (брюшной условно называют ту поверхность животного, на которой помещается околоротовое углубление), примерно посередине между перистомом и задним концом тела.Таким образом, внутриклеточное пищеварение представляет собой сложный процесс, слагающийся из нескольких последовательно сменяющих друг друга фаз. Кроме пищеварительных вакуолей, в теле инфузории туфельки имеются выполняющие также очень важные жизненные функции сократительные вакуоли. У туфельки их две, они расположены в передней и задней трети тела. Каждая из вакуолей состоит из центрального резервуара и приводящих каналов (5—7), которые расположены радиально вокруг центрального резервуара. Цикл работы сократительной вакуоли начинается с того, что приводящие каналы заполняются жидкостью и становятся хорошо видимыми (рис. 89). Затем жидкое содержимое их изливается в центральный резервуар, сами каналы после опорожнения становятся на некоторое время невидимыми. Последний этап цикла работы сократительной вакуоли заключается в том, что жидкость из центрального резервуара изливается через особую пору в пелликуле наружу. После этого центральный резервуар на короткий срок перестает быть видимым. В это время приводящие каналы вновь начинают заполняться жидкостью и весь цикл начинается сначала. Обычно передняя и задняя сократительные вакуоли работают последовательно, как бы по очереди. Каков темп пульсации вакуолей? Какое количество жидкости выводится ими наружу? Частота сокращения сократительной вакуоли в большой степени зависит от условий внешней среды, и особенно от температуры и осмотического давления. При комнатной температуре у туфельки сократительная вакуоля проделывает весь цикл пульсации за 10—15 секунд. У морских и паразитических инфузорий темп пульсации сократительных вакуолей обычно значительно ниже, чем у пресноводных.Подсчеты показывают, что примерно за 30—45 минут у туфельки через сократительные вакуоли выводится объем жидкости, равный объему тела инфузории. Таким образом, благодаря деятельности сократительных вакуолей через тело инфузории осуществляется непрерывный ток воды, поступающей снаружи через ротовое отверстие (вместе с пищеварительными вакуолями), а также осмотически непосредственно через пелликулу. Сократительные вакуоли играют важную роль в регулировании тока воды, проходящего через тело инфузории, в регулировании осмотического давления. Этот процесс здесь протекает в принципе так же, как у амеб, только структура сократительной вакуоли намного сложнее.В течение долгих лет среди ученых, занимающихся изучением простейших, шел спор по вопросу о том, имеются ли в цитоплазме какие-нибудь структуры, связанные с появлением сократительной вакуоли, или же она образуется всякий раз заново. На живой инфузории никаких особых структур, которые предшествовали бы ее образованию, наблюдать не удается. После того как произойдет сокращение вакуоли — систола, в цитоплазме на месте бывшей вакуоли не видно решительно никаких структур. Затем заново появляются прозрачный пузырек или приводящие каналы, которые начинают увеличиваться в размерах. Однако никакой связи вновь возникающей вакуоли с существовавшей ранее не обнаруживается. Создается впечатление, что преемственности между следующими друг за другом циклами сократительной вакуоли нет и всякая новая сократительная вакуоля образуется в цитоплазме заново. Однако специальные методы исследования показали, что на самом деле это не так. Применение электронной микроскопии, дающей очень большое увеличение (до 100 тыс. раз), убедительно показало, что у инфузории на том участке, где формируются сократительные вакуоли, имеется особо дифференцированная цитоплазма, состоящая из переплетения тончайших трубочек. Таким образом, оказалось, что сократительная вакуоля возникает в цитоплазме не на «пустом месте», а на основе предшествующего особого органоида клетки, функцией которого является формирование сократительной вакуоли.Как и у всех простейших, у инфузорий имеется клеточное ядро. Однако по строению ядерного аппарата инфузории резко отличаются от всех других групп простейших.Ядерный аппарат инфузорий характеризуется своим дуализмом. Это означает, что у инфузорий имеется два разных типа ядер — большие ядра, или макронуклеусы, и малые ядра, или микронуклеусы. Посмотрим, какое строение имеет ядерный аппарат у инфузории туфельки (рис. 85).В центре тела инфузории (на уровне перистома) помещается большое массивное ядро яйцевидной или бобовидной формы. Это макронуклеус. В тесном соседстве с ним расположено второе ядро во много раз меньших размеров, обычно довольно тесно прилежащее к макронуклеусу. Это микронуклеус. Различие между этими двумя ядрами не сводится только к размерам, оно более значительно, глубоко затрагивает их структуру.Макронуклеус по сравнению с микронуклеусом гораздо богаче особым ядерным веществом (хроматином, или, точнее, дезоксирибонуклеиновой кислотой, сокращенно ДНК), входящим в состав хромосом.Исследования последних лет показали, что макронуклеус обладает в несколько десятков (а у некоторых инфузорий и сотен) раз большим числом хромосом, чем микронуклеусы. Макронуклеус является совсем своеобразным типом многохромосомных (полиплоидных) ядер. Таким образом, различие между микро- и макронуклеусами затрагивает их хромосомный состав, отчего и зависит большее или меньшее богатство их ядерным веществом — хроматином.У одного из наиболее обычных видов инфузории — туфельки (Paramecium caudatum) — имеется один макронуклеус (сокращенно Ма) и один микронуклеус (сокращенно Mi). Такая структура ядерного аппарата свойственна многим инфузориям. У других может быть по нескольку Ма и Mi. Но характерная особенность всех инфузорий — дифференцировка ядер на две качественно различные группы, на Ма и Mi, или, другими словами, явление ядерного дуализма.
Кроме пищеварительных вакуолей, в теле инфузории туфельки имеются выполняющие также очень важные жизненные функции сократительные вакуоли. У туфельки их две, они расположены в передней и задней трети тела. Каждая из вакуолей состоит из центрального резервуара и приводящих каналов (5—7), которые расположены радиально вокруг центрального резервуара. Цикл работы сократительной вакуоли начинается с того, что приводящие каналы заполняются жидкостью и становятся хорошо видимыми (рис. 89). Затем жидкое содержимое их изливается в центральный резервуар, сами каналы после опорожнения становятся на некоторое время невидимыми. Последний этап цикла работы сократительной вакуоли заключается в том, что жидкость из центрального резервуара изливается через особую пору в пелликуле наружу. После этого центральный резервуар на короткий срок перестает быть видимым. В это время приводящие каналы вновь начинают заполняться жидкостью и весь цикл начинается сначала. Обычно передняя и задняя сократительные вакуоли работают последовательно, как бы по очереди. Каков темп пульсации вакуолей? Какое количество жидкости выводится ими наружу? Частота сокращения сократительной вакуоли в большой степени зависит от условий внешней среды, и особенно от температуры и осмотического давления. При комнатной температуре у туфельки сократительная вакуоля проделывает весь цикл пульсации за 10—15 секунд. У морских и паразитических инфузорий темп пульсации сократительных вакуолей обычно значительно ниже, чем у пресноводных.Подсчеты показывают, что примерно за 30—45 минут у туфельки через сократительные вакуоли выводится объем жидкости, равный объему тела инфузории. Таким образом, благодаря деятельности сократительных вакуолей через тело инфузории осуществляется непрерывный ток воды, поступающей снаружи через ротовое отверстие (вместе с пищеварительными вакуолями), а также осмотически непосредственно через пелликулу. Сократительные вакуоли играют важную роль в регулировании тока воды, проходящего через тело инфузории, в регулировании осмотического давления. Этот процесс здесь протекает в принципе так же, как у амеб, только структура сократительной вакуоли намного сложнее.В течение долгих лет среди ученых, занимающихся изучением простейших, шел спор по вопросу о том, имеются ли в цитоплазме какие-нибудь структуры, связанные с появлением сократительной вакуоли, или же она образуется всякий раз заново. На живой инфузории никаких особых структур, которые предшествовали бы ее образованию, наблюдать не удается. После того как произойдет сокращение вакуоли — систола, в цитоплазме на месте бывшей вакуоли не видно решительно никаких структур. Затем заново появляются прозрачный пузырек или приводящие каналы, которые начинают увеличиваться в размерах. Однако никакой связи вновь возникающей вакуоли с существовавшей ранее не обнаруживается. Создается впечатление, что преемственности между следующими друг за другом циклами сократительной вакуоли нет и всякая новая сократительная вакуоля образуется в цитоплазме заново. Однако специальные методы исследования показали, что на самом деле это не так. Применение электронной микроскопии, дающей очень большое увеличение (до 100 тыс. раз), убедительно показало, что у инфузории на том участке, где формируются сократительные вакуоли, имеется особо дифференцированная цитоплазма, состоящая из переплетения тончайших трубочек. Таким образом, оказалось, что сократительная вакуоля возникает в цитоплазме не на «пустом месте», а на основе предшествующего особого органоида клетки, функцией которого является формирование сократительной вакуоли.Как и у всех простейших, у инфузорий имеется клеточное ядро. Однако по строению ядерного аппарата инфузории резко отличаются от всех других групп простейших.Ядерный аппарат инфузорий характеризуется своим дуализмом. Это означает, что у инфузорий имеется два разных типа ядер — большие ядра, или макронуклеусы, и малые ядра, или микронуклеусы. Посмотрим, какое строение имеет ядерный аппарат у инфузории туфельки (рис. 85).В центре тела инфузории (на уровне перистома) помещается большое массивное ядро яйцевидной или бобовидной формы. Это макронуклеус. В тесном соседстве с ним расположено второе ядро во много раз меньших размеров, обычно довольно тесно прилежащее к макронуклеусу. Это микронуклеус. Различие между этими двумя ядрами не сводится только к размерам, оно более значительно, глубоко затрагивает их структуру.Макронуклеус по сравнению с микронуклеусом гораздо богаче особым ядерным веществом (хроматином, или, точнее, дезоксирибонуклеиновой кислотой, сокращенно ДНК), входящим в состав хромосом.Исследования последних лет показали, что макронуклеус обладает в несколько десятков (а у некоторых инфузорий и сотен) раз большим числом хромосом, чем микронуклеусы. Макронуклеус является совсем своеобразным типом многохромосомных (полиплоидных) ядер. Таким образом, различие между микро- и макронуклеусами затрагивает их хромосомный состав, отчего и зависит большее или меньшее богатство их ядерным веществом — хроматином.У одного из наиболее обычных видов инфузории — туфельки (Paramecium caudatum) — имеется один макронуклеус (сокращенно Ма) и один микронуклеус (сокращенно Mi). Такая структура ядерного аппарата свойственна многим инфузориям. У других может быть по нескольку Ма и Mi. Но характерная особенность всех инфузорий — дифференцировка ядер на две качественно различные группы, на Ма и Mi, или, другими словами, явление ядерного дуализма. Как размножаются инфузории? Обратимся в качестве примера опять к инфузории туфельке. Если отсадить в небольшой сосуд (микроаквариум) один-единственный экземпляр туфельки, то уже через сутки там окажется две, а нередко и четыре инфузории. Как это происходит? После некоторого периода активного плавания и питания инфузория несколько вытягивается в длину. Затем точно по середине тела появляется все углубляющаяся поперечная перетяжка (рис. 90). В конце концов инфузория как бы перешнуровывается пополам и из одной особи получаются две, первоначально несколько меньших размеров, чем материнская особь. Весь процесс деления занимает при комнатной температуре около часа. Изучение внутренних процессов показывает, что еще до того, как появляется поперечная перетяжка, начинается процесс деления ядерного аппарата. Первым делится Mi и лишь после него — Ма. Мы не будем здесь останавливаться на детальном рассмотрении процессов ядерного деления и лишь отметим, что Mi делится путем митоза, тогда как деление Ма по внешности напоминает прямое деление ядра — амитоз. Этот бесполый процесс размножения инфузории туфельки, как мы видим, сходен с бесполым размножением амеб и жгутиконосцев. В отличие от них инфузории в процессе бесполого размножения делятся всегда поперек, тогда как у жгутиконосцев плоскость деления параллельна продольной оси тела.Во время деления происходит глубокая внутренняя перестройка тела инфузории. Образуется два новых перистома, две глотки и два ротовых отверстия. К этому же времени приурочено деление базальных ядер ресничек, за счет которых образуются новые реснички. Если бы при размножении число ресничек не возрастало, то в результате каждого деления дочерние особи получили бы примерно половину числа ресничек материнской особи, что привело бы к полному «облысению» инфузорий. На самом деле этого не происходит.
Как размножаются инфузории? Обратимся в качестве примера опять к инфузории туфельке. Если отсадить в небольшой сосуд (микроаквариум) один-единственный экземпляр туфельки, то уже через сутки там окажется две, а нередко и четыре инфузории. Как это происходит? После некоторого периода активного плавания и питания инфузория несколько вытягивается в длину. Затем точно по середине тела появляется все углубляющаяся поперечная перетяжка (рис. 90). В конце концов инфузория как бы перешнуровывается пополам и из одной особи получаются две, первоначально несколько меньших размеров, чем материнская особь. Весь процесс деления занимает при комнатной температуре около часа. Изучение внутренних процессов показывает, что еще до того, как появляется поперечная перетяжка, начинается процесс деления ядерного аппарата. Первым делится Mi и лишь после него — Ма. Мы не будем здесь останавливаться на детальном рассмотрении процессов ядерного деления и лишь отметим, что Mi делится путем митоза, тогда как деление Ма по внешности напоминает прямое деление ядра — амитоз. Этот бесполый процесс размножения инфузории туфельки, как мы видим, сходен с бесполым размножением амеб и жгутиконосцев. В отличие от них инфузории в процессе бесполого размножения делятся всегда поперек, тогда как у жгутиконосцев плоскость деления параллельна продольной оси тела.Во время деления происходит глубокая внутренняя перестройка тела инфузории. Образуется два новых перистома, две глотки и два ротовых отверстия. К этому же времени приурочено деление базальных ядер ресничек, за счет которых образуются новые реснички. Если бы при размножении число ресничек не возрастало, то в результате каждого деления дочерние особи получили бы примерно половину числа ресничек материнской особи, что привело бы к полному «облысению» инфузорий. На самом деле этого не происходит.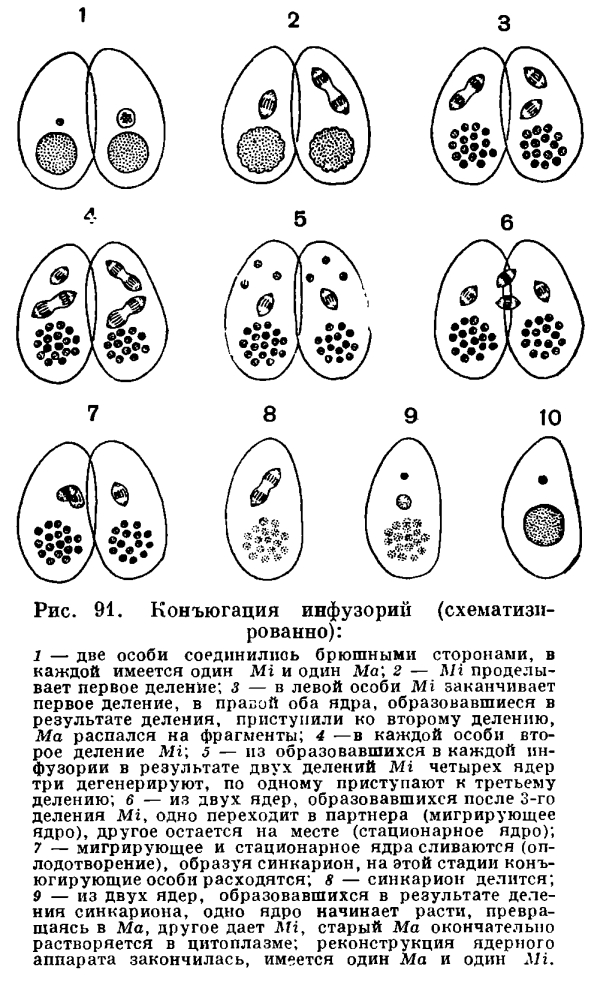 Время от времени у большинства инфузорий, в том числе и у туфелек, наблюдается особая и чрезвычайно своеобразная форма полового процесса, которая получила название конъюгации. Мы не будем здесь детально разбирать все те сложные ядерные изменения, которые сопровождают этот процесс, а отметим только самое главное. Конъюгация протекает следующим образом (рис. 91), Две инфузории сближаются, тесно прикладываются друг к другу брюшными сторонами и в таком виде плавают довольно длительное время вместе (у туфельки примерно в течение 12 часов при комнатной температуре). Затем конъюганты расходятся. Что же происходит в теле инфузории во время конъюгации? Сущность этих процессов сводится к следующему (рис. 91). Большое ядро (макронуклеус) разрушается и постепенно растворяется в цитоплазме. Микронуклеусы сначала делятся, часть ядер, образующихся в результате деления, разрушается (см. рис. 91). В каждом из конъюгантов сохраняется по два ядра. Одно из этих ядер остается на месте в той особи, в которой оно образовалось (стационарное ядро), другое же активно перемещается в партнера по конъюгации (мигрирующее ядро) и сливается с его стационарным ядром. Таким образом, в каждом из конъюгантов на этой стадии имеется по одному ядру, образовавшемуся в результате слияния стационарного и мигрирующего ядер. Это сложное ядро получает название синкарион. Образование синкариона представляет собой не что иное, как процесс оплодотворения. И у многоклеточных существенным моментом оплодотворения является слияние ядер половых клеток. У инфузорий половые клетки не образуются, имеются лишь половые ядра, которые и сливаются между собой. Таким образом происходит взаимное перекрестное оплодотворение.Вскоре после образования синкариона конъюганты расходятся. По строению своего ядерного аппарата они на этой стадии еще очень существенно отличаются от обычных так называемых нейтральных (не конъюгирующих) инфузорий, так как у них имеется лишь по одному ядру. В дальнейшем за счет синкариона происходит восстановление нормального ядерного аппарата. Синкарион делится (один или несколько раз). Часть продуктов этого деления путем сложных преобразований, связанных с увеличением числа хромосом и обогащением хроматином, превращается в макронуклеусы. Другие сохраняют структуру, характерную для микронуклеусов. Таким путем восстанавливается характерный и типичный для инфузорий ядерный аппарат, после чего инфузории приступают к бесполому размножению путем деления.Таким образом, процесс конъюгации включает в себя два существенно важных биологических момента: оплодотворение и восстановление нового макронуклеуса за счет синкариона.В чем заключается биологическое значение конъюгации, какую роль играет она в жизни инфузорий? Мы не можем назвать ее размножением, ибо при этом увеличения числа особей не происходит. Поставленные выше вопросы послужили материалом для многочисленных экспериментальных исследований, осуществлявшихся во многих странах. Основной результат этих исследований сводится к следующему. Во-первых, конъюгация, как и всякий другой половой процесс, при котором происходит объединение в одном организме двух наследственных начал (отцовского и материнского), ведет к повышению наследственной изменчивости, наследственного многообразия. Повышение наследственной изменчивости увеличивает приспособительные возможности организма к условиям окружающей среды. Второй биологически важной стороной конъюгации является развитие нового макронуклеуса за счет продуктов деления синкариона и одновременно с этим разрушение старого. Экспериментальные данные показывают, что именно макронуклеус играет исключительно важную роль в жизни инфузорий. Им контролируются все основные жизненные процессы и определяется важнейший из них — образование (синтез) белка, составляющего основную часть протоплазмы живой клетки. При длительном бесполом размножении путем деления происходит как бы своеобразный процесс «старения» макронуклеуса, а вместе с тем и всей клетки: снижается активность процесса обмена веществ, снижается темп деления. После конъюгации (в процессе которой, как мы видели, старый макронуклеус разрушается) происходит восстановление уровня обмена веществ и темпа деления. Поскольку при конъюгации происходит процесс оплодотворения, который у большинства других организмов связан с размножением и появлением нового поколения, у инфузорий особь, образовавшуюся после конъюгации, тоже можно рассматривать как новое половое поколение, которое возникает здесь как бы за счет «омолаживания» старого.На примере инфузории туфельки мы познакомились с типичным представителем обширного класса инфузорий. Однако класс этот характерен чрезвычайным разнообразием видов как по строению, так и по образу жизни. Познакомимся ближе с некоторыми наиболее характерными и интересными формами.У инфузории туфельки реснички равномерно покрывают всю поверхность тела. Это характерная черта строения отряда равноресничных инфузорий (Holotricha). Многим инфузориям свойствен иной характер развития ресничного покрова. Дело в том, что реснички инфузорий способны, соединяясь вместе, образовывать более сложные комплексы. Например, нередко наблюдается, что расположенные в один или два ряда близко друг к другу реснички соединяются (слипаются) вместе, образуя пластинку, которая, так же как и реснички, способна к биению. Такие пластинчатые сократимые образования получили название мембранелл (если они короткие) или мембран (если они длинные). В других случаях соединяются вместе реснички, расположенные тесным пучком. Эти образования — цирры — напоминают кисточку, отдельные волоски которой слеплены друг с другом. Разного рода сложные ресничные образования характерны для многих инфузорий. Очень часто ресничный покров развивается не равномерно, а лишь на некоторых участках тела.ИНФУЗОРИЯ ТРУБАЧ (STENTOR POLYMORPHIC)В пресных водах очень часто встречаются виды крупных красивых инфузорий, относящихся к роду трубачей (Stentor). Это название вполне соответствует форме тела этих животных, которая действительно напоминает трубу (рис. 92), широко раскрывшуюся на одном конце. При первом же знакомстве с живыми трубачами можно заметить одну особенность, которая не свойственна туфельке. При малейшем раздражении, в том числе механическом (например, постукивание карандашом по стеклу, где имеется капля воды с трубачами), тело их резко и очень быстро (в долю секунды) сокращается, принимая почти правильную шарообразную форму. Затем довольно медленно (время измеряется секундами) трубач расправляется, принимая характерную для него форму. Эта способность трубача быстро сокращаться обусловлена наличием особых мускульных волоконец, расположенных вдоль тела и в эктоплазме. Таким образом, и в одноклеточном организме может развиться мышечная система.
Время от времени у большинства инфузорий, в том числе и у туфелек, наблюдается особая и чрезвычайно своеобразная форма полового процесса, которая получила название конъюгации. Мы не будем здесь детально разбирать все те сложные ядерные изменения, которые сопровождают этот процесс, а отметим только самое главное. Конъюгация протекает следующим образом (рис. 91), Две инфузории сближаются, тесно прикладываются друг к другу брюшными сторонами и в таком виде плавают довольно длительное время вместе (у туфельки примерно в течение 12 часов при комнатной температуре). Затем конъюганты расходятся. Что же происходит в теле инфузории во время конъюгации? Сущность этих процессов сводится к следующему (рис. 91). Большое ядро (макронуклеус) разрушается и постепенно растворяется в цитоплазме. Микронуклеусы сначала делятся, часть ядер, образующихся в результате деления, разрушается (см. рис. 91). В каждом из конъюгантов сохраняется по два ядра. Одно из этих ядер остается на месте в той особи, в которой оно образовалось (стационарное ядро), другое же активно перемещается в партнера по конъюгации (мигрирующее ядро) и сливается с его стационарным ядром. Таким образом, в каждом из конъюгантов на этой стадии имеется по одному ядру, образовавшемуся в результате слияния стационарного и мигрирующего ядер. Это сложное ядро получает название синкарион. Образование синкариона представляет собой не что иное, как процесс оплодотворения. И у многоклеточных существенным моментом оплодотворения является слияние ядер половых клеток. У инфузорий половые клетки не образуются, имеются лишь половые ядра, которые и сливаются между собой. Таким образом происходит взаимное перекрестное оплодотворение.Вскоре после образования синкариона конъюганты расходятся. По строению своего ядерного аппарата они на этой стадии еще очень существенно отличаются от обычных так называемых нейтральных (не конъюгирующих) инфузорий, так как у них имеется лишь по одному ядру. В дальнейшем за счет синкариона происходит восстановление нормального ядерного аппарата. Синкарион делится (один или несколько раз). Часть продуктов этого деления путем сложных преобразований, связанных с увеличением числа хромосом и обогащением хроматином, превращается в макронуклеусы. Другие сохраняют структуру, характерную для микронуклеусов. Таким путем восстанавливается характерный и типичный для инфузорий ядерный аппарат, после чего инфузории приступают к бесполому размножению путем деления.Таким образом, процесс конъюгации включает в себя два существенно важных биологических момента: оплодотворение и восстановление нового макронуклеуса за счет синкариона.В чем заключается биологическое значение конъюгации, какую роль играет она в жизни инфузорий? Мы не можем назвать ее размножением, ибо при этом увеличения числа особей не происходит. Поставленные выше вопросы послужили материалом для многочисленных экспериментальных исследований, осуществлявшихся во многих странах. Основной результат этих исследований сводится к следующему. Во-первых, конъюгация, как и всякий другой половой процесс, при котором происходит объединение в одном организме двух наследственных начал (отцовского и материнского), ведет к повышению наследственной изменчивости, наследственного многообразия. Повышение наследственной изменчивости увеличивает приспособительные возможности организма к условиям окружающей среды. Второй биологически важной стороной конъюгации является развитие нового макронуклеуса за счет продуктов деления синкариона и одновременно с этим разрушение старого. Экспериментальные данные показывают, что именно макронуклеус играет исключительно важную роль в жизни инфузорий. Им контролируются все основные жизненные процессы и определяется важнейший из них — образование (синтез) белка, составляющего основную часть протоплазмы живой клетки. При длительном бесполом размножении путем деления происходит как бы своеобразный процесс «старения» макронуклеуса, а вместе с тем и всей клетки: снижается активность процесса обмена веществ, снижается темп деления. После конъюгации (в процессе которой, как мы видели, старый макронуклеус разрушается) происходит восстановление уровня обмена веществ и темпа деления. Поскольку при конъюгации происходит процесс оплодотворения, который у большинства других организмов связан с размножением и появлением нового поколения, у инфузорий особь, образовавшуюся после конъюгации, тоже можно рассматривать как новое половое поколение, которое возникает здесь как бы за счет «омолаживания» старого.На примере инфузории туфельки мы познакомились с типичным представителем обширного класса инфузорий. Однако класс этот характерен чрезвычайным разнообразием видов как по строению, так и по образу жизни. Познакомимся ближе с некоторыми наиболее характерными и интересными формами.У инфузории туфельки реснички равномерно покрывают всю поверхность тела. Это характерная черта строения отряда равноресничных инфузорий (Holotricha). Многим инфузориям свойствен иной характер развития ресничного покрова. Дело в том, что реснички инфузорий способны, соединяясь вместе, образовывать более сложные комплексы. Например, нередко наблюдается, что расположенные в один или два ряда близко друг к другу реснички соединяются (слипаются) вместе, образуя пластинку, которая, так же как и реснички, способна к биению. Такие пластинчатые сократимые образования получили название мембранелл (если они короткие) или мембран (если они длинные). В других случаях соединяются вместе реснички, расположенные тесным пучком. Эти образования — цирры — напоминают кисточку, отдельные волоски которой слеплены друг с другом. Разного рода сложные ресничные образования характерны для многих инфузорий. Очень часто ресничный покров развивается не равномерно, а лишь на некоторых участках тела.ИНФУЗОРИЯ ТРУБАЧ (STENTOR POLYMORPHIC)В пресных водах очень часто встречаются виды крупных красивых инфузорий, относящихся к роду трубачей (Stentor). Это название вполне соответствует форме тела этих животных, которая действительно напоминает трубу (рис. 92), широко раскрывшуюся на одном конце. При первом же знакомстве с живыми трубачами можно заметить одну особенность, которая не свойственна туфельке. При малейшем раздражении, в том числе механическом (например, постукивание карандашом по стеклу, где имеется капля воды с трубачами), тело их резко и очень быстро (в долю секунды) сокращается, принимая почти правильную шарообразную форму. Затем довольно медленно (время измеряется секундами) трубач расправляется, принимая характерную для него форму. Эта способность трубача быстро сокращаться обусловлена наличием особых мускульных волоконец, расположенных вдоль тела и в эктоплазме. Таким образом, и в одноклеточном организме может развиться мышечная система.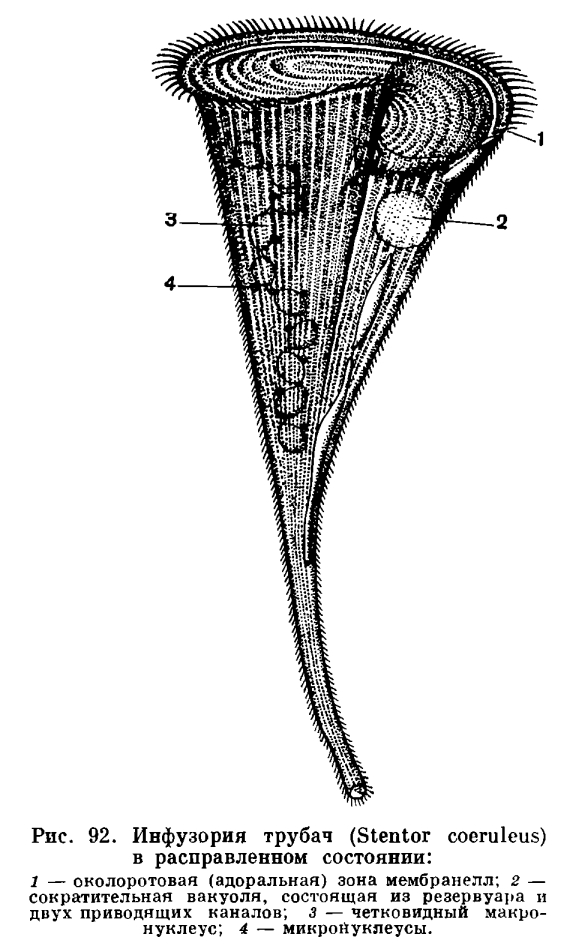 В роде трубачей имеются виды, некоторые из которых характеризуются довольно яркой окраской. Очень обычен в пресных водах голубой трубач (Stentor coeruleus), который окрашен в ярко-голубой цвет. Эта окраска трубача обусловлена тем, что в эктоплазме его расположены мельчайшие зерна синего пигмента.Другой вид трубача (Stentor polymorphus) нередко бывает окрашен в зеленый цвет. Причина этой окраски совсем иная. Зеленый цвет обусловлен тем, что в эндоплазме инфузории живут и размножаются мелкие одноклеточные зеленые водоросли, которые и придают телу трубача характерную окраску. Stentor polymorphus — типичный пример взаимно полезного сожительства — симбиоза. Трубач и водоросли находятся во взаимно симбиотических отношениях: трубач защищает водоросли, живущие в его теле, и снабжает их углекислотой, образующейся в результате дыхания; со своей стороны водоросли дают трубачу кислород, освобождающийся в процессе фотосинтеза. По-видимому, часть водорослей переваривается инфузорией, являясь пищей для трубача.Трубачи медленно плавают в воде широким концом вперед. Но они могут также временно прикрепляться к субстрату задним узким концом тела, на котором при этом образуется небольшая присоска.В теле трубача можно различить расширяющийся сзади наперед туловищный отдел и почти перпендикулярно к нему расположенное широкое околоротовое (перистомальное) поле. Это поле напоминает асимметричную плоскую воронку, с одного края которой имеется углубление — глотка, ведущая в эндоплазму инфузории. Тело трубача покрыто продольными рядами коротких ресничек. По краю перистомального поля по кругу располагается мощно развитая околоротовая (адоральная) зона мембранелл (рис. 92). Эта зона состоит из большого числа отдельных мерцательных пластинок, каждая из которых в свою очередь слагается из множества слипшихся друг с другом, расположенных двумя тесно сближенными рядами ресничек.В области ротового отверстия околоротовые мембранеллы заворачивают в сторону глотки, образуя левозакрученную спираль. Ток воды, вызываемый колебанием околоротовых мембранелл, направлен в сторону ротового отверстия (в глубину воронки, образуемой передним концом тела). Вместе с водой в глотку попадают и взвешенные в воде пищевые частицы. Пищевые объекты у трубача разнообразнее, чем у туфельки. Наряду с бактериями он поедает мелких простейших (например, жгутиковых), одноклеточные водоросли и т. п.У трубача хорошо развита сократительная вакуоля, имеющая своеобразное строение. Центральный резервуар расположен в передней трети тела, несколько ниже ротового отверстия. От него отходят два длинных приводящих канала. Один из них идет от резервуара к заднему концу тела, второй расположен в области перистомального поля параллельно околоротовой зоне мембранелл.Очень своеобразно устроен ядерный аппарат трубача. Макронуклеус здесь поделен на четки (их около 10), связанные друг с другом тонкими перемычками. Микронуклеусов несколько. Они очень мелкие и обычно тесно прилегают к четкам макронуклеуса.Инфузория трубач — излюбленный объект для экспериментальных исследований по регенерации. Многочисленными опытами была доказана высокая регенеративная способность трубачей. Инфузорию тонким скальпелем можно разрезать на множество частей, и каждая из них через короткое время (несколько часов, иногда сутки и более) превратится в пропорционально построенного, но маленького трубача, который затем в результате энергичного питания достигает типичного для данного вида размера. Для завершения восстановительных процессов в регенерирующем кусочке должен быть хотя бы один сегмент четковидного макронуклеуса.Трубач, как мы видели, обладает разными ресничками: с одной стороны короткими, покрывающими все тело, а с другой — околоротовой зоной мембранелл. В соответствии с этой характерной чертой строения отряд инфузорий, к которому относится трубач, получил название разноресничных инфузорий (Heterotricha).ИНФУЗОРИЯ БУРСАРИЯ (BURSARIA TRUNCATELLA)Второй интересный представитель разноресничных инфузорий — часто встречающаяся в пресных водах бурсария (Bursaria truncatella, рис. 93). Это гигант среди инфузорий: ее размеры могут достигать 2 мм, наиболее обычные величины — 0, 5—1, 0 мм. Бурсарию хорошо видно невооруженным глазом. В соответствии со своим названием бурсария имеет форму мешка, широко открытого на переднем конце (bursa — латинское слово, в переводе на русский язык означает «кошель», «мешок») и несколько расширенного на заднем конце. Все тело инфузории покрыто продольно идущими рядами коротких ресничек. Биение их обусловливает довольно медленное поступательное движение животного. Бурсария плывет как бы «переваливаясь» с бока на бок.
В роде трубачей имеются виды, некоторые из которых характеризуются довольно яркой окраской. Очень обычен в пресных водах голубой трубач (Stentor coeruleus), который окрашен в ярко-голубой цвет. Эта окраска трубача обусловлена тем, что в эктоплазме его расположены мельчайшие зерна синего пигмента.Другой вид трубача (Stentor polymorphus) нередко бывает окрашен в зеленый цвет. Причина этой окраски совсем иная. Зеленый цвет обусловлен тем, что в эндоплазме инфузории живут и размножаются мелкие одноклеточные зеленые водоросли, которые и придают телу трубача характерную окраску. Stentor polymorphus — типичный пример взаимно полезного сожительства — симбиоза. Трубач и водоросли находятся во взаимно симбиотических отношениях: трубач защищает водоросли, живущие в его теле, и снабжает их углекислотой, образующейся в результате дыхания; со своей стороны водоросли дают трубачу кислород, освобождающийся в процессе фотосинтеза. По-видимому, часть водорослей переваривается инфузорией, являясь пищей для трубача.Трубачи медленно плавают в воде широким концом вперед. Но они могут также временно прикрепляться к субстрату задним узким концом тела, на котором при этом образуется небольшая присоска.В теле трубача можно различить расширяющийся сзади наперед туловищный отдел и почти перпендикулярно к нему расположенное широкое околоротовое (перистомальное) поле. Это поле напоминает асимметричную плоскую воронку, с одного края которой имеется углубление — глотка, ведущая в эндоплазму инфузории. Тело трубача покрыто продольными рядами коротких ресничек. По краю перистомального поля по кругу располагается мощно развитая околоротовая (адоральная) зона мембранелл (рис. 92). Эта зона состоит из большого числа отдельных мерцательных пластинок, каждая из которых в свою очередь слагается из множества слипшихся друг с другом, расположенных двумя тесно сближенными рядами ресничек.В области ротового отверстия околоротовые мембранеллы заворачивают в сторону глотки, образуя левозакрученную спираль. Ток воды, вызываемый колебанием околоротовых мембранелл, направлен в сторону ротового отверстия (в глубину воронки, образуемой передним концом тела). Вместе с водой в глотку попадают и взвешенные в воде пищевые частицы. Пищевые объекты у трубача разнообразнее, чем у туфельки. Наряду с бактериями он поедает мелких простейших (например, жгутиковых), одноклеточные водоросли и т. п.У трубача хорошо развита сократительная вакуоля, имеющая своеобразное строение. Центральный резервуар расположен в передней трети тела, несколько ниже ротового отверстия. От него отходят два длинных приводящих канала. Один из них идет от резервуара к заднему концу тела, второй расположен в области перистомального поля параллельно околоротовой зоне мембранелл.Очень своеобразно устроен ядерный аппарат трубача. Макронуклеус здесь поделен на четки (их около 10), связанные друг с другом тонкими перемычками. Микронуклеусов несколько. Они очень мелкие и обычно тесно прилегают к четкам макронуклеуса.Инфузория трубач — излюбленный объект для экспериментальных исследований по регенерации. Многочисленными опытами была доказана высокая регенеративная способность трубачей. Инфузорию тонким скальпелем можно разрезать на множество частей, и каждая из них через короткое время (несколько часов, иногда сутки и более) превратится в пропорционально построенного, но маленького трубача, который затем в результате энергичного питания достигает типичного для данного вида размера. Для завершения восстановительных процессов в регенерирующем кусочке должен быть хотя бы один сегмент четковидного макронуклеуса.Трубач, как мы видели, обладает разными ресничками: с одной стороны короткими, покрывающими все тело, а с другой — околоротовой зоной мембранелл. В соответствии с этой характерной чертой строения отряд инфузорий, к которому относится трубач, получил название разноресничных инфузорий (Heterotricha).ИНФУЗОРИЯ БУРСАРИЯ (BURSARIA TRUNCATELLA)Второй интересный представитель разноресничных инфузорий — часто встречающаяся в пресных водах бурсария (Bursaria truncatella, рис. 93). Это гигант среди инфузорий: ее размеры могут достигать 2 мм, наиболее обычные величины — 0, 5—1, 0 мм. Бурсарию хорошо видно невооруженным глазом. В соответствии со своим названием бурсария имеет форму мешка, широко открытого на переднем конце (bursa — латинское слово, в переводе на русский язык означает «кошель», «мешок») и несколько расширенного на заднем конце. Все тело инфузории покрыто продольно идущими рядами коротких ресничек. Биение их обусловливает довольно медленное поступательное движение животного. Бурсария плывет как бы «переваливаясь» с бока на бок. От переднего конца в глубь тела (примерно на 2/3 его длины) вдается околоротовое углубление — перистом. С брюшной стороны оно сообщается с наружной средой узкой щелью, на спинной стороне полость перистома с наружной средой не сообщается. Если посмотреть на поперечный разрез верхней трети тела бурсарии (рис. 93, Б), то видно, что полость перистома занимает большую часть тела, тогда как цитоплазма окружает ее в виде ободка.На переднем конце тела, слева берет начало очень мощно развитая у бурсарии зона околоротовых (адоральных) мембранелл (рис. 93, 4). Она спускается в глубину полости перистома, заворачивая налево. Адоральная зона доходит до самой глубокой части перистома. Кроме околоротовых мембранелл, других ресничных образований в полости перистома нет, за исключением ресничной полоски, идущей по брюшной стороне полости перистома (рис. 93, 10). На внутренней задней стенке перистомальной полости почти по всей ее длине проходит узкая щель (рис. 93, 7), края которой обычно тесно прилегают друг к другу. Это ротовая щель. Ее края раздвигаются только в момент приема пищи.Бурсарии не обладают узкой пищевой специализацией, но в основном это хищники. При поступательном движении они наталкиваются на различных мелких животных. Благодаря работе мембранелл околоротовой зоны добыча с силой втягивается в обширную перистомальную полость, откуда уже не может выплыть наружу. Пищевые объекты прижимаются к спинной стенке перистомальной полости и через раздвигающуюся ротовую щель проникают в эндоплазму. Бурсарии очень прожорливы, они могут проглатывать довольно крупные объекты: например, излюбленной пищей их являются инфузории туфельки. Бурсария способна подряд проглотить 6—7 туфелек. В результате в эндоплазме бурсарии образуются очень крупные пищеварительные вакуоли.Ядерный аппарат бурсарии устроен довольно сложно. Они имеют один длинный колбасовидный макронуклеус и большое (примерно до 30) количество мелких микронуклеусов, беспорядочно разбросанных в эндоплазме инфузории.Бурсарии относятся к числу немногих видов пресноводных инфузорий, у которых отсутствует сократительная вакуоля. Как осуществляется у этой крупной инфузории осморегуляция, до сих пор остается не вполне ясным. Под эктоплазмой бурсарии в разных участках тела можно наблюдать различной формы и размеров пузырьки жидкости— вакуоли, которые меняют свой объем. По-видимому, эти неправильной формы вакуоли и соответствуют по своей функции сократительным вакуолям других инфузорий.
От переднего конца в глубь тела (примерно на 2/3 его длины) вдается околоротовое углубление — перистом. С брюшной стороны оно сообщается с наружной средой узкой щелью, на спинной стороне полость перистома с наружной средой не сообщается. Если посмотреть на поперечный разрез верхней трети тела бурсарии (рис. 93, Б), то видно, что полость перистома занимает большую часть тела, тогда как цитоплазма окружает ее в виде ободка.На переднем конце тела, слева берет начало очень мощно развитая у бурсарии зона околоротовых (адоральных) мембранелл (рис. 93, 4). Она спускается в глубину полости перистома, заворачивая налево. Адоральная зона доходит до самой глубокой части перистома. Кроме околоротовых мембранелл, других ресничных образований в полости перистома нет, за исключением ресничной полоски, идущей по брюшной стороне полости перистома (рис. 93, 10). На внутренней задней стенке перистомальной полости почти по всей ее длине проходит узкая щель (рис. 93, 7), края которой обычно тесно прилегают друг к другу. Это ротовая щель. Ее края раздвигаются только в момент приема пищи.Бурсарии не обладают узкой пищевой специализацией, но в основном это хищники. При поступательном движении они наталкиваются на различных мелких животных. Благодаря работе мембранелл околоротовой зоны добыча с силой втягивается в обширную перистомальную полость, откуда уже не может выплыть наружу. Пищевые объекты прижимаются к спинной стенке перистомальной полости и через раздвигающуюся ротовую щель проникают в эндоплазму. Бурсарии очень прожорливы, они могут проглатывать довольно крупные объекты: например, излюбленной пищей их являются инфузории туфельки. Бурсария способна подряд проглотить 6—7 туфелек. В результате в эндоплазме бурсарии образуются очень крупные пищеварительные вакуоли.Ядерный аппарат бурсарии устроен довольно сложно. Они имеют один длинный колбасовидный макронуклеус и большое (примерно до 30) количество мелких микронуклеусов, беспорядочно разбросанных в эндоплазме инфузории.Бурсарии относятся к числу немногих видов пресноводных инфузорий, у которых отсутствует сократительная вакуоля. Как осуществляется у этой крупной инфузории осморегуляция, до сих пор остается не вполне ясным. Под эктоплазмой бурсарии в разных участках тела можно наблюдать различной формы и размеров пузырьки жидкости— вакуоли, которые меняют свой объем. По-видимому, эти неправильной формы вакуоли и соответствуют по своей функции сократительным вакуолям других инфузорий.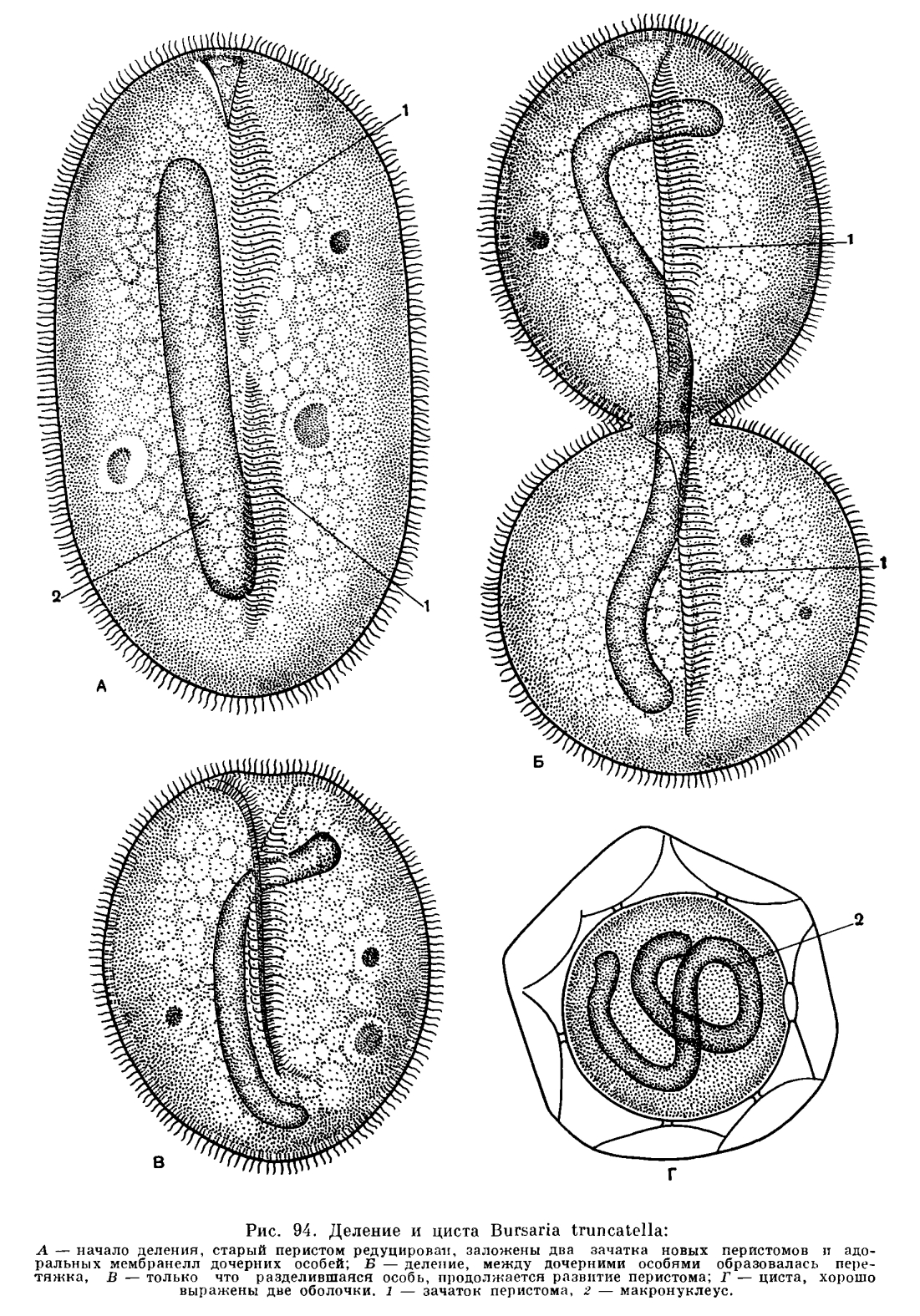 Интересно наблюдать за последовательными стадиями бесполого размножения бурсарии. На начальных стадиях деления происходит полная редукция всей полости перистома и околоротовой зоны мембранелл (рис. 94). Сохраняется лишь наружный ресничный покров. Инфузория приобретает форму яйца. После этого тело перешнуровывается поперечной бороздой на две половины. В каждой из образовавшихся дочерних инфузорий путем довольно сложных преобразований развивается типичный перистом и околоротовая зона мембранелл. Весь процесс деления бурсарии протекает быстро и занимает немногим больше часа.Очень легко наблюдать у бурсарии еще один важный жизненный процесс, наступление которого связано с неблагоприятными для инфузории условиями, — процесс образования цист (инцистирование). Это явление свойственно, например, амебам. Но оказывается, что и такие сложно организованные простейшие, как инфузории, способны переходить в недеятельное состояние. Если культуру, где живут бурсарии, вовремя не покормить или сильно охладить, то уже через несколько часов начнется массовое инцистирование. Протекает этот процесс следующим образом. Бурсарид, так же как перед делением, теряют перистом и околоротовую зону мембранелл. Затем они становятся совершенно шаровидными, после чего выделяют характерной формы двойную оболочку (рис. 94, Г).В состоянии цист бурсарии могут находиться месяцами. При наступлении благоприятных условий оболочка цисты лопается, бурсарии выходят из нее, развивают перистом и переходят к активной жизни.СТИЛОНИХИЯ (STYLONICHIA MYTILUS)Очень сложно и разнообразно дифференцированный ресничный аппарат имеют инфузории, относящиеся к отряду брюхоресничных (Hypotricha), многочисленные виды которых обитают как в пресной, так и в морской воде. Одним из наиболее обычных, часто встречающихся представителей этой интересной группы можно назвать стилонихию (Stylonichia mytilus). Это довольно крупная инфузория (длина до 0, 3 мм), живущая на дне пресноводных водоемов, на водной растительности (рис. 95). В отличие от туфельки, трубача и бурсарии у стилонихии отсутствует сплошной ресничный покров, а весь ресничный аппарат представлен ограниченным числом строго определенно расположенных ресничных образований.
Интересно наблюдать за последовательными стадиями бесполого размножения бурсарии. На начальных стадиях деления происходит полная редукция всей полости перистома и околоротовой зоны мембранелл (рис. 94). Сохраняется лишь наружный ресничный покров. Инфузория приобретает форму яйца. После этого тело перешнуровывается поперечной бороздой на две половины. В каждой из образовавшихся дочерних инфузорий путем довольно сложных преобразований развивается типичный перистом и околоротовая зона мембранелл. Весь процесс деления бурсарии протекает быстро и занимает немногим больше часа.Очень легко наблюдать у бурсарии еще один важный жизненный процесс, наступление которого связано с неблагоприятными для инфузории условиями, — процесс образования цист (инцистирование). Это явление свойственно, например, амебам. Но оказывается, что и такие сложно организованные простейшие, как инфузории, способны переходить в недеятельное состояние. Если культуру, где живут бурсарии, вовремя не покормить или сильно охладить, то уже через несколько часов начнется массовое инцистирование. Протекает этот процесс следующим образом. Бурсарид, так же как перед делением, теряют перистом и околоротовую зону мембранелл. Затем они становятся совершенно шаровидными, после чего выделяют характерной формы двойную оболочку (рис. 94, Г).В состоянии цист бурсарии могут находиться месяцами. При наступлении благоприятных условий оболочка цисты лопается, бурсарии выходят из нее, развивают перистом и переходят к активной жизни.СТИЛОНИХИЯ (STYLONICHIA MYTILUS)Очень сложно и разнообразно дифференцированный ресничный аппарат имеют инфузории, относящиеся к отряду брюхоресничных (Hypotricha), многочисленные виды которых обитают как в пресной, так и в морской воде. Одним из наиболее обычных, часто встречающихся представителей этой интересной группы можно назвать стилонихию (Stylonichia mytilus). Это довольно крупная инфузория (длина до 0, 3 мм), живущая на дне пресноводных водоемов, на водной растительности (рис. 95). В отличие от туфельки, трубача и бурсарии у стилонихии отсутствует сплошной ресничный покров, а весь ресничный аппарат представлен ограниченным числом строго определенно расположенных ресничных образований. Тело стилонихии (как и большинства других брюхоресничных инфузорий) сильно сплющено в спинно-брюшном направлении, и у нее ясно различимы спинная и брюшная стороны, передний и задний концы. Спереди тело несколько расширено, сзади — сужено. При рассматривании животного с брюшной стороны ясно видно, что в передней трети слева расположен сложно устроенный перистом и ротовое отверстие.На спинной стороне довольно редко расположены реснички, которые не способны к биению. Их скорее можно назвать тонкими эластичными щетинками. Они неподвижны и не имеют отношения к функции движения. Этим ресничкам приписывают обычно осязательную, чувствительную, функцию.Все ресничные образования, связанные с движением и захватом пищи, сосредоточены на брюшной стороне животного (рис. 95). Здесь имеется небольшое количество толстых пальцевидных образований, расположенных несколькими группами. Это брюшные цирры. Каждая из них — сложное ресничное образование, результат тесного соединения (слипания) многих десятков отдельных ресничек. Таким образом, цирры представляют собой как бы кисточки, отдельные волоски которых тесно сближены и соединены между собой.При помощи цирр животное довольно быстро передвигается, «бегает» по субстрату. Кроме «ползания» и «бегания» по субстрату стилонихия способна производить довольно резкие и сильные скачки, сразу отрываясь при этом от субстрата. Эти резкие движения осуществляются при помощи двух мощных хвостовых цирр (рис. 95), которые в обычном «ползании» участия не принимают.По краю тела справа и слева расположены два ряда краевых (маргинальных) цирр. С правого края животного они проходят вдоль всего тела, с левого же края доходят лишь до области перистома. Эти ресничные образования служат для поступательного движения животного, когда оно оторвано от субстрата и свободно плавает в воде.Mы видим, таким образом, что разнообразный и специализированный ресничный аппарат стилонихии позволяет ей делать весьма разнообразные движения в отличие, например, от простого скольжения в воде, как у туфельки или трубача.Сложно устроен также ресничный аппарат, связанный с функцией питания. Мы видели уже, что околоротовое углубление (перистом), на дне которого помещается ротовое отверстие, ведущее в глотку, расположен в передней половине животного слева. По левому краю начиная с самого переднего конца тела проходит сильно развитая зона околоротовых (адоральных) мембранелл. Своим биением они направляют ток воды в сторону ротового отверстия. Кроме того, в области перистомального углубления расположены еще три сократимые перепонки (мембраны), своими внутренними концами заходящие в глотку, и ряд особых околоротовых ресничек (рис. 95). Весь этот сложный аппарат служит для улавливания и направления пищи в ротовое отверстие.Стилонихия относится к числу простейших с очень широким диапазоном пищевых объектов. Ее с полным правом можно назвать всеядным животным. Она может питаться, как и туфелька, бактериями. В число ее пищевых объектов входят жгутиконосцы, одноклеточные водоросли (нередко диатомовые). Наконец, стилонихия может быть и хищником, нападая на другие, более мелкие виды инфузорий и поглощая их.У стилонихии имеется сократительная вакуоля. Она состоит из центрального резервуара, расположенного у левого заднего конца перистома, и одного направленного назад приводящего канала.Ядерный аппарат, как всегда у инфузорий, состоит из макронуклеуса и микронуклеуса.Макронуклеус слагается из двух половинок, соединенных тонкой перетяжкой; микронуклеусов два, они расположены непосредственно около обеих половинок Ма.Стилонихия, отчасти бурсария, трубач — это все инфузории с широким диапазоном пищевых объектов. Способность поглощать различную пищу свойственна большинству инфузорий. Однако среди них можно найти и такие виды, которые в отношении характера пищи строго специализированы.ИНФУЗОРИИ-ХИЩНИКИСреди инфузорий есть хищники, которые очень «разборчивы» в отношении своей жертвы. Хорошим примером может служить инфузория дидинии (Didinium nasutum). Дидиний — относительно небольшая инфузория, длиной в среднем около 0, 1—0, 15 мм. Передний конец вытянут в виде хоботка, на конце которого помещается ротовое отверстие. Ресничный аппарат представлен двумя венчиками ресничек (рис. 96). Дидиний быстро плавает в воде, часто меняя направление движения. Предпочитаемая пища дидиниев — инфузории туфельки. В данном случае хищник оказывается меньше своей жертвы. Дидиний внедряется в добычу хоботом, а затем, постепенно все более и более расширяя ротовое отверстие, проглатывает туфельку целиком! В хоботке имеется особый, так называемый палочковый, аппарат. Он состоит из ряда эластичных прочных палочек, располагающихся в цитоплазме по периферии хоботка. Предполагают, что этот аппарат увеличивает прочность стенок хоботка, который не разрывается при проглатывании такой огромной по сравнению с дидинием добычи, как туфелька. Дидиний — пример крайнего случая хищничества среди простейших. Если сравнить заглатывание дидинием своей добычи — туфельки —с хищничеством у высших животных, то аналогичные примеры найти трудно.
Тело стилонихии (как и большинства других брюхоресничных инфузорий) сильно сплющено в спинно-брюшном направлении, и у нее ясно различимы спинная и брюшная стороны, передний и задний концы. Спереди тело несколько расширено, сзади — сужено. При рассматривании животного с брюшной стороны ясно видно, что в передней трети слева расположен сложно устроенный перистом и ротовое отверстие.На спинной стороне довольно редко расположены реснички, которые не способны к биению. Их скорее можно назвать тонкими эластичными щетинками. Они неподвижны и не имеют отношения к функции движения. Этим ресничкам приписывают обычно осязательную, чувствительную, функцию.Все ресничные образования, связанные с движением и захватом пищи, сосредоточены на брюшной стороне животного (рис. 95). Здесь имеется небольшое количество толстых пальцевидных образований, расположенных несколькими группами. Это брюшные цирры. Каждая из них — сложное ресничное образование, результат тесного соединения (слипания) многих десятков отдельных ресничек. Таким образом, цирры представляют собой как бы кисточки, отдельные волоски которых тесно сближены и соединены между собой.При помощи цирр животное довольно быстро передвигается, «бегает» по субстрату. Кроме «ползания» и «бегания» по субстрату стилонихия способна производить довольно резкие и сильные скачки, сразу отрываясь при этом от субстрата. Эти резкие движения осуществляются при помощи двух мощных хвостовых цирр (рис. 95), которые в обычном «ползании» участия не принимают.По краю тела справа и слева расположены два ряда краевых (маргинальных) цирр. С правого края животного они проходят вдоль всего тела, с левого же края доходят лишь до области перистома. Эти ресничные образования служат для поступательного движения животного, когда оно оторвано от субстрата и свободно плавает в воде.Mы видим, таким образом, что разнообразный и специализированный ресничный аппарат стилонихии позволяет ей делать весьма разнообразные движения в отличие, например, от простого скольжения в воде, как у туфельки или трубача.Сложно устроен также ресничный аппарат, связанный с функцией питания. Мы видели уже, что околоротовое углубление (перистом), на дне которого помещается ротовое отверстие, ведущее в глотку, расположен в передней половине животного слева. По левому краю начиная с самого переднего конца тела проходит сильно развитая зона околоротовых (адоральных) мембранелл. Своим биением они направляют ток воды в сторону ротового отверстия. Кроме того, в области перистомального углубления расположены еще три сократимые перепонки (мембраны), своими внутренними концами заходящие в глотку, и ряд особых околоротовых ресничек (рис. 95). Весь этот сложный аппарат служит для улавливания и направления пищи в ротовое отверстие.Стилонихия относится к числу простейших с очень широким диапазоном пищевых объектов. Ее с полным правом можно назвать всеядным животным. Она может питаться, как и туфелька, бактериями. В число ее пищевых объектов входят жгутиконосцы, одноклеточные водоросли (нередко диатомовые). Наконец, стилонихия может быть и хищником, нападая на другие, более мелкие виды инфузорий и поглощая их.У стилонихии имеется сократительная вакуоля. Она состоит из центрального резервуара, расположенного у левого заднего конца перистома, и одного направленного назад приводящего канала.Ядерный аппарат, как всегда у инфузорий, состоит из макронуклеуса и микронуклеуса.Макронуклеус слагается из двух половинок, соединенных тонкой перетяжкой; микронуклеусов два, они расположены непосредственно около обеих половинок Ма.Стилонихия, отчасти бурсария, трубач — это все инфузории с широким диапазоном пищевых объектов. Способность поглощать различную пищу свойственна большинству инфузорий. Однако среди них можно найти и такие виды, которые в отношении характера пищи строго специализированы.ИНФУЗОРИИ-ХИЩНИКИСреди инфузорий есть хищники, которые очень «разборчивы» в отношении своей жертвы. Хорошим примером может служить инфузория дидинии (Didinium nasutum). Дидиний — относительно небольшая инфузория, длиной в среднем около 0, 1—0, 15 мм. Передний конец вытянут в виде хоботка, на конце которого помещается ротовое отверстие. Ресничный аппарат представлен двумя венчиками ресничек (рис. 96). Дидиний быстро плавает в воде, часто меняя направление движения. Предпочитаемая пища дидиниев — инфузории туфельки. В данном случае хищник оказывается меньше своей жертвы. Дидиний внедряется в добычу хоботом, а затем, постепенно все более и более расширяя ротовое отверстие, проглатывает туфельку целиком! В хоботке имеется особый, так называемый палочковый, аппарат. Он состоит из ряда эластичных прочных палочек, располагающихся в цитоплазме по периферии хоботка. Предполагают, что этот аппарат увеличивает прочность стенок хоботка, который не разрывается при проглатывании такой огромной по сравнению с дидинием добычи, как туфелька. Дидиний — пример крайнего случая хищничества среди простейших. Если сравнить заглатывание дидинием своей добычи — туфельки —с хищничеством у высших животных, то аналогичные примеры найти трудно. Дидиний, проглотивший парамецию, разумеется, очень сильно раздувается. Процесс переваривания протекает очень быстро, при комнатной температуре он занимает всего около двух часов. Затем непереваренные остатки выбрасываются наружу и дидиний начинает охотиться за очередной жертвой. Специальными исследованиями было выяснено, что суточный «рацион» дидиния составляет 12 туфелек — поистине колоссальный аппетит! Нужно иметь в виду, что в промежутках между очередными «охотами» дидинии иногда делятся. При недостатке пищи дидинии очень легко инцистируются и так же легко вновь выходят из цист.РАСТИТЕЛЬНОЯДНЫЕ ИНФУЗОРИИГораздо реже, чем хищничество, встречается среди инфузорий «чистое вегетарианство» — питание исключительно растительной пищей. Одним из немногих примеров инфузорий-«вегетарианцев» могут служить представители рода пассула (Nassula). Объектом питания их являются нитчатые сине-зеленые водоросли (рис. 97).
Дидиний, проглотивший парамецию, разумеется, очень сильно раздувается. Процесс переваривания протекает очень быстро, при комнатной температуре он занимает всего около двух часов. Затем непереваренные остатки выбрасываются наружу и дидиний начинает охотиться за очередной жертвой. Специальными исследованиями было выяснено, что суточный «рацион» дидиния составляет 12 туфелек — поистине колоссальный аппетит! Нужно иметь в виду, что в промежутках между очередными «охотами» дидинии иногда делятся. При недостатке пищи дидинии очень легко инцистируются и так же легко вновь выходят из цист.РАСТИТЕЛЬНОЯДНЫЕ ИНФУЗОРИИГораздо реже, чем хищничество, встречается среди инфузорий «чистое вегетарианство» — питание исключительно растительной пищей. Одним из немногих примеров инфузорий-«вегетарианцев» могут служить представители рода пассула (Nassula). Объектом питания их являются нитчатые сине-зеленые водоросли (рис. 97). Они проникают в эндоплазму через рот, расположенный сбоку, а затем закручиваются инфузорией в плотную спираль, которая постепенно переваривается. Пигменты водорослей частично поступают в цитоплазму инфузории и окрашивают ее в яркий темно-зеленый цвет.СУВОЙКА (VORTICELLA NEBULIFERA)Интересную и довольно большую по числу видов группу инфузорий составляют сидячие, прикрепленные к субстрату формы, образующие отряд кругоресничных (Peritricha). Широко распространенными представителями этой группы являются сувойки (виды рода Vorticella).Сувойки напоминают изящный цветок вроде колокольчика или ландыша, сидящий на длинном стебельке, который своим концом прикреплен к субстрату. Большую часть жизни сувойка проводит в прикрепленном к субстрату состоянии.
Они проникают в эндоплазму через рот, расположенный сбоку, а затем закручиваются инфузорией в плотную спираль, которая постепенно переваривается. Пигменты водорослей частично поступают в цитоплазму инфузории и окрашивают ее в яркий темно-зеленый цвет.СУВОЙКА (VORTICELLA NEBULIFERA)Интересную и довольно большую по числу видов группу инфузорий составляют сидячие, прикрепленные к субстрату формы, образующие отряд кругоресничных (Peritricha). Широко распространенными представителями этой группы являются сувойки (виды рода Vorticella).Сувойки напоминают изящный цветок вроде колокольчика или ландыша, сидящий на длинном стебельке, который своим концом прикреплен к субстрату. Большую часть жизни сувойка проводит в прикрепленном к субстрату состоянии.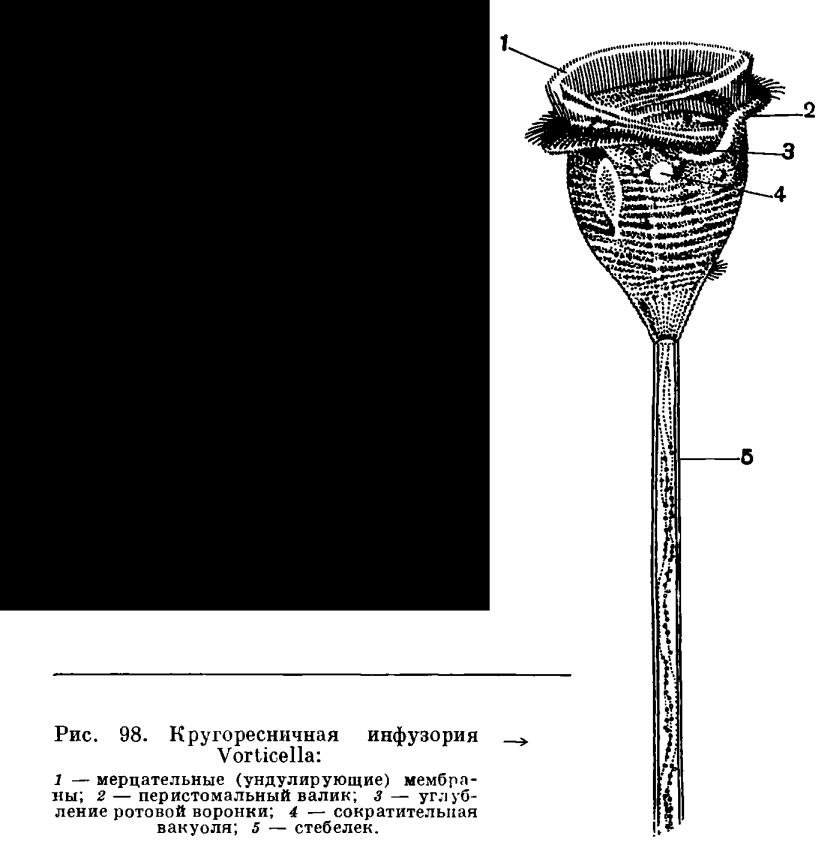 Рассмотрим строение тела инфузории. У разных видов их размеры варьируют в довольно широких пределах (примерно до 150 мк). Ротовой диск (рис. 98) расположен на расширенном переднем участке тела, которое совершенно лишено ресничек. Ресничный аппарат расположен лишь по краю ротового (перистомального) диска (рис. 98) в особой бороздке, снаружи от которой образуется валик (перистомальная губа). По краю валика идут три мерцательные мембраны, из которых две расположены вертикально, одна (наружная) — горизонтально. Они образуют несколько больше одного полного оборота спирали. Эти мембраны находятся в постоянном мерцательном движении, направляя ток воды к ротовому отверстию. Ротовой аппарат начинается довольно глубоко воронкой у края перистомального поля (рис. 98), в глубине которой располагается ротовое отверстие, ведущее в короткую глотку. Сувойки, так же как и туфельки, питаются бактериями. Их ротовое отверстие постоянно открыто, и возникает непрерывный ток воды в направлении рта.Одна сократительная вакуоля без приводящих каналов расположена недалеко от ротового отверстия. Макронуклеус имеет лентовидную или колбасовидную форму, к нему тесно примыкает единственный маленький микронуклеус.Сувойка способна резко сокращать стебелек, который в долю секунды закручивается штопором. Одновременно с этим сокращается и тело инфузории: перистомальный диск и мембраны втягиваются внутрь и весь передний конец замыкается.
Рассмотрим строение тела инфузории. У разных видов их размеры варьируют в довольно широких пределах (примерно до 150 мк). Ротовой диск (рис. 98) расположен на расширенном переднем участке тела, которое совершенно лишено ресничек. Ресничный аппарат расположен лишь по краю ротового (перистомального) диска (рис. 98) в особой бороздке, снаружи от которой образуется валик (перистомальная губа). По краю валика идут три мерцательные мембраны, из которых две расположены вертикально, одна (наружная) — горизонтально. Они образуют несколько больше одного полного оборота спирали. Эти мембраны находятся в постоянном мерцательном движении, направляя ток воды к ротовому отверстию. Ротовой аппарат начинается довольно глубоко воронкой у края перистомального поля (рис. 98), в глубине которой располагается ротовое отверстие, ведущее в короткую глотку. Сувойки, так же как и туфельки, питаются бактериями. Их ротовое отверстие постоянно открыто, и возникает непрерывный ток воды в направлении рта.Одна сократительная вакуоля без приводящих каналов расположена недалеко от ротового отверстия. Макронуклеус имеет лентовидную или колбасовидную форму, к нему тесно примыкает единственный маленький микронуклеус.Сувойка способна резко сокращать стебелек, который в долю секунды закручивается штопором. Одновременно с этим сокращается и тело инфузории: перистомальный диск и мембраны втягиваются внутрь и весь передний конец замыкается. Естественно возникает вопрос: поскольку сувойки прикреплены к субстрату, каким путем осуществляется их расселение по водоему? Это происходит путем образования свободноплавающей стадии— бродяжки. На заднем конце тела инфузории возникает венчик ресничек (рис. 99). Одновременно перистомальный диск втягивается внутрь и инфузория отделяется от стебелька. Образовавшаяся бродяжка способна плавать в течение нескольких часов. Затем события разыгрываются в обратном порядке: инфузория прикрепляется к субстрату задним концом, вырастает стебелек, редуцируется задний венчик ресничек, на переднем конце расправляется перистомальный диск, начинают работать адоральные мембраны. Образование бродяжек у сувойки нередко связано с процессом бесполого размножения. Инфузория на стебельке делится, причем одна из дочерних особей (а иногда и обе) становится бродяжкой и уплывает.Многие виды сувоек при неблагоприятных условиях способны инцистироваться.Среди сидячих инфузорий, относящихся к отряду кругоресничных, лишь относительно немногие виды вроде рассмотренных выше сувоек являются одиночно живущими формами. Большая часть относящихся сюда видов — организмы колониальные.Обычно колониальность возникает в результате не вполне завершенного бесполого или вегетативного размножения. Образующиеся в результате размножения особи в большей или меньшей степени сохраняют связь друг с другом и все вместе образуют органическую индивидуальность высшего порядка, объединяющую большие количества отдельных особей, которая и получает название колонии (с примерами колониальных организмов мы уже познакомились в классе жгутиконосцев.
Естественно возникает вопрос: поскольку сувойки прикреплены к субстрату, каким путем осуществляется их расселение по водоему? Это происходит путем образования свободноплавающей стадии— бродяжки. На заднем конце тела инфузории возникает венчик ресничек (рис. 99). Одновременно перистомальный диск втягивается внутрь и инфузория отделяется от стебелька. Образовавшаяся бродяжка способна плавать в течение нескольких часов. Затем события разыгрываются в обратном порядке: инфузория прикрепляется к субстрату задним концом, вырастает стебелек, редуцируется задний венчик ресничек, на переднем конце расправляется перистомальный диск, начинают работать адоральные мембраны. Образование бродяжек у сувойки нередко связано с процессом бесполого размножения. Инфузория на стебельке делится, причем одна из дочерних особей (а иногда и обе) становится бродяжкой и уплывает.Многие виды сувоек при неблагоприятных условиях способны инцистироваться.Среди сидячих инфузорий, относящихся к отряду кругоресничных, лишь относительно немногие виды вроде рассмотренных выше сувоек являются одиночно живущими формами. Большая часть относящихся сюда видов — организмы колониальные.Обычно колониальность возникает в результате не вполне завершенного бесполого или вегетативного размножения. Образующиеся в результате размножения особи в большей или меньшей степени сохраняют связь друг с другом и все вместе образуют органическую индивидуальность высшего порядка, объединяющую большие количества отдельных особей, которая и получает название колонии (с примерами колониальных организмов мы уже познакомились в классе жгутиконосцев. Колонии кругоресничных инфузорий образуются в результате того, что разделившиеся особи не превращаются в бродяжек, а сохраняют связь друг с другом при помощи стебельков (рис. 100). При этом основной стебелек колонии, так же как и его первые разветвления, не может быть отнесен ни к одной из особей, а принадлежит всей колонии в целом. Иногда колония состоит лишь из небольшого числа особей, у других же видов инфузорий число отдельных особей колонии может достигать нескольких сотен. Однако рост любой колонии не беспределен. По достижении характерных для данного вида размеров колония перестает увеличиваться и образующиеся в результате деления особи развивают венчик ресничек, становятся бродяжками и уплывают, давая начало новым колониям.Колонии кругоресничных инфузорий бывают двух типов. У одних стебелек колонии несократим: при раздражении сокращаются лишь отдельные особи колонии, втягивая перистом, вся же колония в целом не претерпевает изменений (к такому типу колоний относятся, например, роды Epistylis, Opercularia). У других (например, род Carchesium) стебелек всей колонии способен сокращаться, так как цитоплазма проходит внутри всех веточек и связывает, таким образом, между собой всех особей колонии. При раздражении таких колоний они сокращаются целиком. Вся колония в данном случае реагирует как единое целое, как органическая индивидуальность.Среди всех колониальных кругоресничных инфузорий особый интерес представляет, пожалуй, зоотамний (Zoothamnium arbuscula). Колонии этой инфузории отличаются особенной правильностью строения. Кроме того, в пределах колонии здесь намечается интересное биологическое явление полиморфизма.
Колонии кругоресничных инфузорий образуются в результате того, что разделившиеся особи не превращаются в бродяжек, а сохраняют связь друг с другом при помощи стебельков (рис. 100). При этом основной стебелек колонии, так же как и его первые разветвления, не может быть отнесен ни к одной из особей, а принадлежит всей колонии в целом. Иногда колония состоит лишь из небольшого числа особей, у других же видов инфузорий число отдельных особей колонии может достигать нескольких сотен. Однако рост любой колонии не беспределен. По достижении характерных для данного вида размеров колония перестает увеличиваться и образующиеся в результате деления особи развивают венчик ресничек, становятся бродяжками и уплывают, давая начало новым колониям.Колонии кругоресничных инфузорий бывают двух типов. У одних стебелек колонии несократим: при раздражении сокращаются лишь отдельные особи колонии, втягивая перистом, вся же колония в целом не претерпевает изменений (к такому типу колоний относятся, например, роды Epistylis, Opercularia). У других (например, род Carchesium) стебелек всей колонии способен сокращаться, так как цитоплазма проходит внутри всех веточек и связывает, таким образом, между собой всех особей колонии. При раздражении таких колоний они сокращаются целиком. Вся колония в данном случае реагирует как единое целое, как органическая индивидуальность.Среди всех колониальных кругоресничных инфузорий особый интерес представляет, пожалуй, зоотамний (Zoothamnium arbuscula). Колонии этой инфузории отличаются особенной правильностью строения. Кроме того, в пределах колонии здесь намечается интересное биологическое явление полиморфизма. Колония зоотамния имеет вид зонтика. На одном, главном, стебельке колонии располагаются вторичные ветви (рис. 101). Размер взрослой колонии 2—3 мм, так что они хорошо видны простым глазом. Живут зоотамний в небольших прудах с чистой водой. Колонии их обычно находят на подводных растениях, чаще всего на элодее (водяной чуме).Стебельки колонии зоотамния обладают сократимостью, так как сократимая цитоплазма проходит через все ветви колонии, за исключением базальной части главного стебелька. При сокращении, которое происходит очень быстро и резко, вся колония собирается в комочек.
Колония зоотамния имеет вид зонтика. На одном, главном, стебельке колонии располагаются вторичные ветви (рис. 101). Размер взрослой колонии 2—3 мм, так что они хорошо видны простым глазом. Живут зоотамний в небольших прудах с чистой водой. Колонии их обычно находят на подводных растениях, чаще всего на элодее (водяной чуме).Стебельки колонии зоотамния обладают сократимостью, так как сократимая цитоплазма проходит через все ветви колонии, за исключением базальной части главного стебелька. При сокращении, которое происходит очень быстро и резко, вся колония собирается в комочек. Для зоотамния характерно строго закономерное расположение ветвей. К субстрату прикрепляется один главный стебелек. От верхней части его в плоскости, перпендикулярной к стебельку, отходит девять главных ветвей колонии, строго закономерно расположенных друг относительно друга (рис. 102, 6). От этих ветвей отходят вторичные веточки, на которых сидят отдельные особи колонии. На каждой вторичной веточке может быть до 50 инфузорий. Общее количество особей в составе колонии достигает 2—3 тыс.Большая часть особей колонии по своему строению напоминает небольших одиночных сувоек, размером 40—60 мк. Но кроме мелких особей, которые называются микрозоидами, на взрослых колониях, примерно на середине главных ветвей, развиваются особи совершенно другого вида и размера (рис. 102, 5). Это крупные шаровидные особи диаметром 200—250 мк, превосходящие по своей массе объем микрозоида в сотню и более раз. Крупные особи получили название макрозоидов.По своему строению они существенно отличаются от мелких особей колонии. Перистом у них не выражен: он втягивается внутрь и не функционирует. Макрозоид с самого начала своего развития из микрозоида перестает самостоятельно принимать пищу. У него отсутствуют пищеварительные вакуоли. Рост макрозоида осуществляется, очевидно, за счет веществ, поступающих через цитоплазматические мостики, соединяющие между собой всех особей колонии. В участке тела макрозоида, которым он прикрепляется к стебельку, имеется скопление особых зернышек (гранул), которые, как увидим, в его дальнейшей судьбе играют существенную роль. Что же представляют собой эти крупные шаровидные макрозоиды, какова их биологическая роль в жизни колонии зоотамния? Наблюдение показывает, что макрозоиды — это будущие бродяжки, из которых развиваются новые колонии. Достигнув предельного размера, макрозоид развивает венчик ресничек, отделяется от колонии и уплывает. Его форма при этом несколько меняется, из шаровидного он становится коническим. Через некоторое время бродяжка прикрепляется к субстрату всегда той стороной, на которой расположена зернистость. Сразу же начинается образование и рост стебелька, причем на построение стебелька затрачиваются гранулы, которые локализованы на заднем конце бродяжки. По мере роста стебелька зернистость исчезает. После того как стебелек достигнет окончательной, характерной для зоотамния длины, начинается ряд быстро следующих друг за другом делений, ведущих к образованию колонии. Деления эти совершаются в строго определенной последовательности (рис. 102).На рассмотрении деталей этого процесса мы останавливаться не будем. Обратим лишь внимание на следующее интересное явление. Во время первых делений бродяжки зоотамния при развитии колонии у образующих особей перистом и рот не функционируют. Питание начинается позже, когда молодая колония состоит уже из 12—16 особей. Таким образом, все первые стадии развития колонии осуществляются исключительно за счет тех запасов, которые образовались в теле макрозоида в период его роста и развития на материнской колонии. Существует бесспорное сходство между развитием бродяжки зоотамния и развитием яйца у многоклеточных животных. Это сходство выражается в том, что развитие в обоих случаях осуществляется за счет ранее накопленных запасов, без восприятия пищи из внешней среды.При изучении сидячих кругоресничных инфузорий возникает вопрос: а как же осуществляется у них характерная для инфузорий форма полового процесса — конъюгация? Оказывается, что в связи с сидячим образом жизни она претерпевает некоторые существенные изменения. К началу полового процесса на колонии образуются особые, очень мелкие бродяжки. Активно двигаясь при помощи венчика ресничек, они ползают в течение некоторого времени по колонии, а затем вступают в конъюгацию с крупными нормальными сидячими особями колонии. Таким образом, здесь происходит дифференцировка конъюгантов на две группы особей: мелкие, подвижные (микроконъюганты) и более крупные, неподвижные (макроконъюганты). Эта дифференцировка конъюгантов на две категории, из которых одна (микроконъюганты) подвижная, явилась необходимым приспособлением к сидячему образу жизни. Без этого нормальный ход полового процесса (конъюгации) не мог быть, очевидно, обеспечен.СОСУЩИЕ ИНФУЗОРИИ (SUCTORIA)Весьма своеобразную в отношении способа питания группу представляют сосущие инфузории (Suctoria). Эти организмы, подобно сувойкам и другим кругоресничным инфузориям, сидячие. Количество относящихся к этому отряду видов измеряется несколькими десятками. Форма тела сосущих инфузорий весьма разнообразна. Некоторые характерные виды их изображены на рисунке 103. Одни сидят на субстрате на более или менее длинных стебельках, другие не имеют стебельков, у некоторых тело довольно сильно ветвится и т. п. Однако, несмотря на разнообразие формы, все сосущие инфузории характеризуются следующими двумя признаками:1) полным отсутствием (у взрослых форм) ресничного аппарата,2) наличием особых придатков — щупалец, служащих для высасывания добычи.
Для зоотамния характерно строго закономерное расположение ветвей. К субстрату прикрепляется один главный стебелек. От верхней части его в плоскости, перпендикулярной к стебельку, отходит девять главных ветвей колонии, строго закономерно расположенных друг относительно друга (рис. 102, 6). От этих ветвей отходят вторичные веточки, на которых сидят отдельные особи колонии. На каждой вторичной веточке может быть до 50 инфузорий. Общее количество особей в составе колонии достигает 2—3 тыс.Большая часть особей колонии по своему строению напоминает небольших одиночных сувоек, размером 40—60 мк. Но кроме мелких особей, которые называются микрозоидами, на взрослых колониях, примерно на середине главных ветвей, развиваются особи совершенно другого вида и размера (рис. 102, 5). Это крупные шаровидные особи диаметром 200—250 мк, превосходящие по своей массе объем микрозоида в сотню и более раз. Крупные особи получили название макрозоидов.По своему строению они существенно отличаются от мелких особей колонии. Перистом у них не выражен: он втягивается внутрь и не функционирует. Макрозоид с самого начала своего развития из микрозоида перестает самостоятельно принимать пищу. У него отсутствуют пищеварительные вакуоли. Рост макрозоида осуществляется, очевидно, за счет веществ, поступающих через цитоплазматические мостики, соединяющие между собой всех особей колонии. В участке тела макрозоида, которым он прикрепляется к стебельку, имеется скопление особых зернышек (гранул), которые, как увидим, в его дальнейшей судьбе играют существенную роль. Что же представляют собой эти крупные шаровидные макрозоиды, какова их биологическая роль в жизни колонии зоотамния? Наблюдение показывает, что макрозоиды — это будущие бродяжки, из которых развиваются новые колонии. Достигнув предельного размера, макрозоид развивает венчик ресничек, отделяется от колонии и уплывает. Его форма при этом несколько меняется, из шаровидного он становится коническим. Через некоторое время бродяжка прикрепляется к субстрату всегда той стороной, на которой расположена зернистость. Сразу же начинается образование и рост стебелька, причем на построение стебелька затрачиваются гранулы, которые локализованы на заднем конце бродяжки. По мере роста стебелька зернистость исчезает. После того как стебелек достигнет окончательной, характерной для зоотамния длины, начинается ряд быстро следующих друг за другом делений, ведущих к образованию колонии. Деления эти совершаются в строго определенной последовательности (рис. 102).На рассмотрении деталей этого процесса мы останавливаться не будем. Обратим лишь внимание на следующее интересное явление. Во время первых делений бродяжки зоотамния при развитии колонии у образующих особей перистом и рот не функционируют. Питание начинается позже, когда молодая колония состоит уже из 12—16 особей. Таким образом, все первые стадии развития колонии осуществляются исключительно за счет тех запасов, которые образовались в теле макрозоида в период его роста и развития на материнской колонии. Существует бесспорное сходство между развитием бродяжки зоотамния и развитием яйца у многоклеточных животных. Это сходство выражается в том, что развитие в обоих случаях осуществляется за счет ранее накопленных запасов, без восприятия пищи из внешней среды.При изучении сидячих кругоресничных инфузорий возникает вопрос: а как же осуществляется у них характерная для инфузорий форма полового процесса — конъюгация? Оказывается, что в связи с сидячим образом жизни она претерпевает некоторые существенные изменения. К началу полового процесса на колонии образуются особые, очень мелкие бродяжки. Активно двигаясь при помощи венчика ресничек, они ползают в течение некоторого времени по колонии, а затем вступают в конъюгацию с крупными нормальными сидячими особями колонии. Таким образом, здесь происходит дифференцировка конъюгантов на две группы особей: мелкие, подвижные (микроконъюганты) и более крупные, неподвижные (макроконъюганты). Эта дифференцировка конъюгантов на две категории, из которых одна (микроконъюганты) подвижная, явилась необходимым приспособлением к сидячему образу жизни. Без этого нормальный ход полового процесса (конъюгации) не мог быть, очевидно, обеспечен.СОСУЩИЕ ИНФУЗОРИИ (SUCTORIA)Весьма своеобразную в отношении способа питания группу представляют сосущие инфузории (Suctoria). Эти организмы, подобно сувойкам и другим кругоресничным инфузориям, сидячие. Количество относящихся к этому отряду видов измеряется несколькими десятками. Форма тела сосущих инфузорий весьма разнообразна. Некоторые характерные виды их изображены на рисунке 103. Одни сидят на субстрате на более или менее длинных стебельках, другие не имеют стебельков, у некоторых тело довольно сильно ветвится и т. п. Однако, несмотря на разнообразие формы, все сосущие инфузории характеризуются следующими двумя признаками:1) полным отсутствием (у взрослых форм) ресничного аппарата,2) наличием особых придатков — щупалец, служащих для высасывания добычи.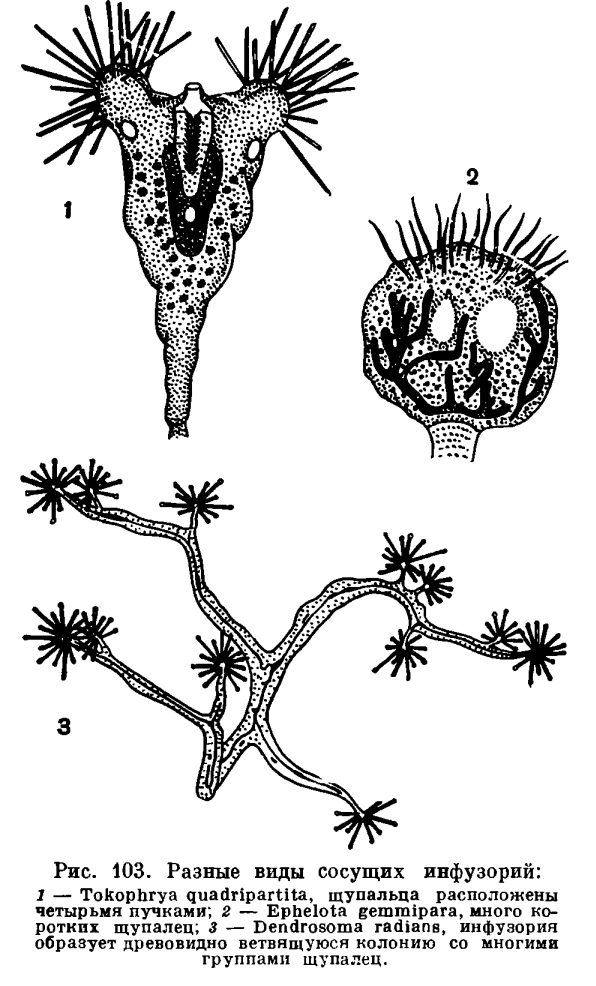 У разных видов сосущих инфузорий количество щупалец неодинаково. Нередко они бывают собраны группами. При большом увеличении микроскопа можно рассмотреть, что на конце щупальце снабжено небольшим булавовидным утолщением.Как функционируют щупальца? На этот вопрос нетрудно ответить, наблюдая в течение некоторого времени за сосущими инфузориями. Если какое-нибудь небольшое простейшее (жгутиконосец, инфузория) прикоснется к щупальцу суктории, то оно к нему мгновенно прилипнет. Все попытки жертвы оторваться бывают обычно тщетными. Если продолжить наблюдение за прилипшей к щупальцам жертвой, то можно видеть, что она постепенно начинает уменьшаться в размерах. Ее содержимое через шупальца постепенно «перекачивается» внутрь эндоплазмы сосущей инфузории до тех пор, пока от жертвы не останется одна пелликула, которая отбрасывается. Таким образом, щупальца сосущих инфузорий — это совершенно своеобразные, нигде больше в животном мире не встречающиеся органы улавливания и вместе с тем высасывания пищи (рис. 103).Сосущие инфузории — это неподвижные хищники, которые не гоняются за добычей, но мгновенно улавливают ее, если только неосторожная добыча к ним прикоснется сама.
У разных видов сосущих инфузорий количество щупалец неодинаково. Нередко они бывают собраны группами. При большом увеличении микроскопа можно рассмотреть, что на конце щупальце снабжено небольшим булавовидным утолщением.Как функционируют щупальца? На этот вопрос нетрудно ответить, наблюдая в течение некоторого времени за сосущими инфузориями. Если какое-нибудь небольшое простейшее (жгутиконосец, инфузория) прикоснется к щупальцу суктории, то оно к нему мгновенно прилипнет. Все попытки жертвы оторваться бывают обычно тщетными. Если продолжить наблюдение за прилипшей к щупальцам жертвой, то можно видеть, что она постепенно начинает уменьшаться в размерах. Ее содержимое через шупальца постепенно «перекачивается» внутрь эндоплазмы сосущей инфузории до тех пор, пока от жертвы не останется одна пелликула, которая отбрасывается. Таким образом, щупальца сосущих инфузорий — это совершенно своеобразные, нигде больше в животном мире не встречающиеся органы улавливания и вместе с тем высасывания пищи (рис. 103).Сосущие инфузории — это неподвижные хищники, которые не гоняются за добычей, но мгновенно улавливают ее, если только неосторожная добыча к ним прикоснется сама. Почему же эти своеобразные организмы мы относим к классу инфузорий? На первый взгляд они не имеют с ними ничего общего. О принадлежности сукторий к инфузориям говорят следующие факты. Во-первых, они обладают типичным для инфузорий ядерным аппаратом, состоящим из макронуклеуса и микронуклеуса. Во-вторых, во время размножения у них появляются реснички, отсутствующие «у взрослых» особей. Бесполое размножение и вместе с тем расселение сосущих инфузорий осуществляется путем образования бродяжек, снабженных несколькими кольцевыми венчиками ресничек. Образование бродяжек у сукторий может происходить по-разному. Иногда они образуются в результате не вполне равномерного деления (почкованием), при котором каждая отделяющаяся наружу почка получает участок макронуклеуса и один микронуклеус (рис. 104, Л). На одной материнской особи может образоваться сразу несколько дочерних почек (рис. 104, 5). У других видов {рис. 104, Г, Д) наблюдается очень своеобразный способ «внутреннего почкования». При этом внутри тела суктории-матери образуется полость, в которой и формируется почка-бродяжка. Наружу она выходит через специальные отверстия, сквозь которые с известным трудом «протискивается».Такое развитие зародыша внутри тела матери, а затем акт деторождения — интересная аналогия простейшего с тем, что происходит у вышестоящих многоклеточных организмов.На предыдущих страницах было рассмотрено несколько типичных свободноживущих представителей класса инфузорий, по-разному приспособленных к различным условиям среды. Интересно подойти к вопросу приспособления инфузорий к условиям жизни и, с другой стороны, посмотреть, каковы характерные общие черты инфузорий, живущих в определенных, резко очерченных условиях среды.В качестве примера возьмем две очень резко различающиеся среды обитания: жизнь в составе планктона и жизнь на дне в толще песка.ПЛАНКТОННЫЕ ИНФУЗОРИИВ составе как морского, так и пресноводного планктона встречается довольно большое число видов инфузорий.Особенно ярко выражены черты приспособлений к жизни в толще воды у радиолярий. Основная линия приспособления к планктонному образу жизни сводится к выработке таких черт строения, которые способствуют парению организма в толще воды.
Почему же эти своеобразные организмы мы относим к классу инфузорий? На первый взгляд они не имеют с ними ничего общего. О принадлежности сукторий к инфузориям говорят следующие факты. Во-первых, они обладают типичным для инфузорий ядерным аппаратом, состоящим из макронуклеуса и микронуклеуса. Во-вторых, во время размножения у них появляются реснички, отсутствующие «у взрослых» особей. Бесполое размножение и вместе с тем расселение сосущих инфузорий осуществляется путем образования бродяжек, снабженных несколькими кольцевыми венчиками ресничек. Образование бродяжек у сукторий может происходить по-разному. Иногда они образуются в результате не вполне равномерного деления (почкованием), при котором каждая отделяющаяся наружу почка получает участок макронуклеуса и один микронуклеус (рис. 104, Л). На одной материнской особи может образоваться сразу несколько дочерних почек (рис. 104, 5). У других видов {рис. 104, Г, Д) наблюдается очень своеобразный способ «внутреннего почкования». При этом внутри тела суктории-матери образуется полость, в которой и формируется почка-бродяжка. Наружу она выходит через специальные отверстия, сквозь которые с известным трудом «протискивается».Такое развитие зародыша внутри тела матери, а затем акт деторождения — интересная аналогия простейшего с тем, что происходит у вышестоящих многоклеточных организмов.На предыдущих страницах было рассмотрено несколько типичных свободноживущих представителей класса инфузорий, по-разному приспособленных к различным условиям среды. Интересно подойти к вопросу приспособления инфузорий к условиям жизни и, с другой стороны, посмотреть, каковы характерные общие черты инфузорий, живущих в определенных, резко очерченных условиях среды.В качестве примера возьмем две очень резко различающиеся среды обитания: жизнь в составе планктона и жизнь на дне в толще песка.ПЛАНКТОННЫЕ ИНФУЗОРИИВ составе как морского, так и пресноводного планктона встречается довольно большое число видов инфузорий.Особенно ярко выражены черты приспособлений к жизни в толще воды у радиолярий. Основная линия приспособления к планктонному образу жизни сводится к выработке таких черт строения, которые способствуют парению организма в толще воды.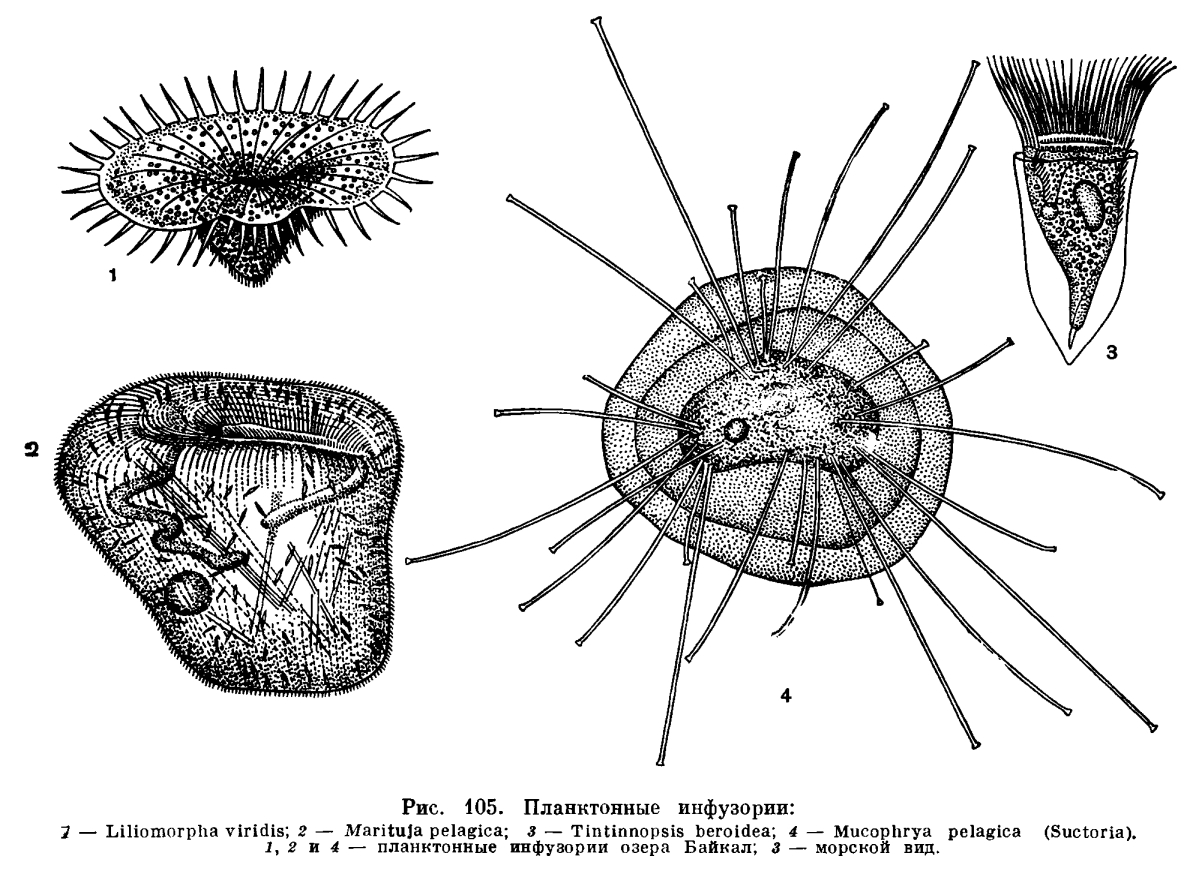 Типичным планктонным, к тому же почти исключительно морским семейством инфузорий является тинтинниды (Tintinnidae, рис. 105, 5). Общее число извест ных до сих пор видов тинтиннид около 300. Это мелкие формы, характерные тем, что протоплазматическое тело инфузории помещается в прозрачном, легком и вместе с тем прочном домике, состоящем из органического вещества. Из домика выдается наружу диск, несущий венчик ресничек, находящихся в постоянном мерцательном движении. В состоянии парения инфузорий в толще воды поддерживает здесь главным образом постоянная активная работа ресничного аппарата. Домик, очевидно, выполняет функцию защиты нижней части тела инфузории. В пресной воде обитает всего 2 вида тинтиннид (не считая 7 видов, характерных только для озера Байкал).У пресноводных инфузорий наблюдаются некоторые другие приспособления к жизни в планктоне. У многих из них цитоплазма очень сильно вакуолизирована (Loxodes, Condylostoma, Trachelius), так что напоминает пену. Это ведет к значительному уменьшению удельного веса. Все перечисленные инфузории обладают, кроме того, ресничным покровом, благодаря работе которого тело инфузории, по удельному весу лишь немногим превышающее удельный вес воды, легко поддерживается в состоянии «парения». У некоторых видов форма тела способствует увеличению удельной поверхности и облегчает парение в воде. Например, некоторые планктонные инфузории озера Байкал напоминают по форме зонтик или парашют (Liliomorpha, рис. 105, 2). Существует в озере Байкал одна планктонная сосущая инфузория (Mucophrya pelagica, рис. 105, 4), которая резко отличается от своих сидячих родичей. Этот вид лишен стебелька. Его протоплазматическое тело окружено широким слизистым футляром — приспособлением, ведущим к уменьшению веса. Наружу торчат длинные тонкие щупальца, которые наряду со своей прямой функцией, вероятно, выполняют и другую — увеличение удельной поверхности, способствующее парению в воде.Нужно, наконец, упомянуть еще об одной, так сказать, косвенной форме приспособления инфузорий к жизни в планктоне. Это прикрепление мелких инфузорий к другим организмам, ведущим планктонный образ жизни. Так, среди кругоресничных инфузорий (Peritricha) имеются довольно многочисленные виды, которые прикрепляются к планктонным веслоногим рачкам. Это обычный и нормальный для данных видов инфузорий образ жизни.Наряду с кругоресничными инфузориями и среди сосущих (Suctoria) имеются виды, поселяющиеся на планктонных организмах.ИНФУЗОРИИ, ЖИВУЩИЕ В ПЕСКЕЧрезвычайно своеобразную среду обитания представляют песчаные пляжи и отмели. По побережью морей они занимают огромные пространства и характеризуются своеобразной фауной.Проведенные за последние годы в различных странах многочисленные исследования показали, что толща многих морских песков очень богата разнообразной микроскопической или приближающейся по своим размерам к микроскопической фауной. Между частичками песка имеются многочисленные заполненные водой мелкие и мельчайшие пространства. Оказывается, что эти пространства богато заселены организмами, относящимися к самым различным группам животного мира. Здесь живут десятки видов ракообразных, кольчатые черви, круглые черви, особенно многочисленные плоские черви, некоторые моллюски, кишечнополостные. В большом количестве здесь встречаются и простейшие, главным образом инфузории. По современным данным, в состав фауны инфузорий, населяющих толщу морских песков, входит примерно 250—300 видов. Если иметь в виду не только инфузорий, а и другие группы населяющих толщу песка организмов, то общее число видов их будет очень велико. Всю совокупность животных, населяющих толщу песка, живущих в мельчайших просветах между песчинками, называют псаммофил ьной фауной.Богатство и видовой состав псаммофильной фауны определяется многими факторами. Среди них особенно важное значение имеет размер частиц песка. Крупнозернистые пески имеют бедную фауну. Бедна также фауна очень мелкозернистых заиленных песков (с диаметром частиц менее 0, 1 мм), где, очевидно, просветы между частицами слишком мелки для обитания в них животных. Наиболее богаты жизнью пески средне- и мелкозернистые.Второй фактор, играющий важную роль в развитии псаммофильной фауны, — богатство песка органическими остатками, разлагающимися органическими веществами (так называемая степень сапробности). Пески, лишенные органических веществ, бедны жизнью. С другой стороны, почти безжизненны и пески, очень богатые органическими веществами, так как распад органических веществ ведет к обеднению кислородом. Нередко к этому добавляется анаэробное сероводородное брожение.Наличие свободного сероводорода является крайне отрицательно действующим на развитие фауны фактором.В поверхностных слоях песка иногда развивается довольно богатая флора одноклеточных водорослей (диатомовые, перидиниевые). Это фактор, благоприятствующий развитию псаммофильной фауны, так как многие мелкие животные (в том числе и инфузории) питаются водорослями.Наконец, фактор, очень отрицательно действующий на псаммофильную фауну, — прибой. Это вполне понятно, так как прибой, перемывающий верхние слои песка, убивает здесь все живое. Наиболее богата псаммофильная фауна в защищенных, хорошо прогреваемых бухточках. Приливы и отливы не препятствуют развитию псаммофильной фауны. Когда в отлив вода временно уходит, обнажая песок, то в толще песка, в промежутках между песчинками, она сохраняется, и это не препятствует существованию животных.У инфузорий, входящих в состав псаммофильной фауны и относящихся к различным систематическим группам (отрядам, семействам), вырабатываются в процессе эволюции многие общие черты, являющиеся приспособлениями к своеобразным условиям существования между частицами песка.
Типичным планктонным, к тому же почти исключительно морским семейством инфузорий является тинтинниды (Tintinnidae, рис. 105, 5). Общее число извест ных до сих пор видов тинтиннид около 300. Это мелкие формы, характерные тем, что протоплазматическое тело инфузории помещается в прозрачном, легком и вместе с тем прочном домике, состоящем из органического вещества. Из домика выдается наружу диск, несущий венчик ресничек, находящихся в постоянном мерцательном движении. В состоянии парения инфузорий в толще воды поддерживает здесь главным образом постоянная активная работа ресничного аппарата. Домик, очевидно, выполняет функцию защиты нижней части тела инфузории. В пресной воде обитает всего 2 вида тинтиннид (не считая 7 видов, характерных только для озера Байкал).У пресноводных инфузорий наблюдаются некоторые другие приспособления к жизни в планктоне. У многих из них цитоплазма очень сильно вакуолизирована (Loxodes, Condylostoma, Trachelius), так что напоминает пену. Это ведет к значительному уменьшению удельного веса. Все перечисленные инфузории обладают, кроме того, ресничным покровом, благодаря работе которого тело инфузории, по удельному весу лишь немногим превышающее удельный вес воды, легко поддерживается в состоянии «парения». У некоторых видов форма тела способствует увеличению удельной поверхности и облегчает парение в воде. Например, некоторые планктонные инфузории озера Байкал напоминают по форме зонтик или парашют (Liliomorpha, рис. 105, 2). Существует в озере Байкал одна планктонная сосущая инфузория (Mucophrya pelagica, рис. 105, 4), которая резко отличается от своих сидячих родичей. Этот вид лишен стебелька. Его протоплазматическое тело окружено широким слизистым футляром — приспособлением, ведущим к уменьшению веса. Наружу торчат длинные тонкие щупальца, которые наряду со своей прямой функцией, вероятно, выполняют и другую — увеличение удельной поверхности, способствующее парению в воде.Нужно, наконец, упомянуть еще об одной, так сказать, косвенной форме приспособления инфузорий к жизни в планктоне. Это прикрепление мелких инфузорий к другим организмам, ведущим планктонный образ жизни. Так, среди кругоресничных инфузорий (Peritricha) имеются довольно многочисленные виды, которые прикрепляются к планктонным веслоногим рачкам. Это обычный и нормальный для данных видов инфузорий образ жизни.Наряду с кругоресничными инфузориями и среди сосущих (Suctoria) имеются виды, поселяющиеся на планктонных организмах.ИНФУЗОРИИ, ЖИВУЩИЕ В ПЕСКЕЧрезвычайно своеобразную среду обитания представляют песчаные пляжи и отмели. По побережью морей они занимают огромные пространства и характеризуются своеобразной фауной.Проведенные за последние годы в различных странах многочисленные исследования показали, что толща многих морских песков очень богата разнообразной микроскопической или приближающейся по своим размерам к микроскопической фауной. Между частичками песка имеются многочисленные заполненные водой мелкие и мельчайшие пространства. Оказывается, что эти пространства богато заселены организмами, относящимися к самым различным группам животного мира. Здесь живут десятки видов ракообразных, кольчатые черви, круглые черви, особенно многочисленные плоские черви, некоторые моллюски, кишечнополостные. В большом количестве здесь встречаются и простейшие, главным образом инфузории. По современным данным, в состав фауны инфузорий, населяющих толщу морских песков, входит примерно 250—300 видов. Если иметь в виду не только инфузорий, а и другие группы населяющих толщу песка организмов, то общее число видов их будет очень велико. Всю совокупность животных, населяющих толщу песка, живущих в мельчайших просветах между песчинками, называют псаммофил ьной фауной.Богатство и видовой состав псаммофильной фауны определяется многими факторами. Среди них особенно важное значение имеет размер частиц песка. Крупнозернистые пески имеют бедную фауну. Бедна также фауна очень мелкозернистых заиленных песков (с диаметром частиц менее 0, 1 мм), где, очевидно, просветы между частицами слишком мелки для обитания в них животных. Наиболее богаты жизнью пески средне- и мелкозернистые.Второй фактор, играющий важную роль в развитии псаммофильной фауны, — богатство песка органическими остатками, разлагающимися органическими веществами (так называемая степень сапробности). Пески, лишенные органических веществ, бедны жизнью. С другой стороны, почти безжизненны и пески, очень богатые органическими веществами, так как распад органических веществ ведет к обеднению кислородом. Нередко к этому добавляется анаэробное сероводородное брожение.Наличие свободного сероводорода является крайне отрицательно действующим на развитие фауны фактором.В поверхностных слоях песка иногда развивается довольно богатая флора одноклеточных водорослей (диатомовые, перидиниевые). Это фактор, благоприятствующий развитию псаммофильной фауны, так как многие мелкие животные (в том числе и инфузории) питаются водорослями.Наконец, фактор, очень отрицательно действующий на псаммофильную фауну, — прибой. Это вполне понятно, так как прибой, перемывающий верхние слои песка, убивает здесь все живое. Наиболее богата псаммофильная фауна в защищенных, хорошо прогреваемых бухточках. Приливы и отливы не препятствуют развитию псаммофильной фауны. Когда в отлив вода временно уходит, обнажая песок, то в толще песка, в промежутках между песчинками, она сохраняется, и это не препятствует существованию животных.У инфузорий, входящих в состав псаммофильной фауны и относящихся к различным систематическим группам (отрядам, семействам), вырабатываются в процессе эволюции многие общие черты, являющиеся приспособлениями к своеобразным условиям существования между частицами песка.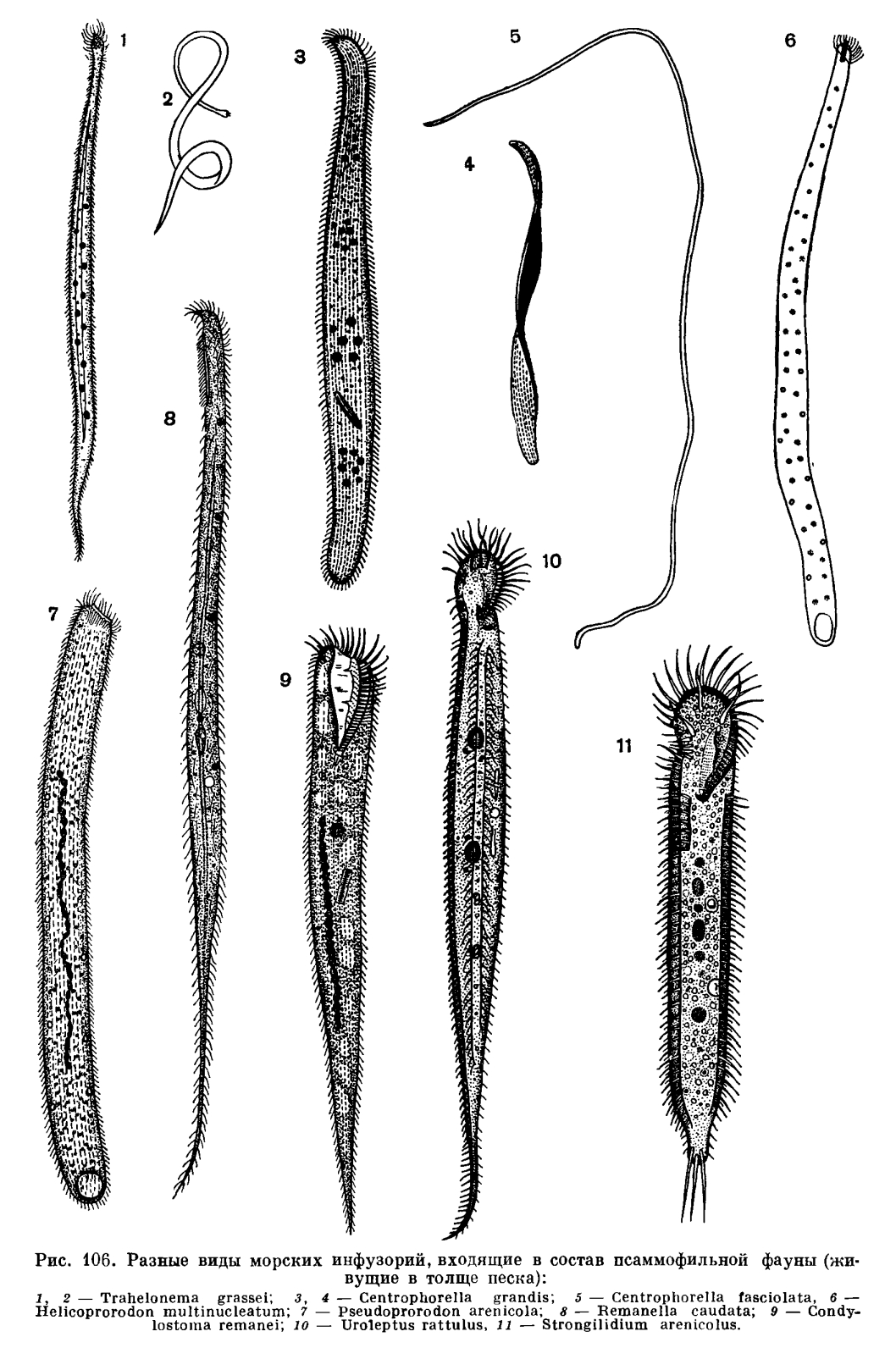 На рисунке 106 изображены некоторые виды псаммофильной фауны инфузорий, относящиеся к разным отрядам и семействам. Между ними много сходства. Тело большинства их более или менее сильно вытянуто в длину, червеобразно. Это дает возможность легко «протискиваться» в мельчайшие отверстия между песчинками. У очень многих видов (рис. 106) удлинение тела сочетается с его уплощением. Ресничный аппарат всегда хорошо развит, что позволяет активно, с известной силой, двигаться в узких просветах. Нередко реснички развиваются на одной стороне червеобразного уплощенного тела, противоположная сторона оказывается голой. Эта особенность связана, вероятно, с резко выраженной у большинства псаммофильных в»идов способностью очень тесно и очень прочно при посредстве ресничного аппарата приставать (прикрепляться) к субстрату (явление, называемое тигмотаксисом). Это свойство позволяет животным оставаться на месте в тех случаях, когда Б узких просветах, где они живут, возникают токи воды. При этом, вероятно, выгоднее, чтобы сторона, противоположная той, которой животное прикрепилось к субстрату, была гладкой.Чем питаются псаммофильные инфузории? Значительную часть «рациона» у многих видов составляют водоросли, в особенности диатомовые. Бактерии в меньшей степени служат им пищей. Это зависит в значительной мере и от того, что в песках, не сильно загрязненных, бактерий мало. Наконец, особенно среди наиболее крупных псаммофильных инфузорий имеется немалое количество хищных форм, которые поедают других инфузорий, относящихся к более мелким видам. Псаммофильные инфузории распространены, по-видимому, повсеместно.ИНФУЗОРИИ АПОСТОМАТЫВ процессе приспособления к условиям жизни очень сложные жизненные циклы, сопровождающиеся сменой нескольких форм строения, выработались у своеобразной морской группы инфузорий — апостомат (Apostomata). Сложность и своеобразие жизненного пути этих инфузорий зависит от того, что часть стадий цикла они проводят как свободноживущие организмы, другие же стадии являются паразитическими или полупаразитическими. Рассмотрим на конкретном примере жизненный цикл апостомат.
На рисунке 106 изображены некоторые виды псаммофильной фауны инфузорий, относящиеся к разным отрядам и семействам. Между ними много сходства. Тело большинства их более или менее сильно вытянуто в длину, червеобразно. Это дает возможность легко «протискиваться» в мельчайшие отверстия между песчинками. У очень многих видов (рис. 106) удлинение тела сочетается с его уплощением. Ресничный аппарат всегда хорошо развит, что позволяет активно, с известной силой, двигаться в узких просветах. Нередко реснички развиваются на одной стороне червеобразного уплощенного тела, противоположная сторона оказывается голой. Эта особенность связана, вероятно, с резко выраженной у большинства псаммофильных в»идов способностью очень тесно и очень прочно при посредстве ресничного аппарата приставать (прикрепляться) к субстрату (явление, называемое тигмотаксисом). Это свойство позволяет животным оставаться на месте в тех случаях, когда Б узких просветах, где они живут, возникают токи воды. При этом, вероятно, выгоднее, чтобы сторона, противоположная той, которой животное прикрепилось к субстрату, была гладкой.Чем питаются псаммофильные инфузории? Значительную часть «рациона» у многих видов составляют водоросли, в особенности диатомовые. Бактерии в меньшей степени служат им пищей. Это зависит в значительной мере и от того, что в песках, не сильно загрязненных, бактерий мало. Наконец, особенно среди наиболее крупных псаммофильных инфузорий имеется немалое количество хищных форм, которые поедают других инфузорий, относящихся к более мелким видам. Псаммофильные инфузории распространены, по-видимому, повсеместно.ИНФУЗОРИИ АПОСТОМАТЫВ процессе приспособления к условиям жизни очень сложные жизненные циклы, сопровождающиеся сменой нескольких форм строения, выработались у своеобразной морской группы инфузорий — апостомат (Apostomata). Сложность и своеобразие жизненного пути этих инфузорий зависит от того, что часть стадий цикла они проводят как свободноживущие организмы, другие же стадии являются паразитическими или полупаразитическими. Рассмотрим на конкретном примере жизненный цикл апостомат. Инфузория спирофрия (Spirophrya subparasitica) в инцистированном состоянии часто может быть обнаружена сидящей на небольшой ножке на мелких планктонных морских ракообразных (особенно часто на рачках рода Idia). Пока рачок активно плавает в морской воде, сидящие на нем спирофрии не претерпевают никаких изменений. Для дальнейшего развития инфузорий необходимо, чтобы рачок был съеден морским гидроидным полипом, что случается нередко (рис. 107). Как только цисты спирофрии вместе с рачком проникнут в пищеварительную полость, из них сразу же выходят мелкие инфузории, которые начинают энергично питаться пищевой кашицей, образующейся в результате переваривания заглоченного рачка. В течение часа размеры инфузории увеличиваются в 3—4 раза. Однако размножения на этой стадии не происходит. Перед нами типичная стадия роста инфузории, которую называют трофонтом. Через некоторое время вместе с непереваренными остатками пищи трофонт выбрасывается полипом наружу в морскую воду. Здесь он, активно плавая, спускается по телу полипа к его подошве, где и прикрепляется, окружаясь при этом цистой. Эта стадия инцистированной, сидящей на полипе крупной инфузории получила название томонта. Это фаза размножения. Томонт не питается, но быстро последовательно несколько раз делится (рис. 107, 7). В результате получается целая группа очень мелких инфузорий. Количество их зависит от размеров томонта, который определяется в свою очередь величиной трофонта, давшего ему начало. Мелкие инфузории, образовавшиеся в результате деления томонта (они называются томитами или бродяжками), представляют собой стадию расселения.Они выходят из цисты, быстро плавают (не питаясь при этом, а используя имеющиеся у них в цитоплазме запасы). Если им «посчастливится» натолкнуться на веслоногого рачка, то они тотчас же прикрепляются к нему и инцистируются. Это стадия, с которой мы начали рассмотрение цикла.В рассмотренном нами жизненном цикле спирофрии обращает на себя внимание резкая разграниченность стадий, имеющих разное биологическое значение. Трофонт — это стадия роста. Он только растет, накапливает большое количество цитоплазмы и всякого рода резервных веществ за счет энергичного и быстрого питания. К размножению трофонт не способен. Обратное явление наблюдается у томонта — неспособность питаться и энергичное быстрое размножение. После каждого деления не происходит роста, и поэтому размножение томонта сводится к быстрому распаду на множество бродяжек. Наконец, бродяжки выполняют свою специальную и только им свойственную функцию: это особи — расселительницы и распространительницы вида. Они не способны ни питаться, ни размножаться.ЖИЗНЕННЫЙ ЦИКЛ ИХТИОФТИРИУСАСреди инфузорий, часть жизненного цикла которых протекает в паразитическом, часть в свободноживущем состоянии, большой интерес, особенно в практическом отношении, представляет ихтиофтириус (Ichthyophthirius multifiliis). Эта инфузория в течение большей части своей жизни является паразитом рыб. Она может вызвать тяжелое заболевание, в особенности поражающее мальков, часто приводящее к гибели хозяина (рис. 108).
Инфузория спирофрия (Spirophrya subparasitica) в инцистированном состоянии часто может быть обнаружена сидящей на небольшой ножке на мелких планктонных морских ракообразных (особенно часто на рачках рода Idia). Пока рачок активно плавает в морской воде, сидящие на нем спирофрии не претерпевают никаких изменений. Для дальнейшего развития инфузорий необходимо, чтобы рачок был съеден морским гидроидным полипом, что случается нередко (рис. 107). Как только цисты спирофрии вместе с рачком проникнут в пищеварительную полость, из них сразу же выходят мелкие инфузории, которые начинают энергично питаться пищевой кашицей, образующейся в результате переваривания заглоченного рачка. В течение часа размеры инфузории увеличиваются в 3—4 раза. Однако размножения на этой стадии не происходит. Перед нами типичная стадия роста инфузории, которую называют трофонтом. Через некоторое время вместе с непереваренными остатками пищи трофонт выбрасывается полипом наружу в морскую воду. Здесь он, активно плавая, спускается по телу полипа к его подошве, где и прикрепляется, окружаясь при этом цистой. Эта стадия инцистированной, сидящей на полипе крупной инфузории получила название томонта. Это фаза размножения. Томонт не питается, но быстро последовательно несколько раз делится (рис. 107, 7). В результате получается целая группа очень мелких инфузорий. Количество их зависит от размеров томонта, который определяется в свою очередь величиной трофонта, давшего ему начало. Мелкие инфузории, образовавшиеся в результате деления томонта (они называются томитами или бродяжками), представляют собой стадию расселения.Они выходят из цисты, быстро плавают (не питаясь при этом, а используя имеющиеся у них в цитоплазме запасы). Если им «посчастливится» натолкнуться на веслоногого рачка, то они тотчас же прикрепляются к нему и инцистируются. Это стадия, с которой мы начали рассмотрение цикла.В рассмотренном нами жизненном цикле спирофрии обращает на себя внимание резкая разграниченность стадий, имеющих разное биологическое значение. Трофонт — это стадия роста. Он только растет, накапливает большое количество цитоплазмы и всякого рода резервных веществ за счет энергичного и быстрого питания. К размножению трофонт не способен. Обратное явление наблюдается у томонта — неспособность питаться и энергичное быстрое размножение. После каждого деления не происходит роста, и поэтому размножение томонта сводится к быстрому распаду на множество бродяжек. Наконец, бродяжки выполняют свою специальную и только им свойственную функцию: это особи — расселительницы и распространительницы вида. Они не способны ни питаться, ни размножаться.ЖИЗНЕННЫЙ ЦИКЛ ИХТИОФТИРИУСАСреди инфузорий, часть жизненного цикла которых протекает в паразитическом, часть в свободноживущем состоянии, большой интерес, особенно в практическом отношении, представляет ихтиофтириус (Ichthyophthirius multifiliis). Эта инфузория в течение большей части своей жизни является паразитом рыб. Она может вызвать тяжелое заболевание, в особенности поражающее мальков, часто приводящее к гибели хозяина (рис. 108). Паразитарные стадии развития ихтиофтириуса протекают непосредственно в покровах рыбы: в коже, на плавниках, очень часто на жабрах рыбы.Рыба заражается очень мелкими свободноплавающими бродяжками (20—30 мк диаметром), плавающими в воде. Бродяжки при соприкосновении с рыбой немедленно прикрепляются к ее поверхности и сразу же активными движениями внедряются в ткани. Проникнув в живую ткань, инфузории начинают активно питаться и расти. Питание осуществляется путем заглатывания при помощи помещающегося на переднем конце тела ротового отверстия клеточных элементов хозяина. Растущие в тканях рыбы ихтиофтириусы вскоре становятся видны простым глазом. Рыба как бы обсыпана манной крупой (рис. 108). Продолжительность стадии роста и паразитирования ихтиофтириуса в субэпителиальном слое рыбы в высокой степени зависит от температуры. По исследованиям Бауера, при 28—30°С этот срок составляет 4 суток, при 19—21°С—7Г при 13—15°С-10-11, а при 8-9°С период этот затягивается до 40 дней.К концу периода роста ихтиофтириус по сравнению с бродяжками достигает очень большой величины: 0, 5—1 мм в диаметре. По достижении предельной величины инфузории активными движениями выходят из тканей рыбы в воду и некоторое время медленно плавают при помощи покрывающего все их тело ресничного аппарата. Вскоре крупные ихтиофтириусы оседают на каком-нибудь подводном предмете и выделяют цисту. Тотчас же вслед за инцистированием начинаются последовательные деления инфузории: сначала пополам, затем каждая дочерняя особь делится опять на две и т. д. до 10—11 раз. В результате внутри цисты образуется до 2000 мелких почти округлых особей, покрытых ресничками. Внутри цисты бродяжки активно двигаются. Они прободают оболочку и выходят наружу. Активно плавающие бродяжки заражают новых особей рыб.Быстрота деления ихтиофтириуса в цистах, так же как и темп его роста в тканях рыб, в большой степени зависит от температуры. По исследованиям различных авторов, приводятся следующие цифры: при 26—27°С процесс развития бродяжек в цисте занимает 10—12 часов, при 15—16°С требуется 28—30 часов, при 4—5°С он тянется уже в течение 6—7 суток.Ихтиофтириус — широко распространенный и очень опасный паразит рыб. Особенно большой вред он приносит в рыбоводческих хозяйствах, где происходит нерест и выращивание мальков. Ихтиофтириус, нападая на мальков, в короткий срок приводит их к гибели. Известны многочисленные случаи поголовного уничтожения мальков как карповых, так и лососевых рыб в прудовых хозяйствах.Борьба с ихтиофтириусом представляет значительные трудности. Главное значение здесь имеют профилактические мероприятия, направленные на то, чтобы не допустить свободно плавающих в воде бродяжек проникнуть в ткани рыбы. Для этого полезно осуществлять частую пересадку больных рыб в новые водоемы или аквариумы, создавать условия проточности, что особенно эффективно в борьбе с ихтиофтириусом.ИНФУЗОРИИ ТРИХОДИНЫОчень своеобразны паразитирующие главным образом на рыбах инфузории семейства урцелариид (Urcelariidae) отряда кругоресничных.
Паразитарные стадии развития ихтиофтириуса протекают непосредственно в покровах рыбы: в коже, на плавниках, очень часто на жабрах рыбы.Рыба заражается очень мелкими свободноплавающими бродяжками (20—30 мк диаметром), плавающими в воде. Бродяжки при соприкосновении с рыбой немедленно прикрепляются к ее поверхности и сразу же активными движениями внедряются в ткани. Проникнув в живую ткань, инфузории начинают активно питаться и расти. Питание осуществляется путем заглатывания при помощи помещающегося на переднем конце тела ротового отверстия клеточных элементов хозяина. Растущие в тканях рыбы ихтиофтириусы вскоре становятся видны простым глазом. Рыба как бы обсыпана манной крупой (рис. 108). Продолжительность стадии роста и паразитирования ихтиофтириуса в субэпителиальном слое рыбы в высокой степени зависит от температуры. По исследованиям Бауера, при 28—30°С этот срок составляет 4 суток, при 19—21°С—7Г при 13—15°С-10-11, а при 8-9°С период этот затягивается до 40 дней.К концу периода роста ихтиофтириус по сравнению с бродяжками достигает очень большой величины: 0, 5—1 мм в диаметре. По достижении предельной величины инфузории активными движениями выходят из тканей рыбы в воду и некоторое время медленно плавают при помощи покрывающего все их тело ресничного аппарата. Вскоре крупные ихтиофтириусы оседают на каком-нибудь подводном предмете и выделяют цисту. Тотчас же вслед за инцистированием начинаются последовательные деления инфузории: сначала пополам, затем каждая дочерняя особь делится опять на две и т. д. до 10—11 раз. В результате внутри цисты образуется до 2000 мелких почти округлых особей, покрытых ресничками. Внутри цисты бродяжки активно двигаются. Они прободают оболочку и выходят наружу. Активно плавающие бродяжки заражают новых особей рыб.Быстрота деления ихтиофтириуса в цистах, так же как и темп его роста в тканях рыб, в большой степени зависит от температуры. По исследованиям различных авторов, приводятся следующие цифры: при 26—27°С процесс развития бродяжек в цисте занимает 10—12 часов, при 15—16°С требуется 28—30 часов, при 4—5°С он тянется уже в течение 6—7 суток.Ихтиофтириус — широко распространенный и очень опасный паразит рыб. Особенно большой вред он приносит в рыбоводческих хозяйствах, где происходит нерест и выращивание мальков. Ихтиофтириус, нападая на мальков, в короткий срок приводит их к гибели. Известны многочисленные случаи поголовного уничтожения мальков как карповых, так и лососевых рыб в прудовых хозяйствах.Борьба с ихтиофтириусом представляет значительные трудности. Главное значение здесь имеют профилактические мероприятия, направленные на то, чтобы не допустить свободно плавающих в воде бродяжек проникнуть в ткани рыбы. Для этого полезно осуществлять частую пересадку больных рыб в новые водоемы или аквариумы, создавать условия проточности, что особенно эффективно в борьбе с ихтиофтириусом.ИНФУЗОРИИ ТРИХОДИНЫОчень своеобразны паразитирующие главным образом на рыбах инфузории семейства урцелариид (Urcelariidae) отряда кругоресничных.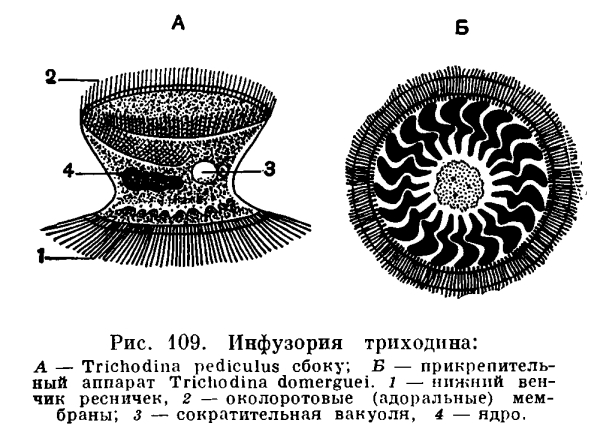 Особенно характерны многочисленные представители рода триходин (Trichodina) и некоторых других близких к нему родов (рис. 109). За немногими исключениями, все триходины и близкие к ним формы являются эктопаразитами, т. е. они живут на поверхности (а не в толще тканей и не внутри органов) своих хозяев (рыб). Место локализации триходин — кожные покровы, плавники, очень часто жабры рыб. Они встречаются как на морских, так и на пресноводных рыбах. Количество видов триходин измеряется несколькими десятками. Триходины не являются прикрепленными животными, а, напротив, активно передвигаются по поверхности своих хозяев. Вне организма хозяина, в воде, они долго жить не могут и, будучи отделены от рыбы, довольно быстро погибают.Вся система приспособлений триходин к жизни на поверхности хозяина направлена на то, чтобы не оторваться от тела хозяина (что почти всегда равносильно гибели), сохраняя при этом подвижность. Приспособления эти весьма совершенны. Тело большинства триходин имеет форму довольно плоского диска, иногда шапочки. Сторона, обращенная к телу хозяина, слегка вогнутая, она образует прикрепительную присоску. По наружному краю присоски расположен венчик хорошо развитых ресничек, при помощи которых главным образом и происходит передвижение (ползание) инфузории по поверхности тела рыбы. Этот венчик соответствует венчику, имеющемуся у бродяжек сидячих кругоресничных инфузории, рассмотренных выше. Таким образом, триходину можно сравнить с бродяжкой. На брюшной поверхности (на присоске) у триходин имеется очень сложный опорный и прикрепительный аппарат, способствующий удержанию инфузории на хозяине. Не вдаваясь в детали его строения, отметим, что основу его составляет сложной конфигурации кольцо, слагающееся из отдельных сегментов, несущих наружный и внутренний зубцы (рис. 109, Б). Это кольцо образует эластичную и вместе с тем прочную основу брюшной поверхности, выполняющей функцию присоски. Разные виды триходин отличаются друг от друга по количеству сегментов, образующих кольцо, и по конфигурации наружного и внутреннего крючьев.На противоположной от диска стороне тела триходины расположен перистом и ротовой аппарат. Строение его более или менее типично для кругоресничных инфузорий. Закрученные по часовой стрелке адоральные мембраны ведут в углубление, на дне которого расположен рот. Ядерный аппарат триходин устроен типично для инфузорий: один лентовидный макронуклеус и один расположенный рядом с ним микронуклеус. Имеется одна сократительная вакуоля.Триходины широко распространены в водоемах всех типов. Особенно часто их находят на мальках разных видов рыб. При массовом размножении триходины наносят большой вред рыбе, в особенности если массами покрывают жабры. Это нарушает нормальное дыхание рыбы.Для того чтобы очистить рыбу от триходин, рекомендуют делать лечебные ванны из 2-процентного раствора поваренной соли или 0, 01-процентного раствора марганцовокислого калия (для мальков — в течение 10—20 минут).ИНФУЗОРИИ КИШЕЧНОГО ТРАКТА КОПЫТНЫХСреди инфузорий имеется множество видов, ведущих эндопаразитический образ жизни, т. е. живущих внутри (в различных органах и тканях) животных-хозяев. Особенно многочисленны и разнообразны инфузории, населяющие кишечный тракт копытных животных. Большинство их образует самостоятельный отряд инфузорий, называемый энтодиниоморфами (Еntodiniomorpha). У жвачных (рогатый скот, овцы, козы, антилопы, олени, лоси) эти инфузории в огромных количествах населяют передние отделы желудка. Желудок жвачных состоит из чэтырех отделов — рубца, сетки, книжки и сычуга. Из них только сычуг соответствует желудку других млекопитающих: он имеет пищеварительные железы, в него выделяются пищеварительные ферменты и соляная кислота. Реакция среды в сычуге резкокислая. Первые три отдела не имеют пищеварительных желез. Их внутренняя стенка частично ороговевает. Эти три отдела по происхождению следует рассматривать как нижние расширенные части пищевода. Названия сетки и книжки даны по форме и характеру складок их внутренней стенки. Самый вместительный отдел — рубец. Заглоченная и почти не пережеванная растительная пища сначала проходит в рубец. Сюда же попадает большое количество слюны. В рубце пища подвергается сложным процессам брожения, которые увеличивают ее усвояемость. В этом отделе в огромных количествах живут разнообразные микроорганизмы, вызывающие процессы брожения, которые протекают в условиях нейтральной или слабощелочной реакции. В рубце наряду с бактериями обитает масса простейших — очень мелких жгутиконосцев и инфузорий, так как здесь создаются весьма благоприятные условия для их развития.Из рубца через сетку пища отрыгивается в ротовую полость, где дополнительно пережевывается (жвачка). Проглатываемая вновь пищевая пережеванная масса по особой трубке, образуемой складками пищевода, идет уже не в рубец, а в книжку и оттуда в сычуг, где подвергается действию пищеварительных соков жвачного. В сычуге в условиях кислой реакции и наличия пищеварительных ферментов инфузории погибают. Попадая туда со жвачкой, они перевариваются.Количество простейших в рубце (а также в сетке) может достигать колоссальных величин. Если взять каплю содержимого рубца и рассмотреть ее под микроскопом (при нагревании, так как при комнатной температуре инфузории останавливаются), то в поле зрения инфузории буквально кишат. Трудно даже в условиях культуры получить такую массу инфузорий. Количество инфузорий в 1 см3 содержимого рубца достигает миллиона, а нередко и более. В пересчете на весь объем рубца это дает поистине астрономические цифры! Богатство содержимого рубца инфузориями в большой степени зависит от характера пищи жвачного. Если пища богата клетчаткой и бедна углеводами и белками (трава, солома), то инфузорий в рубце относительно немного. При добавлении в пищевой рацион углеводов и белков (отруби) количество инфузорий резко увеличивается и достигает огромных цифр. Нужно иметь в виду, что существует постоянный отток инфузорий. Попадая вместе с жвачкой в сычуг, они погибают. Высокий уровень количества инфузорий поддерживается энергичным размножением их.У непарнокопытных (лошадь, осел, зебра) в пищевом тракте тоже имеется большое количество инфузорий, однако локализация их в хозяине иная. У непарнокопытных нет сложного желудка, благодаря чему возможность развития простейших в передних отделах пищевого тракта отсутствует. Зато у непарнокопытных очень сильно развиты толстая и слепая кишка, которые обычно забиты пищевыми массами и играют существенную роль в пищеварении. В этом отделе кишечника, подобно тому как в рубце и сетке жвачных, развивается очень богатая фауна простейших, преимущественно инфузорий, большинство которых также относится к отряду энтодиниоморф. Однако по видовому составу фауна рубца жвачных и фауна толстого кишечника непарнокопытных не совпадают.ИНФУЗОРИИ КИШЕЧНОГО ТРАКТА ЖВАЧНЫХНаибольший интерес представляют инфузории семейства офриосколецид (Ophryoscolecidae), относящегося к отряду энтодиниоморф. Характерный признак этого отряда — отсутствие сплошного ресничного покрова. Сложные ресничные образования — цирры — расположены на переднем конце тела инфузорий в области ротового отверстия. К этим основным элементам ресничного аппарата могут прибавиться еще дополнительные группы цирр, расположенные либо на переднем, либо на заднем конце тела. Общее количество видов инфузорий семейства офриосколецид — около 120.
Особенно характерны многочисленные представители рода триходин (Trichodina) и некоторых других близких к нему родов (рис. 109). За немногими исключениями, все триходины и близкие к ним формы являются эктопаразитами, т. е. они живут на поверхности (а не в толще тканей и не внутри органов) своих хозяев (рыб). Место локализации триходин — кожные покровы, плавники, очень часто жабры рыб. Они встречаются как на морских, так и на пресноводных рыбах. Количество видов триходин измеряется несколькими десятками. Триходины не являются прикрепленными животными, а, напротив, активно передвигаются по поверхности своих хозяев. Вне организма хозяина, в воде, они долго жить не могут и, будучи отделены от рыбы, довольно быстро погибают.Вся система приспособлений триходин к жизни на поверхности хозяина направлена на то, чтобы не оторваться от тела хозяина (что почти всегда равносильно гибели), сохраняя при этом подвижность. Приспособления эти весьма совершенны. Тело большинства триходин имеет форму довольно плоского диска, иногда шапочки. Сторона, обращенная к телу хозяина, слегка вогнутая, она образует прикрепительную присоску. По наружному краю присоски расположен венчик хорошо развитых ресничек, при помощи которых главным образом и происходит передвижение (ползание) инфузории по поверхности тела рыбы. Этот венчик соответствует венчику, имеющемуся у бродяжек сидячих кругоресничных инфузории, рассмотренных выше. Таким образом, триходину можно сравнить с бродяжкой. На брюшной поверхности (на присоске) у триходин имеется очень сложный опорный и прикрепительный аппарат, способствующий удержанию инфузории на хозяине. Не вдаваясь в детали его строения, отметим, что основу его составляет сложной конфигурации кольцо, слагающееся из отдельных сегментов, несущих наружный и внутренний зубцы (рис. 109, Б). Это кольцо образует эластичную и вместе с тем прочную основу брюшной поверхности, выполняющей функцию присоски. Разные виды триходин отличаются друг от друга по количеству сегментов, образующих кольцо, и по конфигурации наружного и внутреннего крючьев.На противоположной от диска стороне тела триходины расположен перистом и ротовой аппарат. Строение его более или менее типично для кругоресничных инфузорий. Закрученные по часовой стрелке адоральные мембраны ведут в углубление, на дне которого расположен рот. Ядерный аппарат триходин устроен типично для инфузорий: один лентовидный макронуклеус и один расположенный рядом с ним микронуклеус. Имеется одна сократительная вакуоля.Триходины широко распространены в водоемах всех типов. Особенно часто их находят на мальках разных видов рыб. При массовом размножении триходины наносят большой вред рыбе, в особенности если массами покрывают жабры. Это нарушает нормальное дыхание рыбы.Для того чтобы очистить рыбу от триходин, рекомендуют делать лечебные ванны из 2-процентного раствора поваренной соли или 0, 01-процентного раствора марганцовокислого калия (для мальков — в течение 10—20 минут).ИНФУЗОРИИ КИШЕЧНОГО ТРАКТА КОПЫТНЫХСреди инфузорий имеется множество видов, ведущих эндопаразитический образ жизни, т. е. живущих внутри (в различных органах и тканях) животных-хозяев. Особенно многочисленны и разнообразны инфузории, населяющие кишечный тракт копытных животных. Большинство их образует самостоятельный отряд инфузорий, называемый энтодиниоморфами (Еntodiniomorpha). У жвачных (рогатый скот, овцы, козы, антилопы, олени, лоси) эти инфузории в огромных количествах населяют передние отделы желудка. Желудок жвачных состоит из чэтырех отделов — рубца, сетки, книжки и сычуга. Из них только сычуг соответствует желудку других млекопитающих: он имеет пищеварительные железы, в него выделяются пищеварительные ферменты и соляная кислота. Реакция среды в сычуге резкокислая. Первые три отдела не имеют пищеварительных желез. Их внутренняя стенка частично ороговевает. Эти три отдела по происхождению следует рассматривать как нижние расширенные части пищевода. Названия сетки и книжки даны по форме и характеру складок их внутренней стенки. Самый вместительный отдел — рубец. Заглоченная и почти не пережеванная растительная пища сначала проходит в рубец. Сюда же попадает большое количество слюны. В рубце пища подвергается сложным процессам брожения, которые увеличивают ее усвояемость. В этом отделе в огромных количествах живут разнообразные микроорганизмы, вызывающие процессы брожения, которые протекают в условиях нейтральной или слабощелочной реакции. В рубце наряду с бактериями обитает масса простейших — очень мелких жгутиконосцев и инфузорий, так как здесь создаются весьма благоприятные условия для их развития.Из рубца через сетку пища отрыгивается в ротовую полость, где дополнительно пережевывается (жвачка). Проглатываемая вновь пищевая пережеванная масса по особой трубке, образуемой складками пищевода, идет уже не в рубец, а в книжку и оттуда в сычуг, где подвергается действию пищеварительных соков жвачного. В сычуге в условиях кислой реакции и наличия пищеварительных ферментов инфузории погибают. Попадая туда со жвачкой, они перевариваются.Количество простейших в рубце (а также в сетке) может достигать колоссальных величин. Если взять каплю содержимого рубца и рассмотреть ее под микроскопом (при нагревании, так как при комнатной температуре инфузории останавливаются), то в поле зрения инфузории буквально кишат. Трудно даже в условиях культуры получить такую массу инфузорий. Количество инфузорий в 1 см3 содержимого рубца достигает миллиона, а нередко и более. В пересчете на весь объем рубца это дает поистине астрономические цифры! Богатство содержимого рубца инфузориями в большой степени зависит от характера пищи жвачного. Если пища богата клетчаткой и бедна углеводами и белками (трава, солома), то инфузорий в рубце относительно немного. При добавлении в пищевой рацион углеводов и белков (отруби) количество инфузорий резко увеличивается и достигает огромных цифр. Нужно иметь в виду, что существует постоянный отток инфузорий. Попадая вместе с жвачкой в сычуг, они погибают. Высокий уровень количества инфузорий поддерживается энергичным размножением их.У непарнокопытных (лошадь, осел, зебра) в пищевом тракте тоже имеется большое количество инфузорий, однако локализация их в хозяине иная. У непарнокопытных нет сложного желудка, благодаря чему возможность развития простейших в передних отделах пищевого тракта отсутствует. Зато у непарнокопытных очень сильно развиты толстая и слепая кишка, которые обычно забиты пищевыми массами и играют существенную роль в пищеварении. В этом отделе кишечника, подобно тому как в рубце и сетке жвачных, развивается очень богатая фауна простейших, преимущественно инфузорий, большинство которых также относится к отряду энтодиниоморф. Однако по видовому составу фауна рубца жвачных и фауна толстого кишечника непарнокопытных не совпадают.ИНФУЗОРИИ КИШЕЧНОГО ТРАКТА ЖВАЧНЫХНаибольший интерес представляют инфузории семейства офриосколецид (Ophryoscolecidae), относящегося к отряду энтодиниоморф. Характерный признак этого отряда — отсутствие сплошного ресничного покрова. Сложные ресничные образования — цирры — расположены на переднем конце тела инфузорий в области ротового отверстия. К этим основным элементам ресничного аппарата могут прибавиться еще дополнительные группы цирр, расположенные либо на переднем, либо на заднем конце тела. Общее количество видов инфузорий семейства офриосколецид — около 120.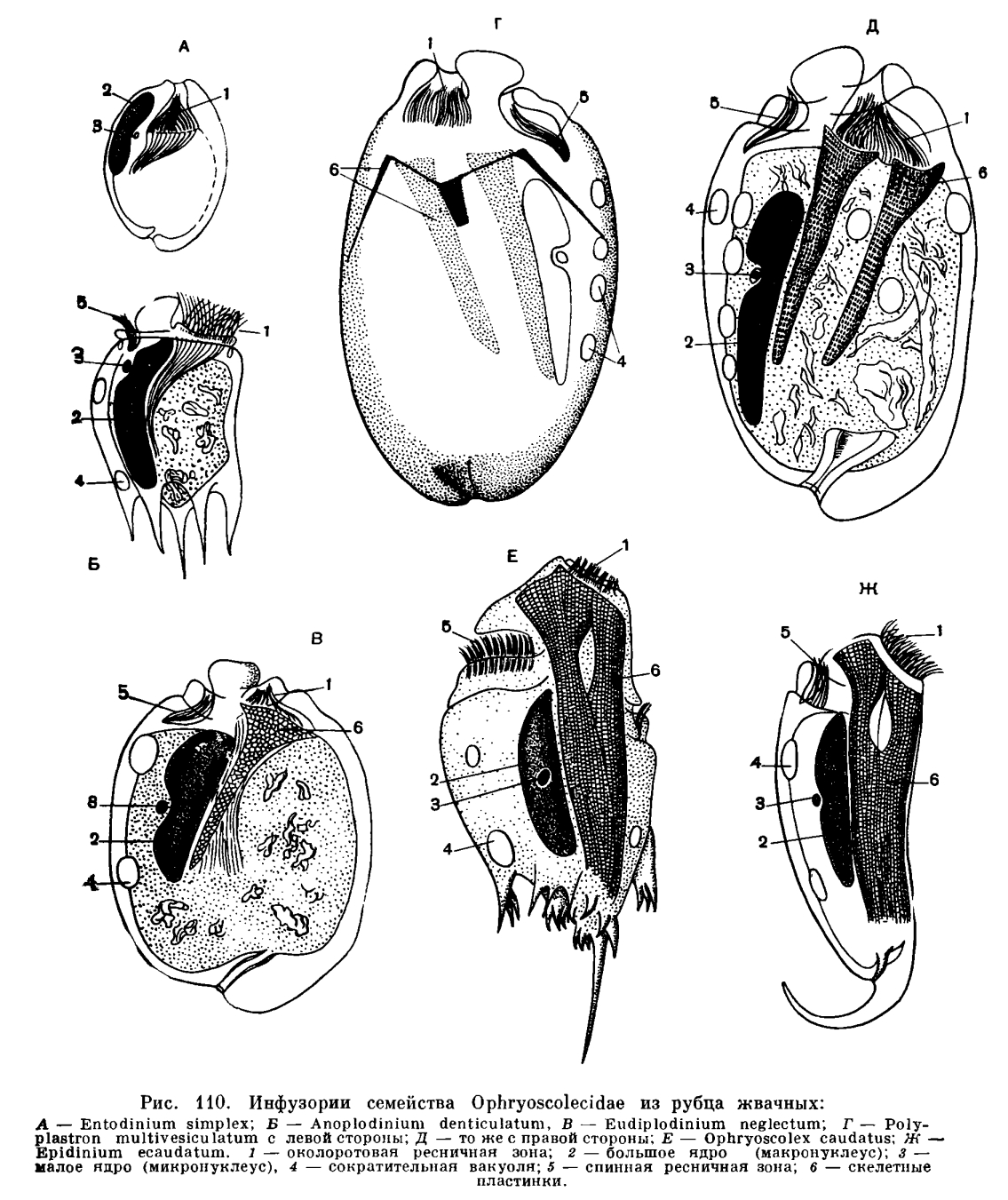 На рисунке 110 изображены некоторые наиболее типичные представители офриосколецид из рубца жвачных. Наиболее просто устроены инфузории рода энтодиний (Entodinium, рис. 110, Л). На переднем конце тела их имеется одна околоротовая зона цирр. Передний конец тела, на котором расположено ротовое отверстие, может втягиваться внутрь. Резко разграничены эктоплазма и эндоплазма. Хорошо видна на заднем конце анальная трубка, служащая для выведения наружу непереваренных остатков пищи. Несколько сложнее строение аноплодиния (Anoplodinium, рис. 110, Б). У них имеются две зоны ресничного аппарата — околоротовые цирры и спинные цирры. Обе они расположены на переднем конце. На заднем конце тела изображенного на рисунке вида имеются длинные острые выросты — это довольно типично для многих видов офриосколецид. Высказывалось предположение, что эти выросты способствуют «проталкиванию» инфузорий между растительными частицами, заполняющими рубец.Виды рода эудиплодиний (Eudiplodinium, рис. 110, Б) сходны с аноплодинием, но, в отлпчие от них, имеют скелетную опорную пластинку, расположенную но правому краю вдоль глотки. Эта скелетная пластинка состоит из вещества, близкого по химической природе к клетчатке, т. е. к веществу, из которого состоят оболочки растительных клеток.У рода полипластрон (Polyplastron, рис. 110, Г, Д) наблюдается дальнейшее усложнение скелета. Строение этих инфузорий близко к эудиплодиниям. Отличия сводятся прежде всего к тому, что вместо одной скелетной пластинки эти инфузории имеют пять. Две из них, наиболее крупные, расположены по правой стороне, а три, более мелкие, — по левой стороне инфузории. Вторая особенность полипластрона — увеличение количества сократительных вакуолей. У энтодиниев одна сократительная вакуоля, у аноплодиниев и эудиплодиниев две сократительные вакуоли, у полипластрона их около десятка.У эпидиниев (Epidinium, рис. 110), обладающих хорошо развитым углеводным скелетом, расположенным по правой стороне тела, спинная зона цирр смещается с переднего конца на спинную сторону. На заднем конце инфузорий этого рода часто развиваются шипы.Наиболее сложное строение обнаруживает род офриосколекс (Ophryoscolex), по которому названо и все семейство инфузорий (рис. 110, Е). У них хорошо развита спинная зона цирр, охватывающая около 2/3 окружности тела и скелетные пластинки. На заднем конце образуются многочисленные шипы, из которых один бывает обычно особенно длинным.Знакомство с некоторыми типичными представителями офриосколецид показывает, что в пределах этого семейства произошло значительное усложнение организации (от энтодиниев до офриосколекса).Очень часто, как мы это увидим, например, при знакомстве с червями, паразитизм, особенно эндопаразитизм, приводит к значительному упрощению организации, к регрессивным изменениям многих систем органов. У инфузорий, живущих в кишечнике копытных, этого не произошло. Напротив, здесь имело место прогрессивное развитие, не упрощение, а усложнение организации. Это связано, вероятно, с особенностями среды обитания инфузорий рубца. Они живут как бы в своеобразном водоеме с очень сложными условиями (наличие многочисленных механических препятствий, сложный бактериальный состав среды и многое другое), приспособление к которым было связано с усложнением строения инфузорий.Кроме инфузорий семейства офриосколецид, в рубце жвачных встречаются в небольших количествах представители уже известного нам отряда равноресничных инфузорий. Они представлены небольшим числом видов. Их тело равномерно покрыто продольными рядами ресничек, скелетные элементы отсутствуют. В общей массе инфузорного населения рубца они не играют заметной роли, и поэтому мы не будем здесь останавливаться на их рассмотрении.Чем и как питаются инфузории офриосколециды? Этот вопрос подробно изучался многими учеными, особенно детально профессором В. А. Догелем.
На рисунке 110 изображены некоторые наиболее типичные представители офриосколецид из рубца жвачных. Наиболее просто устроены инфузории рода энтодиний (Entodinium, рис. 110, Л). На переднем конце тела их имеется одна околоротовая зона цирр. Передний конец тела, на котором расположено ротовое отверстие, может втягиваться внутрь. Резко разграничены эктоплазма и эндоплазма. Хорошо видна на заднем конце анальная трубка, служащая для выведения наружу непереваренных остатков пищи. Несколько сложнее строение аноплодиния (Anoplodinium, рис. 110, Б). У них имеются две зоны ресничного аппарата — околоротовые цирры и спинные цирры. Обе они расположены на переднем конце. На заднем конце тела изображенного на рисунке вида имеются длинные острые выросты — это довольно типично для многих видов офриосколецид. Высказывалось предположение, что эти выросты способствуют «проталкиванию» инфузорий между растительными частицами, заполняющими рубец.Виды рода эудиплодиний (Eudiplodinium, рис. 110, Б) сходны с аноплодинием, но, в отлпчие от них, имеют скелетную опорную пластинку, расположенную но правому краю вдоль глотки. Эта скелетная пластинка состоит из вещества, близкого по химической природе к клетчатке, т. е. к веществу, из которого состоят оболочки растительных клеток.У рода полипластрон (Polyplastron, рис. 110, Г, Д) наблюдается дальнейшее усложнение скелета. Строение этих инфузорий близко к эудиплодиниям. Отличия сводятся прежде всего к тому, что вместо одной скелетной пластинки эти инфузории имеют пять. Две из них, наиболее крупные, расположены по правой стороне, а три, более мелкие, — по левой стороне инфузории. Вторая особенность полипластрона — увеличение количества сократительных вакуолей. У энтодиниев одна сократительная вакуоля, у аноплодиниев и эудиплодиниев две сократительные вакуоли, у полипластрона их около десятка.У эпидиниев (Epidinium, рис. 110), обладающих хорошо развитым углеводным скелетом, расположенным по правой стороне тела, спинная зона цирр смещается с переднего конца на спинную сторону. На заднем конце инфузорий этого рода часто развиваются шипы.Наиболее сложное строение обнаруживает род офриосколекс (Ophryoscolex), по которому названо и все семейство инфузорий (рис. 110, Е). У них хорошо развита спинная зона цирр, охватывающая около 2/3 окружности тела и скелетные пластинки. На заднем конце образуются многочисленные шипы, из которых один бывает обычно особенно длинным.Знакомство с некоторыми типичными представителями офриосколецид показывает, что в пределах этого семейства произошло значительное усложнение организации (от энтодиниев до офриосколекса).Очень часто, как мы это увидим, например, при знакомстве с червями, паразитизм, особенно эндопаразитизм, приводит к значительному упрощению организации, к регрессивным изменениям многих систем органов. У инфузорий, живущих в кишечнике копытных, этого не произошло. Напротив, здесь имело место прогрессивное развитие, не упрощение, а усложнение организации. Это связано, вероятно, с особенностями среды обитания инфузорий рубца. Они живут как бы в своеобразном водоеме с очень сложными условиями (наличие многочисленных механических препятствий, сложный бактериальный состав среды и многое другое), приспособление к которым было связано с усложнением строения инфузорий.Кроме инфузорий семейства офриосколецид, в рубце жвачных встречаются в небольших количествах представители уже известного нам отряда равноресничных инфузорий. Они представлены небольшим числом видов. Их тело равномерно покрыто продольными рядами ресничек, скелетные элементы отсутствуют. В общей массе инфузорного населения рубца они не играют заметной роли, и поэтому мы не будем здесь останавливаться на их рассмотрении.Чем и как питаются инфузории офриосколециды? Этот вопрос подробно изучался многими учеными, особенно детально профессором В. А. Догелем.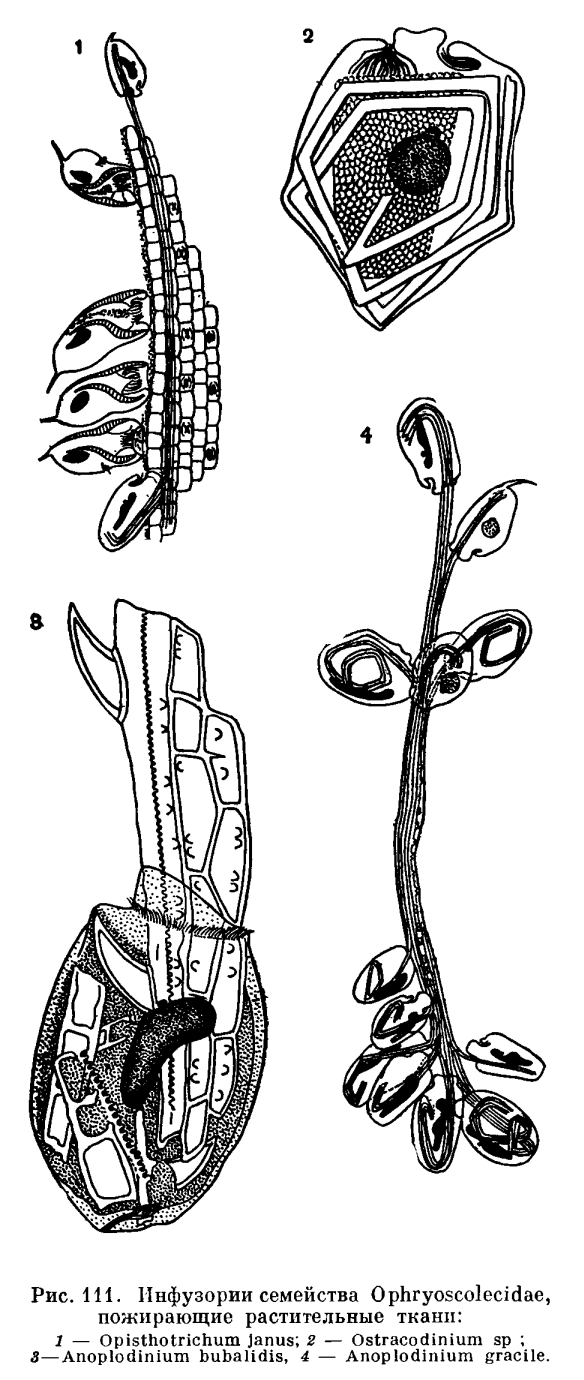 Пища офриосколецид довольно разнообразна, и у разных видов их наблюдается известная специализация. Самые мелкие виды рода энтодиниев питаются бактериями, зернами крахмала, грибками и другими небольшими частицами. Очень многие средние и крупные офриосколециды поглощают частицы растительных тканей, которые составляют главную массу содержимого рубца. Эндоплазма некоторых видов бывает буквально забита растительными частицами. Можно видеть, как инфузории набрасываются на обрывки растительных тканей, буквально разрывают их на кусочки и затем заглатывают, нередко закручивая их в своем теле спиралью (рис. 111, 4). Иногда приходится наблюдать такие картины (рис. 111, 2), когда само тело инфузории оказывается деформированным благодаря заглоченным крупным частицам.У офриосколецид наблюдается иногда хищничество. Более крупные виды пожирают более мелкие. Хищничество (рис. 112) сочетается со способностью тех же видов питаться растительными частицами.
Пища офриосколецид довольно разнообразна, и у разных видов их наблюдается известная специализация. Самые мелкие виды рода энтодиниев питаются бактериями, зернами крахмала, грибками и другими небольшими частицами. Очень многие средние и крупные офриосколециды поглощают частицы растительных тканей, которые составляют главную массу содержимого рубца. Эндоплазма некоторых видов бывает буквально забита растительными частицами. Можно видеть, как инфузории набрасываются на обрывки растительных тканей, буквально разрывают их на кусочки и затем заглатывают, нередко закручивая их в своем теле спиралью (рис. 111, 4). Иногда приходится наблюдать такие картины (рис. 111, 2), когда само тело инфузории оказывается деформированным благодаря заглоченным крупным частицам.У офриосколецид наблюдается иногда хищничество. Более крупные виды пожирают более мелкие. Хищничество (рис. 112) сочетается со способностью тех же видов питаться растительными частицами. Каким путем инфузории проникают в рубец жвачного? Каковы пути заражения офриосколецидами? Оказывается, что новорожденные жвачные не имеют еще в рубце инфузорий. Они отсутствуют также, пока животное питается молоком. Но как только жвачное переходит на растительную пищу, сразу же в рубце и сетке появляются инфузории, количество которых быстро нарастает. Откуда они берутся? Долгое время предполагали, что инфузории рубца образуют какие-то покоящиеся стадии (вероятнее всего, цисты), которые широко рассеиваются в природе и, будучи проглоченными, дают начало активным стадиям инфузорий. Дальнейшие исследования показали, что у инфузорий рубца жвачных никаких покоящихся стадий нет. Удалось доказать, что заражение происходит активными подвижными инфузориями, которые проникают в ротовую полость при отрыгивании жвачки. Если исследовать под микроскопом взятую из ротовой полости жвачку, го в ней всегда имеется большое количество активно плавающих инфузорий. Эти активные формы легко могут проникнуть в рот и далее в рубец других особей жвачного из общего сосуда для пойла, вместе с травой, сеном (на которых может оказаться слюна с инфузориями) и т. п. Этот путь заражения был доказан экспериментально.Если покоящиеся стадии у офриосколецид отсутствуют, то, очевидно, легко получить «безинфузорных» животных, изолировав их, когда они еще питаются молоком. Если не допускать прямого контакта между растущим молодняком и имеющими инфузорий жвачными, то молодые животные могут остаться без инфузорий в рубце. Такие опыты были проведены несколькими учеными в разных странах. Результат был однозначен. При отсутствии контакта между молодняком (отнятым от матери еще в период кормления молоком) и имеющими в рубце инфузорий жвачными, животные вырастают стерильными в отношении инфузорий. Однако достаточно даже кратковременного контакта с животными, имеющими инфузорий (общая кормушка, общее ведро для питья, общее пастбище), чтобы в рубце стерильных животных появилась фауна инфузорий.Массовое развитие фауны простейших в рубце и сетке, естественно, ставит вопрос о характере взаимоотношений между простейшими и их хозяевами — жвачными. Мы уже видели на примере жгутиконосцев, паразитирующих в кишечнике термитов, какие тесные симбиотические взаимоотношения могут возникать между простейшими, населяющими кишечный тракт, и их хозяевами. Не существует ли между инфузориями, населяющими рубец и сетку, аналогичных симбиотических взаимоотношений? Этот вопрос ставился многими исследователями. Однако до сих пор окончательный ответ на него не получен, и требуются дальнейшие исследования. Ввиду большого биологического и практического интереса этого вопроса рассмотрим его несколько подробнее.Выше были приведены результаты опытов по содержанию жвачных, совершенно лишенных инфузорий в рубце и сетке. Это достигается, как мы видели, путем ранней изоляции молодняка. Опыты проводились на овцах и на козах.Таким путем удавалось проводить наблюдения за «безинфузорными» животными в течение значительного промежутка времени (свыше года). Как отражается отсутствие инфузорий в рубце на жизни хозяина? Отрицательно или положительно влияет отсутствие инфузорий на хозяина? Для ответа на этот вопрос на козах провели следующие опыты. Были взяты козлята-близнецы (одного помета и одного пола), чтобы иметь более сходный материал. Затем один из близнецов данной пары воспитывался без инфузорий в рубце (ранняя изоляция), другой же с самого начала питания растительной пищей обильно заражался множеством видов инфузорий. Оба получали совершенно одинаковый рацион и воспитывались в одинаковых условиях. Единственное различие между ними сводилось к наличию или отсутствию инфузорий. На нескольких изученных таким образом парах козлят не было обнаружено каких-либо различий в ходе развития обоих членов каждой пары («инфузорного» и «безинфузорного»). Таким образом, можно утверждать, что сколько-нибудь резкого влияния на жизненные функции животного-хозяина инфузории, живущие в рубце и сетке, не оказывают.Приведенные выше результаты опытов не позволяют, однако, утверждать, что инфузории рубца совершенно безразличны для хозяина. Эти опыты проводились при нормальном пищевом рационе хозяина. Не исключена возможность, что при других условиях, при ином пищевом режиме (например, при недостаточном кормлении) удастся выявить влияние на хозяина фауны простейших, населяющих рубец.В литературе высказывались различные предположения о возможном положительном влиянии протозойной фауны рубца на пищеварительные процессы хозяина. Указывалось, что многие миллионы инфузорий, активно плавающие в рубце и размельчающие растительные ткани, способствуют брожению и перевариванию пищевых масс, находящихся в передних отделах пищеварительного тракта. Значительное количество инфузорий, попадающее вместе с жвачкой в сычуг, переваривается, и белок, составляющий значительную часть тела инфузорий, усваивается. Инфузории, таким образом, могут быть дополнительным источником белка для хозяина. Высказывались также предположения, что инфузории способствуют перевариванию клетчатки, составляющей основную массу пищи жвачных, переводу ее в более усвояемое состояние.Все эти предположения не являются доказанными, и против некоторых из них выдвигались возражения. Указывалось, например, что инфузории строят протоплазму своего тела из белков, которые поступают в рубец с пищей хозяина. Поглощая белок растительный, онп переводят его в животный белок своего тела, который затем переваривается в сычуге. Дает ли это какие-нибудь преимущества хозяину, остается неясно. Все эти вопросы имеют большой практический интерес, так как речь идет о пищеварении жвачных — основных объектов животноводства. Дальнейшие исследования о роли инфузорий рубца в пищеварении жвачных очень желательны.Насколько строго приурочены определенные виды офриосколецид к определенным видам хозяев? Имеются ли, например, в рогатом скоте свои виды инфузорий, в овцах — свои и т. д.? Приспособленность паразита к определенному кругу хозяев называют его специфичностью. Если паразит строго приурочен к одному виду хозяев, это говорит об узкой специфичности. Если данный вид паразита может жить в разных видах хозяев, то это проявление широкой специфичности.Офриосколециды жвачных обладают, как правило, широкой специфичностью. В видовом отношении население рубца и сетки рогатого скота, овец, коз очень близко между собой. Если сравнить видовой состав рубца африканских антилоп с рогатым скотом, то и здесь около 40% от общего числа видов окажется общим. Однако существует немало видов офриосколецид, которые встречаются только в антилопах или только в оленях. Таким образом, на фоне общей широкой специфичности офриосколецид можно говорить об отдельных более узко специфичных видах их.ИНФУЗОРИИ КИШЕЧНИКА НЕПАРНОКОПЫТНЫХОбратимся теперь к краткому знакомству с инфузориями, населяющими толстую и слепую кишку непарнокопытных.В видовом отношении эта фауна, как и фауна рубца жвачных, тоже весьма разнообразна. В настоящее время описано около 100 видов инфузорий, обитающих в толстом кишечнике животных семейства лошадиных. Встречающиеся здесь инфузории в смысле их принадлежности к разным систематическим группам более разнообразны, чем инфузории рубца жвачных.
Каким путем инфузории проникают в рубец жвачного? Каковы пути заражения офриосколецидами? Оказывается, что новорожденные жвачные не имеют еще в рубце инфузорий. Они отсутствуют также, пока животное питается молоком. Но как только жвачное переходит на растительную пищу, сразу же в рубце и сетке появляются инфузории, количество которых быстро нарастает. Откуда они берутся? Долгое время предполагали, что инфузории рубца образуют какие-то покоящиеся стадии (вероятнее всего, цисты), которые широко рассеиваются в природе и, будучи проглоченными, дают начало активным стадиям инфузорий. Дальнейшие исследования показали, что у инфузорий рубца жвачных никаких покоящихся стадий нет. Удалось доказать, что заражение происходит активными подвижными инфузориями, которые проникают в ротовую полость при отрыгивании жвачки. Если исследовать под микроскопом взятую из ротовой полости жвачку, го в ней всегда имеется большое количество активно плавающих инфузорий. Эти активные формы легко могут проникнуть в рот и далее в рубец других особей жвачного из общего сосуда для пойла, вместе с травой, сеном (на которых может оказаться слюна с инфузориями) и т. п. Этот путь заражения был доказан экспериментально.Если покоящиеся стадии у офриосколецид отсутствуют, то, очевидно, легко получить «безинфузорных» животных, изолировав их, когда они еще питаются молоком. Если не допускать прямого контакта между растущим молодняком и имеющими инфузорий жвачными, то молодые животные могут остаться без инфузорий в рубце. Такие опыты были проведены несколькими учеными в разных странах. Результат был однозначен. При отсутствии контакта между молодняком (отнятым от матери еще в период кормления молоком) и имеющими в рубце инфузорий жвачными, животные вырастают стерильными в отношении инфузорий. Однако достаточно даже кратковременного контакта с животными, имеющими инфузорий (общая кормушка, общее ведро для питья, общее пастбище), чтобы в рубце стерильных животных появилась фауна инфузорий.Массовое развитие фауны простейших в рубце и сетке, естественно, ставит вопрос о характере взаимоотношений между простейшими и их хозяевами — жвачными. Мы уже видели на примере жгутиконосцев, паразитирующих в кишечнике термитов, какие тесные симбиотические взаимоотношения могут возникать между простейшими, населяющими кишечный тракт, и их хозяевами. Не существует ли между инфузориями, населяющими рубец и сетку, аналогичных симбиотических взаимоотношений? Этот вопрос ставился многими исследователями. Однако до сих пор окончательный ответ на него не получен, и требуются дальнейшие исследования. Ввиду большого биологического и практического интереса этого вопроса рассмотрим его несколько подробнее.Выше были приведены результаты опытов по содержанию жвачных, совершенно лишенных инфузорий в рубце и сетке. Это достигается, как мы видели, путем ранней изоляции молодняка. Опыты проводились на овцах и на козах.Таким путем удавалось проводить наблюдения за «безинфузорными» животными в течение значительного промежутка времени (свыше года). Как отражается отсутствие инфузорий в рубце на жизни хозяина? Отрицательно или положительно влияет отсутствие инфузорий на хозяина? Для ответа на этот вопрос на козах провели следующие опыты. Были взяты козлята-близнецы (одного помета и одного пола), чтобы иметь более сходный материал. Затем один из близнецов данной пары воспитывался без инфузорий в рубце (ранняя изоляция), другой же с самого начала питания растительной пищей обильно заражался множеством видов инфузорий. Оба получали совершенно одинаковый рацион и воспитывались в одинаковых условиях. Единственное различие между ними сводилось к наличию или отсутствию инфузорий. На нескольких изученных таким образом парах козлят не было обнаружено каких-либо различий в ходе развития обоих членов каждой пары («инфузорного» и «безинфузорного»). Таким образом, можно утверждать, что сколько-нибудь резкого влияния на жизненные функции животного-хозяина инфузории, живущие в рубце и сетке, не оказывают.Приведенные выше результаты опытов не позволяют, однако, утверждать, что инфузории рубца совершенно безразличны для хозяина. Эти опыты проводились при нормальном пищевом рационе хозяина. Не исключена возможность, что при других условиях, при ином пищевом режиме (например, при недостаточном кормлении) удастся выявить влияние на хозяина фауны простейших, населяющих рубец.В литературе высказывались различные предположения о возможном положительном влиянии протозойной фауны рубца на пищеварительные процессы хозяина. Указывалось, что многие миллионы инфузорий, активно плавающие в рубце и размельчающие растительные ткани, способствуют брожению и перевариванию пищевых масс, находящихся в передних отделах пищеварительного тракта. Значительное количество инфузорий, попадающее вместе с жвачкой в сычуг, переваривается, и белок, составляющий значительную часть тела инфузорий, усваивается. Инфузории, таким образом, могут быть дополнительным источником белка для хозяина. Высказывались также предположения, что инфузории способствуют перевариванию клетчатки, составляющей основную массу пищи жвачных, переводу ее в более усвояемое состояние.Все эти предположения не являются доказанными, и против некоторых из них выдвигались возражения. Указывалось, например, что инфузории строят протоплазму своего тела из белков, которые поступают в рубец с пищей хозяина. Поглощая белок растительный, онп переводят его в животный белок своего тела, который затем переваривается в сычуге. Дает ли это какие-нибудь преимущества хозяину, остается неясно. Все эти вопросы имеют большой практический интерес, так как речь идет о пищеварении жвачных — основных объектов животноводства. Дальнейшие исследования о роли инфузорий рубца в пищеварении жвачных очень желательны.Насколько строго приурочены определенные виды офриосколецид к определенным видам хозяев? Имеются ли, например, в рогатом скоте свои виды инфузорий, в овцах — свои и т. д.? Приспособленность паразита к определенному кругу хозяев называют его специфичностью. Если паразит строго приурочен к одному виду хозяев, это говорит об узкой специфичности. Если данный вид паразита может жить в разных видах хозяев, то это проявление широкой специфичности.Офриосколециды жвачных обладают, как правило, широкой специфичностью. В видовом отношении население рубца и сетки рогатого скота, овец, коз очень близко между собой. Если сравнить видовой состав рубца африканских антилоп с рогатым скотом, то и здесь около 40% от общего числа видов окажется общим. Однако существует немало видов офриосколецид, которые встречаются только в антилопах или только в оленях. Таким образом, на фоне общей широкой специфичности офриосколецид можно говорить об отдельных более узко специфичных видах их.ИНФУЗОРИИ КИШЕЧНИКА НЕПАРНОКОПЫТНЫХОбратимся теперь к краткому знакомству с инфузориями, населяющими толстую и слепую кишку непарнокопытных.В видовом отношении эта фауна, как и фауна рубца жвачных, тоже весьма разнообразна. В настоящее время описано около 100 видов инфузорий, обитающих в толстом кишечнике животных семейства лошадиных. Встречающиеся здесь инфузории в смысле их принадлежности к разным систематическим группам более разнообразны, чем инфузории рубца жвачных. В кишечнике лошадиных обитает довольно много видов инфузорий, относящихся к отряду равноресничных, т. е. инфузорий, у которых ресничный аппарат не образует около ротовой зоны мембранелл или цирр (рис. 113, 1).Отряд энтодиниоморф (Entodiniomorpha) тоже богато представлен в кишечнике лошади. В то время как в рубце жвачных встречается лишь одно семейство энтодиниоморф (семейство офриосколецид), в кишечнике лошади живут представители трех семейств, на характеристике которых мы, однако, останавливаться здесь не будем, ограничившись лишь несколькими рисунками типичных для лошади видов (рис. 113).Детальными исследованиями А.Стрелкова было показано, что разные виды инфузорий далеко не равномерно распределяются по ходу толстого кишечника лошади. Имеются две разные группы видов, как бы две фауны. Одна из них населяет слепую кишку и брюшной отдел большой ободочной кишки (начальные отделы толстого кишечника), а другая—спинной отдел большой ободочной и малую ободочную. Эти два комплекса видов довольно резко разграничены. Видов, общих для этих двух разделов, немного — менее десятка.
В кишечнике лошадиных обитает довольно много видов инфузорий, относящихся к отряду равноресничных, т. е. инфузорий, у которых ресничный аппарат не образует около ротовой зоны мембранелл или цирр (рис. 113, 1).Отряд энтодиниоморф (Entodiniomorpha) тоже богато представлен в кишечнике лошади. В то время как в рубце жвачных встречается лишь одно семейство энтодиниоморф (семейство офриосколецид), в кишечнике лошади живут представители трех семейств, на характеристике которых мы, однако, останавливаться здесь не будем, ограничившись лишь несколькими рисунками типичных для лошади видов (рис. 113).Детальными исследованиями А.Стрелкова было показано, что разные виды инфузорий далеко не равномерно распределяются по ходу толстого кишечника лошади. Имеются две разные группы видов, как бы две фауны. Одна из них населяет слепую кишку и брюшной отдел большой ободочной кишки (начальные отделы толстого кишечника), а другая—спинной отдел большой ободочной и малую ободочную. Эти два комплекса видов довольно резко разграничены. Видов, общих для этих двух разделов, немного — менее десятка.
,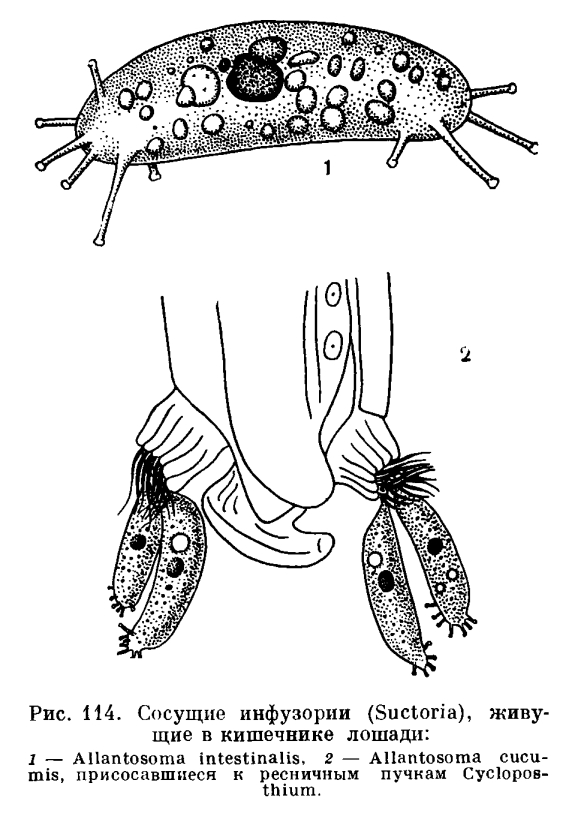 Интересно отметить, что среди многочисленных видов инфузорий, населяющих толстый кишечник непарнокопытных, имеются представители одного рода, относящегося к сосущим инфузориям. Как мы видели выше, сосущие инфузории (Suctoria) — это типичные свободноживущие сидячие организмы с совершенно особым способом питания при помощи щупалец (рис. 103). Один из родов сукторий приспособился к такой, казалось бы, необычной среде обитания, как толстый кишечник лошади, например несколько видов аллантозом (Allantosoma). Эти очень своеобразные животные (рис. 114) не имеют стебелька, реснички отсутствуют, булавовидные, утолщенные на концах щупальца хорошо развиты.При помощи щупалец аллантозомы присасываются к разным видам инфузорий и высасывают их. Нередко при этом добыча во много раз превосходит хищника.Неясен еще вопрос о характере взаимоотношений инфузорий толстого кишечника непарнокопытных с хозяевами. Количество инфузорий может быть столь же велико, а иногда даже больше, чем в рубце жвачных. Имеются данные, показывающие, что количество инфузории в толстых кишках лошади может достигать 3 млн. в 1 см3. Симбиотическое значение, предполагавшееся некоторыми учеными, еще менее вероятно, чем для инфузорий рубца.Наиболее вероятно мнение, что они наносят некоторый вред хозяину, поглощая значительное количество пищи. Часть инфузорий выносится наружу с фекальными массами, и таким образом органические вещества (в том числе и белок), входящие в состав их тела, остаются не использованными хозяином.Вопрос о путях заражения непарнокопытных инфузориями, населяющими толстый кишечник, до сих пор не разрешен.БАЛАНТИДИЙ-ПАРАЗИТ КИШЕЧНИКА ЧЕЛОВЕКАОдин вид паразитических инфузорий из отряда равноресничных — балантидий коли (Balantidium coli) — паразитирует в толстом кишечнике человека. Случаи заражения человека балантидиями очень редки. В этом отношении балантидий несравним с кишечными амебами, которые, как мы видели, встречаются часто. Паразитирование балантидия в кишечнике человека обычно вызывает тяжело протекающий колит. Поэтому паразит этот требует к себе пристального внимания, несмотря на то что встречается он относительно редко.
Интересно отметить, что среди многочисленных видов инфузорий, населяющих толстый кишечник непарнокопытных, имеются представители одного рода, относящегося к сосущим инфузориям. Как мы видели выше, сосущие инфузории (Suctoria) — это типичные свободноживущие сидячие организмы с совершенно особым способом питания при помощи щупалец (рис. 103). Один из родов сукторий приспособился к такой, казалось бы, необычной среде обитания, как толстый кишечник лошади, например несколько видов аллантозом (Allantosoma). Эти очень своеобразные животные (рис. 114) не имеют стебелька, реснички отсутствуют, булавовидные, утолщенные на концах щупальца хорошо развиты.При помощи щупалец аллантозомы присасываются к разным видам инфузорий и высасывают их. Нередко при этом добыча во много раз превосходит хищника.Неясен еще вопрос о характере взаимоотношений инфузорий толстого кишечника непарнокопытных с хозяевами. Количество инфузорий может быть столь же велико, а иногда даже больше, чем в рубце жвачных. Имеются данные, показывающие, что количество инфузории в толстых кишках лошади может достигать 3 млн. в 1 см3. Симбиотическое значение, предполагавшееся некоторыми учеными, еще менее вероятно, чем для инфузорий рубца.Наиболее вероятно мнение, что они наносят некоторый вред хозяину, поглощая значительное количество пищи. Часть инфузорий выносится наружу с фекальными массами, и таким образом органические вещества (в том числе и белок), входящие в состав их тела, остаются не использованными хозяином.Вопрос о путях заражения непарнокопытных инфузориями, населяющими толстый кишечник, до сих пор не разрешен.БАЛАНТИДИЙ-ПАРАЗИТ КИШЕЧНИКА ЧЕЛОВЕКАОдин вид паразитических инфузорий из отряда равноресничных — балантидий коли (Balantidium coli) — паразитирует в толстом кишечнике человека. Случаи заражения человека балантидиями очень редки. В этом отношении балантидий несравним с кишечными амебами, которые, как мы видели, встречаются часто. Паразитирование балантидия в кишечнике человека обычно вызывает тяжело протекающий колит. Поэтому паразит этот требует к себе пристального внимания, несмотря на то что встречается он относительно редко.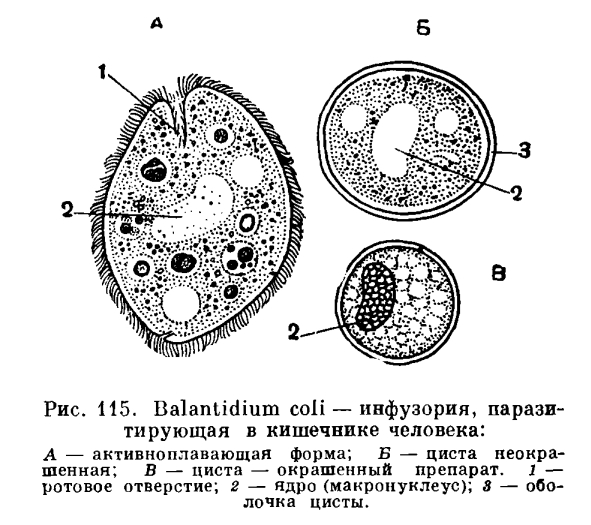 Тело балантидия, паразитирующего в кишечнике человека, имеет яйцевидную форму (рис. 115). Размеры варьируют в очень широких пределах: длина 30—150, ширина 20—110 мк. Эти вариации величины в значительной степени зависят от того, в какой мере инфузория набита пищей. Все тело густо покрыто короткими ресничками, расположенными продольными рядами от переднего конца к заднему. На переднем конце имеется углубление в виде неглубокой щели. Это околоротовое углубление (перистом), на дне которого помещается ротовое отверстие. По краю перистома расположены особые сильно развитые реснички, направляющие пищу в глубину его. Имеются две сократительные вакуоли (одна в передней трети тела, вторая на заднем конце). Ядерный аппарат типичный для инфузорий: бобовидный, расположенный примерно в центре тела макронуклеус и в непосредственном соседстве с ним один маленький микронуклеус.Балантидий захватывает разнообразные пищевые частицы из содержимого толстой кишки. Особенно охотно он питается крахмальными зернами. Если балантидий живет в просвете толстой кишки человека, то он питается содержимым кишечника и не оказывает никакого вредного влияния. Это типичное «носительство», с которым мы познакомились уже при рассмотрении дизентерийной амебы. Однако балантидий реже, чем дизентерийная амеба, остается таким «безобидным квартирантом».Он активно внедряется в слизистую кишечника и проникает в толщу его стенки. Здесь балантидий переключается на питание клетками хозяина и особенно «охотно» заглатывает красные кровяные тельца (эритроциты). В результате нарушения целостности внутренней стенки кишечник изъязвляется, и это сопровождается тяжелой формой кровавого поноса (колита). Поскольку паразит внедряется в самую толщу тканей кишечника, лечение балантидиоза очень затруднительно.В настоящее время в медицине известны некоторые специфические лекарства, убивающие балантидия. Однако успех лечения не всегда бывает положительным именно в силу внутритканевой локализации паразита.Может ли человеческий балантидий паразитировать в других хозяевах, кроме человека? Оказывается, что эта паразитическая инфузория часто встречается в кишечнике свиней. Но, в отличие от человеческого, этот балантидий обычно не вызывает у свиней болезненных явлений и живет в кишечнике их как «безобидный квартирант». Основным хозяином балантидия следует считать свинью, тогда как человек заражается редко, причем почти всегда источником заражения являются свиньи.Каким же путем осуществляется заражение балантидием? У этой инфузории имеются покоящиеся стадии — цисты. Инцистирование происходит при проникновении паразита в заднюю кишку. Цисты выводятся наружу с фекальными массами. Они долгое время (свыше двух месяцев при комнатной температуре) сохраняют жизнеспособность. Если циста будет случайно проглочена свиньей или человеком, то она проходит в неизменном виде через желудок и тонкие кишки. Когда циста достигнет толстой кишки, оболочка ее разрушается и из нее выходит активная стадия балантидия.Из сказанного выше следует, что персонал, обслуживающий свиней, должен соблюдать элементарные гигиенические правила, а главное не приступать к еде, не вымыв предварительно руки. Помимо кишечника человека и свиней, балантидий коли может паразитировать (при непосредственном заражении) в кишечнике крыс и человекообразных обезьян. У обезьян, так же как и у человека, балантидий вызывают тяжелую форму колита.В настоящее время специалистами хорошо разработаны различные методы, позволяющие культивировать балантидиев в искусственной среде — вне организма хозяина.ИНФУЗОРИИ-ПАРАЗИТЫ ЧЕЛОВЕКООБРАЗНЫХ ОБЕЗЬЯНМы видели, что человеческий балантидий может паразитировать в обезьянах, вызывая колиты. Однако в человекообразных обезьянах, кроме того, встречаются очень интересные и сложно устроенные инфузории, которые в организме человека не паразитируют. Они относятся к отряду энтодиниоморф (Entodiniomorpha), т. е. к тому же отряду, который, как мы видели, богато представлен в кишечном тракте копытных млекопитающих.В человекообразных обезьянах (горилле и шимпанзе) встречаются виды рода троглодителла (Troglodytella), паразитирующие в толстой и слепой кишке (рис. 116). В природной обстановке в Африке как гориллы, так и шимпанзе заражены, по-видимому, поголовно. В условиях неволи, в зоологических садах, инфузории постепенно исчезают из организма обезьян.
Тело балантидия, паразитирующего в кишечнике человека, имеет яйцевидную форму (рис. 115). Размеры варьируют в очень широких пределах: длина 30—150, ширина 20—110 мк. Эти вариации величины в значительной степени зависят от того, в какой мере инфузория набита пищей. Все тело густо покрыто короткими ресничками, расположенными продольными рядами от переднего конца к заднему. На переднем конце имеется углубление в виде неглубокой щели. Это околоротовое углубление (перистом), на дне которого помещается ротовое отверстие. По краю перистома расположены особые сильно развитые реснички, направляющие пищу в глубину его. Имеются две сократительные вакуоли (одна в передней трети тела, вторая на заднем конце). Ядерный аппарат типичный для инфузорий: бобовидный, расположенный примерно в центре тела макронуклеус и в непосредственном соседстве с ним один маленький микронуклеус.Балантидий захватывает разнообразные пищевые частицы из содержимого толстой кишки. Особенно охотно он питается крахмальными зернами. Если балантидий живет в просвете толстой кишки человека, то он питается содержимым кишечника и не оказывает никакого вредного влияния. Это типичное «носительство», с которым мы познакомились уже при рассмотрении дизентерийной амебы. Однако балантидий реже, чем дизентерийная амеба, остается таким «безобидным квартирантом».Он активно внедряется в слизистую кишечника и проникает в толщу его стенки. Здесь балантидий переключается на питание клетками хозяина и особенно «охотно» заглатывает красные кровяные тельца (эритроциты). В результате нарушения целостности внутренней стенки кишечник изъязвляется, и это сопровождается тяжелой формой кровавого поноса (колита). Поскольку паразит внедряется в самую толщу тканей кишечника, лечение балантидиоза очень затруднительно.В настоящее время в медицине известны некоторые специфические лекарства, убивающие балантидия. Однако успех лечения не всегда бывает положительным именно в силу внутритканевой локализации паразита.Может ли человеческий балантидий паразитировать в других хозяевах, кроме человека? Оказывается, что эта паразитическая инфузория часто встречается в кишечнике свиней. Но, в отличие от человеческого, этот балантидий обычно не вызывает у свиней болезненных явлений и живет в кишечнике их как «безобидный квартирант». Основным хозяином балантидия следует считать свинью, тогда как человек заражается редко, причем почти всегда источником заражения являются свиньи.Каким же путем осуществляется заражение балантидием? У этой инфузории имеются покоящиеся стадии — цисты. Инцистирование происходит при проникновении паразита в заднюю кишку. Цисты выводятся наружу с фекальными массами. Они долгое время (свыше двух месяцев при комнатной температуре) сохраняют жизнеспособность. Если циста будет случайно проглочена свиньей или человеком, то она проходит в неизменном виде через желудок и тонкие кишки. Когда циста достигнет толстой кишки, оболочка ее разрушается и из нее выходит активная стадия балантидия.Из сказанного выше следует, что персонал, обслуживающий свиней, должен соблюдать элементарные гигиенические правила, а главное не приступать к еде, не вымыв предварительно руки. Помимо кишечника человека и свиней, балантидий коли может паразитировать (при непосредственном заражении) в кишечнике крыс и человекообразных обезьян. У обезьян, так же как и у человека, балантидий вызывают тяжелую форму колита.В настоящее время специалистами хорошо разработаны различные методы, позволяющие культивировать балантидиев в искусственной среде — вне организма хозяина.ИНФУЗОРИИ-ПАРАЗИТЫ ЧЕЛОВЕКООБРАЗНЫХ ОБЕЗЬЯНМы видели, что человеческий балантидий может паразитировать в обезьянах, вызывая колиты. Однако в человекообразных обезьянах, кроме того, встречаются очень интересные и сложно устроенные инфузории, которые в организме человека не паразитируют. Они относятся к отряду энтодиниоморф (Entodiniomorpha), т. е. к тому же отряду, который, как мы видели, богато представлен в кишечном тракте копытных млекопитающих.В человекообразных обезьянах (горилле и шимпанзе) встречаются виды рода троглодителла (Troglodytella), паразитирующие в толстой и слепой кишке (рис. 116). В природной обстановке в Африке как гориллы, так и шимпанзе заражены, по-видимому, поголовно. В условиях неволи, в зоологических садах, инфузории постепенно исчезают из организма обезьян. Как видно из рисунка, троглодптелла относится к числу сложно устроенных энтодиниоморф. У нее, кроме околоротовой зоны цирр (на переднем конце тела), имеются еще три зоны хорошо развитых цирр, кольцеобразно охватывающих тело инфузории. Троглодителлы обладают хорошо развитым, охватывающим почти весь передний конец тела скелетным аппаратом, состоящим из углеводов. Размеры этих своеобразных инфузорий довольно значительны. В длину они достигают 200—280 мк.БЕЗРОТЫЕ ИНФУЗОРИИ АСТОМАТЫНа предыдущих страницах было рассказано о различных паразитических инфузориях, хозяевами которых являются позвоночные животные. Наряду с этим большое количество видов паразитических инфузорий живет в самых различных беспозвоночных животных. Рассмотрим кратко наиболее интересных представителей их.Глубоко приспособленной к паразитизму и выработавшей ряд интересных приспособлений является обширная группа безротых инфузорий — астомат (Astomata). Самая характерная особенность этого отряда отражена в его названии — инфузории астоматы лишены ротового отверстия. Их питание осуществляется осмотическим путем — всасыванием питательных веществ всей поверхностью тела. Поскольку оформленной пищи астоматы не поглощают, пищеварительные вакуоли у них не образуются.Общее число видов астомат превышает сотню. Встречаются они в беспозвоночных, относящихся к самым различным группам (типам и классам) животного мира: в ресничных червях, кольчатых червях, моллюсках, иглокожих. Небольшое число видов астомат паразитирует и в позвоночных животных, а именно в земноводных. Однако астоматы распределены по разным группам хозяев совсем неравномерно. Существует одна группа «излюбленных» хозяев астомат, в которой сосредоточено свыше 75% их видов. Такой группой являются малощетинковые кольчатые черви (Oligochaeta), класс животных, к которому относится всем известный дождевой червь. Однако лишь часть видов малощетпнковых кольчецов живет в почве. Большая часть их — обитатели пресных вод, где они образуют заметную (а иногда и преобладающую) часть донного населения. Эти пресноводные малощетинковые кольчецы и являются «излюбленными» хозяевами астомат. Небольшое число видов олигохет встречается и в море, почти исключительно в прибрежной, литоральной (осушаемой во время отлива) зоне, но в морской фауне они не играют сколько-нибудь заметной роли.Органом, в котором локализуются в олигохетах инфузории астоматы, является кишечник. Поэтому многие черты строения астомат связаны с приспособлением к жизни в кишечнике. Через кишечник постоянно осуществляется движение пищи, иногда довольно активное. Паразиты кишечника, чтобы не быть вынесенными наружу (что равнозначно гибели), должны обладать приспособлениями, направленными на то, чтобы удержаться в кишечнике. Эти приспособления выражены не только у инфузорий, но и у всех других групп кишечных паразитов, к какой бы систематической группе они ни относились и в каких бы хозяевах ни паразитировали. Мы увидим это в дальнейшем, при знакомстве с разными группами паразитических червей.Тело всех паразитирующих в кишечнике олигохет астомат вытянуто в длину, что способствует их движению в просвете кишечника. Ресничный покров всегда хорошо развит, реснички покрывают все тело инфузорий и расположены тесными продольными рядами.
Как видно из рисунка, троглодптелла относится к числу сложно устроенных энтодиниоморф. У нее, кроме околоротовой зоны цирр (на переднем конце тела), имеются еще три зоны хорошо развитых цирр, кольцеобразно охватывающих тело инфузории. Троглодителлы обладают хорошо развитым, охватывающим почти весь передний конец тела скелетным аппаратом, состоящим из углеводов. Размеры этих своеобразных инфузорий довольно значительны. В длину они достигают 200—280 мк.БЕЗРОТЫЕ ИНФУЗОРИИ АСТОМАТЫНа предыдущих страницах было рассказано о различных паразитических инфузориях, хозяевами которых являются позвоночные животные. Наряду с этим большое количество видов паразитических инфузорий живет в самых различных беспозвоночных животных. Рассмотрим кратко наиболее интересных представителей их.Глубоко приспособленной к паразитизму и выработавшей ряд интересных приспособлений является обширная группа безротых инфузорий — астомат (Astomata). Самая характерная особенность этого отряда отражена в его названии — инфузории астоматы лишены ротового отверстия. Их питание осуществляется осмотическим путем — всасыванием питательных веществ всей поверхностью тела. Поскольку оформленной пищи астоматы не поглощают, пищеварительные вакуоли у них не образуются.Общее число видов астомат превышает сотню. Встречаются они в беспозвоночных, относящихся к самым различным группам (типам и классам) животного мира: в ресничных червях, кольчатых червях, моллюсках, иглокожих. Небольшое число видов астомат паразитирует и в позвоночных животных, а именно в земноводных. Однако астоматы распределены по разным группам хозяев совсем неравномерно. Существует одна группа «излюбленных» хозяев астомат, в которой сосредоточено свыше 75% их видов. Такой группой являются малощетинковые кольчатые черви (Oligochaeta), класс животных, к которому относится всем известный дождевой червь. Однако лишь часть видов малощетпнковых кольчецов живет в почве. Большая часть их — обитатели пресных вод, где они образуют заметную (а иногда и преобладающую) часть донного населения. Эти пресноводные малощетинковые кольчецы и являются «излюбленными» хозяевами астомат. Небольшое число видов олигохет встречается и в море, почти исключительно в прибрежной, литоральной (осушаемой во время отлива) зоне, но в морской фауне они не играют сколько-нибудь заметной роли.Органом, в котором локализуются в олигохетах инфузории астоматы, является кишечник. Поэтому многие черты строения астомат связаны с приспособлением к жизни в кишечнике. Через кишечник постоянно осуществляется движение пищи, иногда довольно активное. Паразиты кишечника, чтобы не быть вынесенными наружу (что равнозначно гибели), должны обладать приспособлениями, направленными на то, чтобы удержаться в кишечнике. Эти приспособления выражены не только у инфузорий, но и у всех других групп кишечных паразитов, к какой бы систематической группе они ни относились и в каких бы хозяевах ни паразитировали. Мы увидим это в дальнейшем, при знакомстве с разными группами паразитических червей.Тело всех паразитирующих в кишечнике олигохет астомат вытянуто в длину, что способствует их движению в просвете кишечника. Ресничный покров всегда хорошо развит, реснички покрывают все тело инфузорий и расположены тесными продольными рядами. В простейшем случае, например у представителей рода аноплофрий (Anoplophrуа, рис. 117, Б), никаких особых прикрепительных и опорных образований не развивается. Но у большинства видов других родов имеются разнообразные и подчас довольно сложные структуры двух категорий — опорные и прикрепительные, являющиеся формами приспособления к кишечному паразитизму.Опорные скелетные образования развиваются преимущественно на переднем конце тела, которому приходится испытывать механические воздействия и преодолевать препятствия, проталкиваясь в просвете кишечника между частицами пищи. У видов рода радиофрий (Radiophrya) на переднем конце с одной стороны тела (которую условно считают брюшной) располагаются очень прочные эластичные ребра (спикулы), лежащие в поверхностном слое эктоплазмы (рис. 117, Б, Г, Д). У видов рода менилелла (Mesnilella) тоже имеются опорные лучи (спикулы), которые на большей части своего протяжения лежат в более глубоких слоях цитоплазмы (в эндоплазме, рис. 117, А). Аналогично устроенные опорные образования развиты и у видов некоторых других родов астомат.У многих представителей этой интересной группы инфузорий имеются специальные структуры, позволяющие паразиту прочно прикрепляться к стенке кишечника. У известной уже нам радиофрии на самом переднем конце торчит наружу острый зубец, подвижно сочленяющийся с особыми опорными образованиями, имеющими форму наконечника стрелы. При помощи этого зубца радиофрия может прочно прикрепляться (зацепляться) к стенке кишечника.Другим типом прикрепительных образований астомат, но менее распространенным, являются присоски на переднем конце тела. Такого рода прикрепительный аппарат развит у видов рода гаптофрий (Haptophrya). Разные виды этого рода паразитируют в очень несхожих друг с другом хозяевах: одни — в ресничных червях (турбелляриях), другие — в земноводных.При изучении прикрепительных аппаратов паразитирующих в кишечнике различных хозяев инфузорий астомат обращает на себя внимание их большое сходство с аналогичными органами у других групп паразитов. Например, у ленточных червей основными формами органов прикрепления являются присоски и крючья. У инфузорий астомат — одноклеточных организмов — возникают структуры, весьма напоминающие органы прикрепления сложных многоклеточных животных (ленточные черви). Возникновение сходных структур, выполняющих аналогичную функцию у животных, относящихся к совершенно различным группам животного мира, связано с приспособлением к сходным условиям существования, в данном случае с приспособлением паразитов к жизни в кишечнике хозяев. Это явление носит название конвергенции.Бесполое размножение у некоторых инфузорий астомат протекает своеобразно. Вместо поперечного деления надвое, свойственного большинству инфузорий, у многих астомат происходит неравномерное деление (почкование). При этом почки, отделяющиеся на заднем конце, некоторое время остаются связанными с материнской особью (рис. 117, В). В результате получаются цепочки, состоящие из передней крупной и задних более мелких особей (почек). В дальнейшем почки постепенно отделяются от цепочки и переходят к самостоятельному существованию. Эта своеобразная форма размножения широко распространена, например, у известной уже нам радиофрий. Возникающие в результате почкования цепочки некоторых астомат напоминают по внешнему виду цепочки ленточных червей. Здесь мы вновь встречаемся с явлением конвергенции.Ядерный аппарат астомат имеет характерную для инфузорий структуру: макронуклеус, чаще всего лентовидной формы (рис. 117), и один микронуклеус. Сократительные вакуоли обычно хорошо развиты. У большинства видов имеется несколько (иногда свыше десятка) сократительных вакуолей, расположенных в один продольный ряд.Изучение распределения видов астомат по разным видам хозяев показывает, что большая часть астомат приурочена к строго определенным видам хозяев. Большинству астомат свойственна узкая специфичность: хозяином для них может служить лишь один вид животного.Несмотря на большое количество исследований, посвященных изучению инфузорий астомат, одна очень важная сторона их биологии остается совершенно неясной: каким образом происходит передача этих инфузорий от одной особи хозяина к другой? Никогда еще не удавалось наблюдать образование цист у этих инфузорий.Поэтому высказывают предположение, что заражение происходит активно — подвижными стадиями.Инфузории астоматы представляют собой яркий пример глубокого приспособления инфузорий к паразитическому образу жизни. Это приспособление отразилось на различных сторонах их организации (скелет, органоиды прикрепления) и жизненных отправлений. Они утеряли характерный для инфузорий способ питания путем заглатывания оформленных частиц пищи и перешли к осмотическому питанию.ИНФУЗОРИИ КИШЕЧНИКА МОРСКИХ ЕЖЕЙРазнообразный комплекс паразитических инфузорий обитает в кишечнике морских ежей. Морские ежи—богатый видами класс морских беспозвоночных животных, относящихся к типу иглокожих. Размеры морских ежей различны, но у большинства встречающихся в наших водах видов поперечник их почти шаровидного тела составляет 5—10 см.Очень многочисленны морские ежи в прибрежной зоне наших северных (Баренцево) и дальневосточных морей (Японского моря, тихоокеанского побережья Курильских островов). Большинство морских ежей питается растительной пищей, главным образом водорослями, которые они соскабливают с подводных предметов особыми острыми «зубами», окружающими ротовое отверстие. В кишечнике этих растительноядных ежей имеется богатая фауна инфузорий. Нередко они развиваются здесь в массовом количестве, и содержимое кишечника морского ежа под микроскопом почти так же «кишит» инфузориями, как содержимое рубца жвачных. Нужно сказать, что, кроме глубоких различий в условиях жизни инфузорий кишечника морского ежа и рубца жвачного, имеются и некоторые черты сходства. Они заключаются в том, что и там и здесь инфузории живут в среде, очень богатой растительными остатками. В настоящее время известно свыше 50 видов инфузорий, обитающих в кишечнике морских ежей, которые встречаются только в прибрежной зоне, где ежи питаются водорослями. На больших глубинах, где водоросли уже не растут, инфузорий в морских ежах нет.
В простейшем случае, например у представителей рода аноплофрий (Anoplophrуа, рис. 117, Б), никаких особых прикрепительных и опорных образований не развивается. Но у большинства видов других родов имеются разнообразные и подчас довольно сложные структуры двух категорий — опорные и прикрепительные, являющиеся формами приспособления к кишечному паразитизму.Опорные скелетные образования развиваются преимущественно на переднем конце тела, которому приходится испытывать механические воздействия и преодолевать препятствия, проталкиваясь в просвете кишечника между частицами пищи. У видов рода радиофрий (Radiophrya) на переднем конце с одной стороны тела (которую условно считают брюшной) располагаются очень прочные эластичные ребра (спикулы), лежащие в поверхностном слое эктоплазмы (рис. 117, Б, Г, Д). У видов рода менилелла (Mesnilella) тоже имеются опорные лучи (спикулы), которые на большей части своего протяжения лежат в более глубоких слоях цитоплазмы (в эндоплазме, рис. 117, А). Аналогично устроенные опорные образования развиты и у видов некоторых других родов астомат.У многих представителей этой интересной группы инфузорий имеются специальные структуры, позволяющие паразиту прочно прикрепляться к стенке кишечника. У известной уже нам радиофрии на самом переднем конце торчит наружу острый зубец, подвижно сочленяющийся с особыми опорными образованиями, имеющими форму наконечника стрелы. При помощи этого зубца радиофрия может прочно прикрепляться (зацепляться) к стенке кишечника.Другим типом прикрепительных образований астомат, но менее распространенным, являются присоски на переднем конце тела. Такого рода прикрепительный аппарат развит у видов рода гаптофрий (Haptophrya). Разные виды этого рода паразитируют в очень несхожих друг с другом хозяевах: одни — в ресничных червях (турбелляриях), другие — в земноводных.При изучении прикрепительных аппаратов паразитирующих в кишечнике различных хозяев инфузорий астомат обращает на себя внимание их большое сходство с аналогичными органами у других групп паразитов. Например, у ленточных червей основными формами органов прикрепления являются присоски и крючья. У инфузорий астомат — одноклеточных организмов — возникают структуры, весьма напоминающие органы прикрепления сложных многоклеточных животных (ленточные черви). Возникновение сходных структур, выполняющих аналогичную функцию у животных, относящихся к совершенно различным группам животного мира, связано с приспособлением к сходным условиям существования, в данном случае с приспособлением паразитов к жизни в кишечнике хозяев. Это явление носит название конвергенции.Бесполое размножение у некоторых инфузорий астомат протекает своеобразно. Вместо поперечного деления надвое, свойственного большинству инфузорий, у многих астомат происходит неравномерное деление (почкование). При этом почки, отделяющиеся на заднем конце, некоторое время остаются связанными с материнской особью (рис. 117, В). В результате получаются цепочки, состоящие из передней крупной и задних более мелких особей (почек). В дальнейшем почки постепенно отделяются от цепочки и переходят к самостоятельному существованию. Эта своеобразная форма размножения широко распространена, например, у известной уже нам радиофрий. Возникающие в результате почкования цепочки некоторых астомат напоминают по внешнему виду цепочки ленточных червей. Здесь мы вновь встречаемся с явлением конвергенции.Ядерный аппарат астомат имеет характерную для инфузорий структуру: макронуклеус, чаще всего лентовидной формы (рис. 117), и один микронуклеус. Сократительные вакуоли обычно хорошо развиты. У большинства видов имеется несколько (иногда свыше десятка) сократительных вакуолей, расположенных в один продольный ряд.Изучение распределения видов астомат по разным видам хозяев показывает, что большая часть астомат приурочена к строго определенным видам хозяев. Большинству астомат свойственна узкая специфичность: хозяином для них может служить лишь один вид животного.Несмотря на большое количество исследований, посвященных изучению инфузорий астомат, одна очень важная сторона их биологии остается совершенно неясной: каким образом происходит передача этих инфузорий от одной особи хозяина к другой? Никогда еще не удавалось наблюдать образование цист у этих инфузорий.Поэтому высказывают предположение, что заражение происходит активно — подвижными стадиями.Инфузории астоматы представляют собой яркий пример глубокого приспособления инфузорий к паразитическому образу жизни. Это приспособление отразилось на различных сторонах их организации (скелет, органоиды прикрепления) и жизненных отправлений. Они утеряли характерный для инфузорий способ питания путем заглатывания оформленных частиц пищи и перешли к осмотическому питанию.ИНФУЗОРИИ КИШЕЧНИКА МОРСКИХ ЕЖЕЙРазнообразный комплекс паразитических инфузорий обитает в кишечнике морских ежей. Морские ежи—богатый видами класс морских беспозвоночных животных, относящихся к типу иглокожих. Размеры морских ежей различны, но у большинства встречающихся в наших водах видов поперечник их почти шаровидного тела составляет 5—10 см.Очень многочисленны морские ежи в прибрежной зоне наших северных (Баренцево) и дальневосточных морей (Японского моря, тихоокеанского побережья Курильских островов). Большинство морских ежей питается растительной пищей, главным образом водорослями, которые они соскабливают с подводных предметов особыми острыми «зубами», окружающими ротовое отверстие. В кишечнике этих растительноядных ежей имеется богатая фауна инфузорий. Нередко они развиваются здесь в массовом количестве, и содержимое кишечника морского ежа под микроскопом почти так же «кишит» инфузориями, как содержимое рубца жвачных. Нужно сказать, что, кроме глубоких различий в условиях жизни инфузорий кишечника морского ежа и рубца жвачного, имеются и некоторые черты сходства. Они заключаются в том, что и там и здесь инфузории живут в среде, очень богатой растительными остатками. В настоящее время известно свыше 50 видов инфузорий, обитающих в кишечнике морских ежей, которые встречаются только в прибрежной зоне, где ежи питаются водорослями. На больших глубинах, где водоросли уже не растут, инфузорий в морских ежах нет. Что же представляет собой фауна паразитических инфузорий кишечника морских ежей в отношении своего систематического состава (принадлежности их к определенным систематическим группам)? Оказывается, что в кишечнике морских ежей встречаются представители разных отрядов и семейств инфузорий. На рисунке 118 изображены некоторые характерные представители этой фауны из кишечника морских ежей, обитающих в прибрежных водах Японского моря в окрестностях Владивостока. Здесь имеются различные равноресничные инфузории (Holotricha), брюхоресничные (Hypotrycha), малоресничные (Oligotricha). Интересно отметить, что некоторые из видов инфузорий из кишечника морских ежей относятся к родам, имеющим свободноживущие виды. У них нет тех глубоких приспособлений, которые можпо видеть у энтодиниоморф (Entodiniomorpha) к жизни в кишечнике копытных или у астомат (Astomata) к жизни в кишечнике малощетинковых кольчецов. Лишь у некоторых видов, например у энторипидиум (Entorhipidium), развиваются скелетные лучи (подобно тому как это мы видели у радиофрий, рис. 117). Другие же виды мало чем отличаются от свободноживущих инфузорий.По образу жизни и характеру питания большинство инфузорий кишечника морских ежей растительноядны. Они питаются водорослями, которые в большом количестве наполняют кишечник хозяина. Некоторые виды довольно «привередливы» в выборе пищи. Например, стробилидий (Strobilidium, рис. 118, А) питается почти исключительно крупными диатомовыми водорослями. Имеются здесь и хищники, поедающие представителей других, более мелких видов инфузорий.У инфузорий из кишечника морских ежей, в отличие от астомат, не наблюдается какой-либо строгой приуроченности к определенным видам хозяев. Они живут в самых разных видах морских ежей, питающихся водорослями.Как уже говорилось, простейшие из кишечника морских ежей хотя и ведут эндо-паразитический образ жизни, но почти не отличаются от свободноживущих родичей. Вероятно, эти инфузории относительно недавно перешли к паразитизму. Этот переход осуществлялся независимо для разных видов. В новых условиях паразитической жизни они нашли среду, не очень отличающуюся от морской воды. В кишечнике морских ежей всегда много заглоченной вместе с пищей воды и много водорослей на разных стадиях переваривания. Эти условия оказались весьма благоприятными для жизни инфузорий, которые и размножаются здесь в больших количествах.Пути заражения морских ежей инфузориями не изучены. Однако здесь с большой долей вероятности можно допустить, что оно происходит активными свободноплавающими формами. Дело в том, что инфузории из кишечника морских ежей могут длительное время (многие часы) жить в морской воде. Однако они уже настолько приспособились к жизни в кишечнике ежей, что вне тела их, в морской воде, рано или поздно погибают.Заканчивая знакомство с инфузориями, следует еще раз подчеркнуть, что они представляют собой богатую видами, обширную и процветающую группу (класс) животного мира. Оставаясь на уровне клеточной организации, инфузории достигли по сравнению с другими классами простейших наибольшей сложности строения и функций.Особенно существенную роль в этом прогрессивном развитии (эволюции) сыграло, вероятно, преобразование ядерного аппарата и возникновение ядерного дуализма (качественной неравноценности ядер). Богатство макронуклеуса нуклеиновыми веществами связано с активными процессами обмена веществ, с энергично протекающими процессами синтеза белков цитоплазмы и ядер.Организация инфузорий оказалась очень пластичной, что позволило им приспособиться к различным средам обитания в морской и пресной воде и дать разнообразные паразитические формы.ЗаключениеМы подошли к концу нашего обзора строения и образа жизни обширного типа животного мира — простейших. Характерной особенностью их, как уже неоднократно подчеркивалось выше, является одноклеточность. В отношении своего строения простейшие — клетки. Однако они несравнимы с клетками, входящими в состав тела многоклеточных организмов, потому что сами являются организмами. Таким образом, простейшие — это организмы на клеточном уровне организации. Некоторые высокоорганизованные простейшие, обладающие многими ядрами, уже как бы выходят за морфологические пределы строения клетки, что дает основание некоторым ученым называть таких простейших «надклеточными». Это мало меняет существо дела, ибо типичной для Protozoa является все же одноклеточная организация.В пределах одноклеточности простейшие прошли длинный путь эволюционного развития и дали огромное разнообразие форм, приспособленных к самым различным условиям жизни. В основе родословного ствола простейших стоят два класса: саркодовые и жгутиконосцы. До сих пор в науке дебатируется вопрос, какой из этих классов более примитивный. С одной стороны, низшие представители саркодовых (амебы) обладают самым примитивным строением. Но жгутиконосцы обнаруживают наибольшую пластичность типа обмена веществ и стоят как бы на границе между животным и растительным миром. В жизненном цикле некоторых саркодовых (например, фораминифер) имеются жгутиковые стадии (гаметы), что указывает на их связь со жгутиконосцами. Очевидно, что ни современные саркодовые, ни современные жгутиконосцы не могут быть исходной группой эволюции животного мира, ибо они сами прошли большой путь исторического развития и выработали многочисленные приспособления к современным условиям жизни на Земле. Вероятно, оба этих класса современных простейших следует рассматривать как два ствола в эволюции, берущих свое начало от древних, не сохранившихся до наших дней форм, которые жили на заре развития жизни на нашей планете.В дальнейшей эволюции простейших произошли изменения различного характера. Некоторые из них привели к общему повышению уровня организации, повышению активности, интенсивности жизненных процессов. К числу таких филогенетических (эволюционных) преобразований следует, например, отнести развитие органоидов движения и захвата пищи, которое достигло высокого совершенства в классе инфузорий. Бесспорно, что реснички представляют собой органоиды, соответствующие (гомологичные) жгутикам. В то время как у жгутиконосцев, за немногими исключениями, число жгутиков невелико, у инфузорий число ресничек достигло многих тысяч. Развитие ресничного аппарата резко повысило активность простейших, сделало более многообразными и сложными формы их взаимоотношений со средой, формы реакций на внешние раздражения. Наличие дифференцированного ресничного аппарата, бесспорно, явилось одной из основных причин прогрессивной эволюции в классе инфузорий, где возникло большое многообразие форм, приспособленных к разным средам обитания.Развитие ресничного аппарата инфузорий — пример такого рода эволюционных изменений, которые были названы акад. Северцовым ароморфозами. Ароморфозы характеризуются общим повышением организации, развитием приспособлений широкого значения. Под повышением организации понимают изменения, которые вызывают усиление жизнедеятельности организма; они связаны с функциональной дифференцировкой его частей, ведут к более многообразным формам связи организма с окружающей средой. Развитие ресничного аппарата инфузорий как раз и относится к такого рода преобразованиям строения в процессе эволюции. Это типичный ароморфоз.У простейших, как это было подчеркнуто В. А. Догелем, изменения типа ароморфозов обычно бывают связаны с увеличением числа органоидов. Происходит полимеризация органоидов. Развитие ресничного аппарата у инфузорий— типичный пример такого рода изменений. Вторым примером ароморфоза в эволюции инфузорий может служить их ядерный аппарат. Мы рассмотрели выше особенности строения ядра инфузорий. Ядерный дуализм инфузорий (наличие микронуклеуса и макронуклеуса) сопровождался увеличением в составе макронуклеуса числа хромосом (явление полиплоидии). Поскольку хромосомы связаны с основными синтетическими процессами в клетке, в первую очередь с синтезом белков, этот процесс привел к общему повышению интенсивности основных жизненных функций. И здесь имела место полимеризация, затронувшая хромосомные комплексы ядра.Инфузории — одна из наиболее многочисленных и прогрессивных групп простейших, происходит от жгутиконосцев. Об этом говорит полное морфологическое сходство их органоидов движения. Этот этап эволюции был связан с двумя большими ароморфозами: один из них затрагивал органоиды движения, второй — ядерный аппарат. Оба эти типа изменений связаны между собой, так как оба ведут к повышению жизнедеятельности и усложнению форм взаимосвязей с внешней средой.Наряду с ароморфозами существует и другой тип эволюционных изменений, выражающихся в выработке приспособлений (адаптации) к определенным, резко очерченным условиям существования. Этот тип эволюционных изменений Северцовым был назван идиоадаптациями. В эволюции простейших этот тип изменений играл очень большую роль. Выше, при рассмотрении разных классов простейших, приведепы многочисленные примеры идиоадаптивных изменений. Приспособления к планктонному образу жизни у разных групп простейших, приспособления к жизни в песке у инфузорий, образование защитных оболочек ооцист у кокцидий и многое другое — все это идиоадаптации, игравшие большую роль в возникновении и развитии отдельных групп, но не связанные с общими прогрессивными изменениями организации.Приспособления к различным конкретным средам обитания у простейших носят весьма разнообразный характер. Они обеспечили широкое распространение этого типа в самых разных средах обитания, что подробно рассматривалось выше, при описании отдельных классов.Следует еще остановиться на вопросе о паразитизме у простейших. В типе простейших переход от свободного образа жизни к паразитизму распространен очень широко. Паразитические формы мы встречаем среди всех классов Protozoa. Кроме того, два класса — споровики и книдоспоридии — целиком состоят из паразитов. Один из них (споровики), как вы видели, взял свое начало от жгутиконосцев, второй (книдоспоридий) —от саркодовых. По ходу эволюции паразитических групп простейших у них выработались весьма сложные жизненные циклы, обеспечивающие как размножение паразитов, так и их широкое распространение в природе. Многие паразитические простейшие имеют большое практическое значение для человека как возбудители тяжелых заболеваний самого человека, домашних и промысловых млекопитающих, птиц, рыб, а также полезных насекомых. Поэтому борьба с протозойными паразитарными заболеваниями, профилактика и лечение их играют большую роль в медицинской и ветеринарной практике.
Что же представляет собой фауна паразитических инфузорий кишечника морских ежей в отношении своего систематического состава (принадлежности их к определенным систематическим группам)? Оказывается, что в кишечнике морских ежей встречаются представители разных отрядов и семейств инфузорий. На рисунке 118 изображены некоторые характерные представители этой фауны из кишечника морских ежей, обитающих в прибрежных водах Японского моря в окрестностях Владивостока. Здесь имеются различные равноресничные инфузории (Holotricha), брюхоресничные (Hypotrycha), малоресничные (Oligotricha). Интересно отметить, что некоторые из видов инфузорий из кишечника морских ежей относятся к родам, имеющим свободноживущие виды. У них нет тех глубоких приспособлений, которые можпо видеть у энтодиниоморф (Entodiniomorpha) к жизни в кишечнике копытных или у астомат (Astomata) к жизни в кишечнике малощетинковых кольчецов. Лишь у некоторых видов, например у энторипидиум (Entorhipidium), развиваются скелетные лучи (подобно тому как это мы видели у радиофрий, рис. 117). Другие же виды мало чем отличаются от свободноживущих инфузорий.По образу жизни и характеру питания большинство инфузорий кишечника морских ежей растительноядны. Они питаются водорослями, которые в большом количестве наполняют кишечник хозяина. Некоторые виды довольно «привередливы» в выборе пищи. Например, стробилидий (Strobilidium, рис. 118, А) питается почти исключительно крупными диатомовыми водорослями. Имеются здесь и хищники, поедающие представителей других, более мелких видов инфузорий.У инфузорий из кишечника морских ежей, в отличие от астомат, не наблюдается какой-либо строгой приуроченности к определенным видам хозяев. Они живут в самых разных видах морских ежей, питающихся водорослями.Как уже говорилось, простейшие из кишечника морских ежей хотя и ведут эндо-паразитический образ жизни, но почти не отличаются от свободноживущих родичей. Вероятно, эти инфузории относительно недавно перешли к паразитизму. Этот переход осуществлялся независимо для разных видов. В новых условиях паразитической жизни они нашли среду, не очень отличающуюся от морской воды. В кишечнике морских ежей всегда много заглоченной вместе с пищей воды и много водорослей на разных стадиях переваривания. Эти условия оказались весьма благоприятными для жизни инфузорий, которые и размножаются здесь в больших количествах.Пути заражения морских ежей инфузориями не изучены. Однако здесь с большой долей вероятности можно допустить, что оно происходит активными свободноплавающими формами. Дело в том, что инфузории из кишечника морских ежей могут длительное время (многие часы) жить в морской воде. Однако они уже настолько приспособились к жизни в кишечнике ежей, что вне тела их, в морской воде, рано или поздно погибают.Заканчивая знакомство с инфузориями, следует еще раз подчеркнуть, что они представляют собой богатую видами, обширную и процветающую группу (класс) животного мира. Оставаясь на уровне клеточной организации, инфузории достигли по сравнению с другими классами простейших наибольшей сложности строения и функций.Особенно существенную роль в этом прогрессивном развитии (эволюции) сыграло, вероятно, преобразование ядерного аппарата и возникновение ядерного дуализма (качественной неравноценности ядер). Богатство макронуклеуса нуклеиновыми веществами связано с активными процессами обмена веществ, с энергично протекающими процессами синтеза белков цитоплазмы и ядер.Организация инфузорий оказалась очень пластичной, что позволило им приспособиться к различным средам обитания в морской и пресной воде и дать разнообразные паразитические формы.ЗаключениеМы подошли к концу нашего обзора строения и образа жизни обширного типа животного мира — простейших. Характерной особенностью их, как уже неоднократно подчеркивалось выше, является одноклеточность. В отношении своего строения простейшие — клетки. Однако они несравнимы с клетками, входящими в состав тела многоклеточных организмов, потому что сами являются организмами. Таким образом, простейшие — это организмы на клеточном уровне организации. Некоторые высокоорганизованные простейшие, обладающие многими ядрами, уже как бы выходят за морфологические пределы строения клетки, что дает основание некоторым ученым называть таких простейших «надклеточными». Это мало меняет существо дела, ибо типичной для Protozoa является все же одноклеточная организация.В пределах одноклеточности простейшие прошли длинный путь эволюционного развития и дали огромное разнообразие форм, приспособленных к самым различным условиям жизни. В основе родословного ствола простейших стоят два класса: саркодовые и жгутиконосцы. До сих пор в науке дебатируется вопрос, какой из этих классов более примитивный. С одной стороны, низшие представители саркодовых (амебы) обладают самым примитивным строением. Но жгутиконосцы обнаруживают наибольшую пластичность типа обмена веществ и стоят как бы на границе между животным и растительным миром. В жизненном цикле некоторых саркодовых (например, фораминифер) имеются жгутиковые стадии (гаметы), что указывает на их связь со жгутиконосцами. Очевидно, что ни современные саркодовые, ни современные жгутиконосцы не могут быть исходной группой эволюции животного мира, ибо они сами прошли большой путь исторического развития и выработали многочисленные приспособления к современным условиям жизни на Земле. Вероятно, оба этих класса современных простейших следует рассматривать как два ствола в эволюции, берущих свое начало от древних, не сохранившихся до наших дней форм, которые жили на заре развития жизни на нашей планете.В дальнейшей эволюции простейших произошли изменения различного характера. Некоторые из них привели к общему повышению уровня организации, повышению активности, интенсивности жизненных процессов. К числу таких филогенетических (эволюционных) преобразований следует, например, отнести развитие органоидов движения и захвата пищи, которое достигло высокого совершенства в классе инфузорий. Бесспорно, что реснички представляют собой органоиды, соответствующие (гомологичные) жгутикам. В то время как у жгутиконосцев, за немногими исключениями, число жгутиков невелико, у инфузорий число ресничек достигло многих тысяч. Развитие ресничного аппарата резко повысило активность простейших, сделало более многообразными и сложными формы их взаимоотношений со средой, формы реакций на внешние раздражения. Наличие дифференцированного ресничного аппарата, бесспорно, явилось одной из основных причин прогрессивной эволюции в классе инфузорий, где возникло большое многообразие форм, приспособленных к разным средам обитания.Развитие ресничного аппарата инфузорий — пример такого рода эволюционных изменений, которые были названы акад. Северцовым ароморфозами. Ароморфозы характеризуются общим повышением организации, развитием приспособлений широкого значения. Под повышением организации понимают изменения, которые вызывают усиление жизнедеятельности организма; они связаны с функциональной дифференцировкой его частей, ведут к более многообразным формам связи организма с окружающей средой. Развитие ресничного аппарата инфузорий как раз и относится к такого рода преобразованиям строения в процессе эволюции. Это типичный ароморфоз.У простейших, как это было подчеркнуто В. А. Догелем, изменения типа ароморфозов обычно бывают связаны с увеличением числа органоидов. Происходит полимеризация органоидов. Развитие ресничного аппарата у инфузорий— типичный пример такого рода изменений. Вторым примером ароморфоза в эволюции инфузорий может служить их ядерный аппарат. Мы рассмотрели выше особенности строения ядра инфузорий. Ядерный дуализм инфузорий (наличие микронуклеуса и макронуклеуса) сопровождался увеличением в составе макронуклеуса числа хромосом (явление полиплоидии). Поскольку хромосомы связаны с основными синтетическими процессами в клетке, в первую очередь с синтезом белков, этот процесс привел к общему повышению интенсивности основных жизненных функций. И здесь имела место полимеризация, затронувшая хромосомные комплексы ядра.Инфузории — одна из наиболее многочисленных и прогрессивных групп простейших, происходит от жгутиконосцев. Об этом говорит полное морфологическое сходство их органоидов движения. Этот этап эволюции был связан с двумя большими ароморфозами: один из них затрагивал органоиды движения, второй — ядерный аппарат. Оба эти типа изменений связаны между собой, так как оба ведут к повышению жизнедеятельности и усложнению форм взаимосвязей с внешней средой.Наряду с ароморфозами существует и другой тип эволюционных изменений, выражающихся в выработке приспособлений (адаптации) к определенным, резко очерченным условиям существования. Этот тип эволюционных изменений Северцовым был назван идиоадаптациями. В эволюции простейших этот тип изменений играл очень большую роль. Выше, при рассмотрении разных классов простейших, приведепы многочисленные примеры идиоадаптивных изменений. Приспособления к планктонному образу жизни у разных групп простейших, приспособления к жизни в песке у инфузорий, образование защитных оболочек ооцист у кокцидий и многое другое — все это идиоадаптации, игравшие большую роль в возникновении и развитии отдельных групп, но не связанные с общими прогрессивными изменениями организации.Приспособления к различным конкретным средам обитания у простейших носят весьма разнообразный характер. Они обеспечили широкое распространение этого типа в самых разных средах обитания, что подробно рассматривалось выше, при описании отдельных классов.Следует еще остановиться на вопросе о паразитизме у простейших. В типе простейших переход от свободного образа жизни к паразитизму распространен очень широко. Паразитические формы мы встречаем среди всех классов Protozoa. Кроме того, два класса — споровики и книдоспоридии — целиком состоят из паразитов. Один из них (споровики), как вы видели, взял свое начало от жгутиконосцев, второй (книдоспоридий) —от саркодовых. По ходу эволюции паразитических групп простейших у них выработались весьма сложные жизненные циклы, обеспечивающие как размножение паразитов, так и их широкое распространение в природе. Многие паразитические простейшие имеют большое практическое значение для человека как возбудители тяжелых заболеваний самого человека, домашних и промысловых млекопитающих, птиц, рыб, а также полезных насекомых. Поэтому борьба с протозойными паразитарными заболеваниями, профилактика и лечение их играют большую роль в медицинской и ветеринарной практике.
Жизнь животных: в 6-ти томах. — М.: Просвещение. Под редакцией профессоров Н.А.Гладкова, А.В.Михеева. 1970.
.