- Центральная нервная система
-
основная часть нервной системы животных и человека, состоящая из скопления нервных клеток (нейронов) и их отростков; представлена у беспозвоночных системой тесно связанных между собой нервных узлов (ганглиев), у позвоночных животных и человека спинным и головным мозгом. Главная и специфическая функция Ц. н. с. — осуществление простых и сложных высокодифференцированных отражательных реакций, получивших название Рефлексов. У высших животных и человека низшие и средние отделы Ц. н. с. — Спинной мозг, Продолговатый мозг, Средний мозг, Промежуточный мозг и Мозжечок — регулируют деятельность отдельных органов и систем высокоразвитого организма, осуществляют связь и взаимодействие между ними, обеспечивают единство организма и целостность его деятельности. Высший отдел Ц. н. с. — Кора больших полушарий головного мозга и ближайшие подкорковые образования — в основном регулирует связь и взаимоотношения организма как единого целого с окружающей средой.Основные черты строения и функции. Ц. н. с. связана со всеми органами и тканями через периферическую нервную систему, которая у позвоночных включает Черепномозговые нервы, отходящие от головного мозга, и Спинномозговые нервы — от спинного мозга, межпозвонковые нервные узлы, а также периферический отдел вегетативной нервной системы — нервные узлы, с подходящими к ним (преганглионарными) и отходящими от них (постганглионарными) нервными волокнами. Чувствительные, или афферентные, нервные приводящие волокна несут возбуждение в Ц. н. с. от периферических рецепторов; по отводящим эфферентным (двигательным и вегетативным) нервным волокнам возбуждение из Ц. н. с. направляется к клеткам исполнительных рабочих аппаратов (мышцы, железы, сосуды и т.д.). Во всех отделах Ц. н. с. имеются афферентные нейроны, воспринимающие приходящие с периферии раздражения, и эфферентные нейроны, посылающие нервные импульсы на периферию к различным исполнительным эффекторным органам. Афферентные и эфферентные клетки своими отростками могут контактировать между собой и составлять двухнейронную рефлекторную дугу (См. Рефлекторная дуга), осуществляющую элементарные рефлексы (например, сухожильные рефлексы спинного мозга). Но, как правило, в рефлекторной дуге между афферентными и эфферентными нейронами расположены вставочные нервные клетки, или интернейроны (рис. 1). Связь между различными отделами Ц. н. с. осуществляется также с помощью множества отростков афферентных, эфферентных и вставочных нейронов этих отделов, образующих внутрицентральные короткие и длинные проводящие пути. В состав Ц. н. с. входят также клетки нейроглии (См. Нейроглия), которые выполняют в ней опорную функцию, а также участвуют в метаболизме нервных клеток.Рефлекторный принцип работы Ц. н. с., как основной принцип её деятельности, до 19 в. был экспериментально установлен и изучен только в отношении деятельности отделов, расположенных ниже уровня больших полушарий головного мозга. Были выявлены общие закономерности и приспособительный характер рефлекторной деятельности Ц. н. с., а также специфические, частные особенности функций её различных отделов. В 19 в. И. М. Сеченовым и некоторыми др. прогрессивными учёными была высказана смелая идея о рефлекторной природе деятельности также и высших отделов Ц. н. с., в том числе и психической деятельности. Эта передовая, материалистическая идея послужила основой для классических экспериментальных и теоретических исследований И. П. Павлова, приведших к созданию им учения о высшей нервной деятельности (См. Высшая нервная деятельность). Павлов установил, что рефлексы, осуществляемые у высокоразвитых организмов корой больших полушарий головного мозга, в отличие от рефлексов всех нижележащих отделов Ц. н. с., не врождённые, а формируются в течение индивидуальной жизни организма в процессе его взаимодействия с окружающей средой и обеспечивают его наиболее совершенное приспособление к условиям существования. Этот новый класс рефлексов, в отличие от врождённых, или безусловных рефлексов (См. Безусловные рефлексы), Павлов назвал условными рефлексами (См. Условные рефлексы).В противоположность первоначальным, примитивным представлениям о рефлексе как о трафаретной, машинообразной, к тому же пассивной форме ответной деятельности Ц. н. с., классики нейрофизиологии Э. Пфлюгер, Сеченов, Павлов, Ч. Шеррингтон и др. установили, что и безусловные и особенно условные рефлексы характеризуются значительной динамичностью и вариабельностью: рефлекторные реакции на одни и те же раздражители зависят от условий внешней и внутренней среды организма и от функционального состояния самой Ц. н. с. Основные закономерности деятельности Ц. н. с. связаны в первую очередь с особенностями рефлекторной дуги — структурной основы каждого рефлекторного акта. Рефлекторная дуга проводит возбуждение только в одном направлении — от рецепторного конца к исполнительному органу. Это обусловлено присущей всем нервным клеткам структурной и функциональной поляризацией: на концевых разветвлениях аксонов каждого нейрона существуют микроструктурные образования, т. н. Синапсы, посредством которых он контактирует с телами или Дендритами др. нейронов и односторонне передаёт им свою активность (Белла - Мажанди закон). Многообразные наружные и внутренние рецепторы организма, специализированные в процессе эволюции к тонкому и совершенному восприятию отдельных, качественно специфических видов энергии — световой, звуковой, тепловой, механической и химической, трансформируют их в процесс нервного возбуждения (См. Возбуждение), которое в виде ритмических импульсов передаётся последовательно от одних звеньев рефлекторной дуги к другим. Возбуждение на своём многоэтапном пути к конечному звену претерпевает значительные изменения в ритме, интенсивности, скорости и характере. В исполнительных органах рефлекторное возбуждение может порождать многообразные эффекты, обусловленные специфическими особенностями структуры и функций самих исполнительных органов (мышц, желёз, сосудов и т.д.).В отличие от простого нервного проводника, функциональные особенности центральных отделов рефлекторной дуги характеризуются сравнительно медленным возникновением и протеканием в них процесса возбуждения и фазовых колебаний уровня возбудимости, порождаемых волнами возбуждения. Эта своеобразная функциональная «инертность» — относительно длительное сохранение состояния повышенной возбудимости — обусловливает, в частности, явление суммации (когда слабые неэффективные стимуляции при повторении становятся эффективными), а также близкое ему явление т. н. облегчения. Центральные отделы рефлекторной дуги способны изменять, трансформировать ритм раздражений. Они отвечают на раздражение рецепторов, как правило, своим собственным, присущим им ритмом волн возбуждения: то более высоким, чем ритм раздражения, то более низким; при этом между силой раздражения и интенсивностью рефлекторной реакции имеется только относительное соответствие, наблюдаемое обычно при действии раздражителей умеренной силы. Если действуют сильные и продолжительные раздражения, нервные центры приходят в состояние торможения (См. Торможение), в этом случае слабое раздражение начинает вызывать больший рефлекторный эффект, чем сильное (т. н. парадоксальные отношения). Для центральных отделов рефлекторной дуги характерны также относительно высокая утомляемость и отчётливо выраженная чувствительность к биохимическим изменениям внутренней среды организма, особенно к недостатку кислорода в крови и к некоторым специфическим ядам. Все эти особенности в основном обусловлены свойствами синапсов, дендритов и тел самих нейронов.Для простоты рефлекторная дуга обычно изображается в виде цепи из ряда одиночных клеток разного рода: рецепторной клетки, афферентной, вставочной и эфферентной нервных клеток и исполнительной клетки. В действительности рефлекторная дуга объединяет множество таких цепей, специфические звенья которых представляют собой не одиночную клетку того или иного рода, а ансамбль взаимосвязанных однородных клеток. При этом совокупность рецепторных клеток формирует рецептивное поле рефлекса (Рефлексогенные зоны), объединение исполнительных клеток создаёт его рабочий орган, а совокупность расположенных в Ц. н. с. нейронов образует соответствующий Нервный центр. Павлов определил нервный центр как совокупность нервных элементов, расположенных в различных отделах Ц. н. с., тесно связанных между собой, составляющих единую систему и осуществляющих регуляцию той или иной функции организма. Развивая и конкретизируя эту концепцию, Э. А. Асратян предложил рассматривать центральную часть дуги безусловного рефлекса как многоэтажную структуру, состоящую из ряда ветвей, каждая из которых проходит по одному из отделов Ц. н. с., участвующих в данном рефлексе, и характеризуется специфичностью (рис. 2). Эти ветви не равноценны по своему значению в выполнении рефлексов: для одних рефлексов главными являются ветви одних уровней, для других — ветви других уровней. Например, в дуге сердечно-сосудистых и дыхательных рефлексов главная ветвь проходит через продолговатый мозг, а в дуге пищевых, защитных и половых рефлексов — через промежуточный мозг. Ветвь подобной многоэтажной дуги, проходящая через кору большого мозга, соответствует тому, что Павлов назвал корковым представительством безусловного рефлекса, и служит основанием для образования соответствующих условных рефлексов.Координация рефлекторных реакций. Ц. н. с. осуществляет множество разнообразных рефлексов, отличающихся друг от друга рецепторными, центральными и эффекторными органами, родом, характером, знаком, происхождением, степенью сложности и т.п. Тем не менее они возникают и протекают не разрозненно и хаотично, а координированно — в определённом составе, порядке и последовательности в соответствии с текущими потребностями организма, т. е. как объединённая целостная приспособительная деятельность. Координационная деятельность Ц. н. с. обусловлена её структурными и функциональными особенностями. Степень генетического родства между разными структурными элементами Ц. н. с. и характер связи между ними — важные предпосылки их взаимодействия и координации рефлексов. Особенно существенны в этом отношении структурные особенности, называемые расхождением и схождением путей. При расхождении, или дивергенции, путей каждый афферентный нейрон посредством множества концевых разветвлений аксонов контактирует с большим числом эфферентных нейронов непосредственно или через промежуточный нейрон (рис. 3, а). Это создаёт предпосылки для активирования одним афферентным нейроном множества близких к нему и отдалённых эфферентных нейронов и связанных с ними рефлексов в определённой последовательности. Например, у кошки, лишённой больших полушарий головного мозга, при слабой стимуляции подошвы задней лапы сначала появляется рефлекс сгибания голеностопного сустава; при постепенном усилении стимулирующего тока один за другим к нему присоединяются рефлексы сгибания коленного и тазобедренного суставов той же конечности, разгибательный рефлекс противоположной задней лапы, разгибательный рефлекс одноимённой передней конечности и, наконец, рефлекс сгибания противоположной передней конечности. Это происходит в результате постепенного распространения (иррадиации) возбуждения, возникшего в афферентном звене изначального рефлекса, на связанные с ним структуры близких и отдалённых рефлексов спинного мозга посредством коротких и длинных внутриспинальных проводящих путей. При схождении, или конвергенции, путей нескольких афферентных нейронов с помощью концевых разветвлений аксонов контактируют с одним эфферентным нейроном непосредственно или через промежуточные нейроны (рис. 3, б). В последнем случае ветви ряда афферентных нейронов сходятся к одному интернейрону, который служит своеобразным «общим путём», а ветви нескольких таких интернейронов, в свою очередь, сходятся к одному эфферентному нейрону или их ансамблю как к «общему конечному пути», который «заблокирован» для др. потоков возбуждения. Конвергенция путей в Ц. н. с. создаёт структурную предпосылку для использования одного эфферентного нейрона многими одно- и разнородными афферентными нейронами, число которых превышает число эфферентных в несколько раз. Например, если раздельные раздражения слабым током двух разных участков рецептивного поля чесательного рефлекса не вызывают надлежащих рефлексов, то одновременное раздражение обоих участков током той же силы приводит к появлению чёткого рефлекса в результате суммации подпороговых возбуждений. Взаимное усиление рефлексов происходит также при одновременном раздражении рецептивных полей двух разных рефлексов, имеющих общий конечный путь и вызывающих один и тот же эффект. Явления дивергенции и конвергенции первоначально были обнаружены и изучены в деятельности спинного мозга, но в последующем было установлено, что они присущи и др. отделам Ц. н. с., особенно коре больших полушарий головного мозга. В высших отделах Ц. н. с. резко преобладают афферентные нервные элементы над эфферентными и поэтому весьма ярко выражен принцип схождения множества путей к сенсомоторной области как к основному кортикальному «общему пути». Это служит структурной предпосылкой для участия афферентов в осуществлении разного рода двигательных безусловных рефлексов и для выработки на их основе многообразных условных рефлексов. Новейшие электрофизиологические методики позволили многим современным нейрофизиологам (итальянский физиолог Дж. Моруцци, П. Г. Костюк и др.) выявить и исследовать на уровне отдельных нейронов микроструктурные и микрофункциональные основы и механизмы дивергенции и конвергенции, особенно в нейронах спинного мозга, ретикулярной формации, зрительных бугров, коры головного мозга и др. отделов Ц. н. с. Установлено, что на теле и дендритах каждого нейрона ретикулярной формации и коры головного мозга существуют синаптические контакты множества др. нейронов, активируемых раздражителями разных модальностей и оказывающих как возбуждающее, так и тормозящее влияние на «общий путь».В координации рефлексов и в деятельности Ц. н. с. в целом важное значение имеют такие факторы, как исходное функциональное состояние вовлечённых в данный процесс нервных структур, сила, биологическое значение и продолжительность действия внешнего стимулирующего агента, влияние внутренних нервных и гуморальных факторов и т.п. Однако главная функциональная основа координации рефлекторной деятельности заключается в способности Ц. н. с. процессом возбуждения активировать одни синаптические контакты и пути и (что особенно важно) одновременно блокировать процессом торможения др. синаптические контакты и пути в самых многообразных комбинациях и пространственно-временных соотношениях, соответственно текущим потребностям организма и наличным обстоятельствам во внешней среде. Важнейшая роль торможения в координационной деятельности Ц. н. с. наглядно проявляется в антагонистических взаимодействиях рефлексов, особенно в случаях, когда одновременно стимулируются разные рецепторы или рецептивные поля, имеющие общий исполнительный орган, но вызывающие разнородные рефлексы. Такого типа взаимотормозящие отношения существуют между локомоторными, защитными и позными рефлексами, в которых участвуют одни и те же разгибательные мотонейроны с иннервируемыми ими мышцами конечностей. Каждый из этих рефлексов может сепаратно возникать и беспрепятственно протекать, если стимулируется только его рецептивное поле. Но когда во время одного из рефлексов стимулируется также рецептивное поле другого, а тем более обоих рефлексов, то возникает конфликтная ситуация: происходит борьба за овладение общим для них «конечным путём». Преобладает тот рефлекс, рецептивное поле которого в данный момент стимулируется сильнее и который в данных условиях оказывается нужнее организму. Остальные рефлексы блокируются тормозным процессом и тем самым создают благоприятные условия для беспрепятственного осуществления «победившего» рефлекса. Др. примером такой координации рефлексов может служить и т. н. Реципрокная иннервация антагонистических мышц конечностей, дыхательных и некоторых др. систем. Как было показано русскими учёными П. А. Спиро и Н. Е. Введенским (См. Введенский), Шеррингтоном и др., рефлекторное возбуждение мотонейронов сгибательных мышц конечностей закономерно сопровождается торможением мотонейронов их анатомических антагонистов — разгибательных мышц и наоборот. Антагонистические взаимоотношения могут выявляться также рефлексами, относящимися к разным функциональным системам; например, при глотательном акте кратковременно задерживаются дыхательные движения. Важнейшая роль торможения в координационной деятельности Ц. н. с. наглядно прослеживается при введении в организм подопытного животного определённой дозы стрихнина, временно лишающего Ц. н. с, способности развивать торможение. У такого животного практически исчезает всякая координация рефлексов: любое локальное раздражение вызывает одновременную двигательную реакцию всей мускулатуры тела.Исследованиями Сеченова и голландского физиолога Р. Магнуса было выявлено значение исходного функционального состояния Ц. н. с. для её координационной деятельности. Раздражение определённого рецептивного поля задней конечности животного с перерезанным спинным мозгом вызывает противоположные эффекты в зависимости от исходного состояния раздражаемой конечности: если конечность находится в разогнутом состоянии, то раздражение вызывает сгибательный рефлекс, если же она согнута, то раздражение вызывает разгибательный рефлекс. Исходное положение конечности через соответствующие кожные и мышечные нервные пути отражается в центральном органе; происходит изменение его функционального состояния. Рецепторы, заложенные в исполнительных органах, особенно в двигательном аппарате, информируют адекватные структуры Ц. н. с. не только об исходном состоянии исполнительного органа в покое, но и о характере, интенсивности, продолжительности и всей динамике выполняемой им работы. Непрерывный поток информации, поступающей от собственных рецепторов исполнительных органов в процессе их работы, играет важную роль в коррекции и саморегуляции работы в соответствии с текущими потребностями организма и наличной обстановкой. Этот давно известный в физиологии Ц. н. с. важный принцип рефлекторной саморегуляции функций организма был в последующем определён как один из основных принципов кибернетики под название обратной связи (См. Обратная связь).Своеобразное кольцевое взаимодействие между центральным и периферическим звеньями рефлекторной дуги проявляется иногда в форме взаимного активирования, создания как бы кольцевого вращения возбуждения внутри элементов дуги, порождения особого типа цепи рефлексов: рефлекторное сокращение мышцы стимулирует заложенные в ней рецепторы, это вызывает новое рефлекторное сокращение той же мышцы и т.д. (Ч. Белл, А. Ф. Самойлов). Подобное кольцевое взаимодействие существует и между нейронами центрального звена рефлекторной дуги и проявляется в нескольких формах. Примером может служить установленный немецким учёным Б. Реншоу феномен. Аксон двигательного нейрона спинного мозга направляет в спинной мозг возвратную ветвь, которая через посредство тормозного интернейрона вступает в контакт с тем же двигательным нейроном и оказывает на него тормозящее влияние в случае чрезмерного интенсивного его возбуждения. Подобное явление «отрицательной обратной связи» описано применительно к пирамидным нервным клеткам коры больших полушарий головного мозга и к клеткам др. образований Ц. н. с. Примером же «положительной обратной связи» может служить описанное американским учёным Р. Лоренте де Но возрастающее усиление нервного возбуждения благодаря длительной его циркуляции по многоветвистому замкнутому кольцу нейронов рефлекторного центра: возвратная ветвь аксона одного из этих нейронов контактирует с одним или несколькими интернейронами, которые, контактируя с исходным нейроном, замыкают круг и создают структурную основу для многократной циркуляции возбуждения и своеобразного «самоусиления».Содружественные и антагонистические взаимодействия рефлексов неразрывно связаны между собой и по существу представляют собой разные стороны единого координационного процесса. При наличии прирождённого «общего конечного пути» содружественные рефлексы суммируются и взаимно усиливают друг друга, а антагонистические ведут борьбу за этот путь, и «победивший» рефлекс на определённый период времени затормаживает своих конкурентов. В некоторых случаях, например под воздействием внешнего или внутреннего фактора (локального механического давления, гормона, специфического яда, поляризующего тока и т.п.), значительно и стойко повышается возбудимость центрального аппарата какого-нибудь рефлекса и этот аппарат временно становится своеобразным магистральным «общим конечным путём»: стимуляция рецептивных полей разнородных рефлексов, вместо адекватных им специфических рефлексов, вызывает или усиливает рефлекс, возбудимость центра которого была повышена предварительно. Эта форма координации рефлексов была выявлена и изучена в нескольких модификациях, известных в современной нейрофизиологии под название проторения путей, суммационного рефлекса, облегчения, доминанты (См. Доминанта) и т.п. Считается, что нервный очаг в состоянии повышенной возбудимости или возбуждения оказывает тормозящее влияние на определённые элементы центральных структур разнородных рефлексов, отклоняет идущее по начальным звеньям их дуги возбуждение от обычного маршрута, как бы притягивает к себе, суммирует с собственным возбуждением и порождает или усиливает рефлекс, важный для организма на данном этапе. Координация рефлексов зависит также от функционального состояния их центрального аппарата; например, снижение его возбудимости при утомлении, действии отдельных нейро-гуморальных факторов и специфических ядов ведёт к тому, что даже доминировавшие раньше рефлексы легко уступают «общий конечный путь» др. рефлексам. Т. о., в основе многообразных форм координации рефлексов, как и координационной деятельности Ц. н. с. вообще, лежат активные по своей природе и противоположные по своим эффектам нервные процессы — возбуждение и торможение, различные комбинации и вариации распределения их в сложнейших макро- и микроструктурных элементах Ц. н. с., сложная динамика, взаимное проникновение, борьба и взаимодействие этих процессов.Субординация, специализация и локализация функций. Для деятельности Ц. н. с. характерна функциональная субординация, т. е. определённая соподчинённость между отдельными её органами, возникшая в процессе длительной эволюции. Неоднородность структуры и неравнозначность функций в Ц. н. с., а также соподчинённость между её частями намечаются на довольно ранних стадиях исторического развития организмов. Центральные нервные образования и рецепторы головы развиваются быстрее центральных нервных образований и рецепторов др. частей тела. Развитие ведущего отдела Ц. н. с. происходит не только по линии увеличения его массы, непрерывной дифференциации структуры и специализации функций уже сформировавшихся его отделов, но и по линии развития в нём всё новых центральных образований с рефлекторной деятельностью более высокого уровня и более важного значения, возглавляющих и регулирующих деятельность всех лежащих ниже их частей Ц. н. с. Этот процесс совершенствования, специализации, локализации и субординации функций Ц. н. с. достигает наиболее высокого уровня у высших млекопитающих животных и особенно у человекообразных обезьян и человека.Ц. н. с. находится в состоянии тонуса или тонического возбуждения даже при отсутствии каких-либо видимых внешних признаков её активности. Тоническое возбуждение Ц. н. с. — проявление общей рабочей готовности и благоприятный функциональный фон для возникновения и протекания активной деятельности. В тоническом возбуждении Ц. н. с. наглядно обнаруживается субординация между её образованиями, особенно в продолговатом, среднем и промежуточном мозге. Например, у высших животных хирургическая перерезка или холодовая блокада путей между продолговатым и спинным мозгом, т. е. прекращение субординационного влияния первого на второй, влечёт за собой развитие спинального шока — глубокое и продолжительное угнетение рефлексов спинного мозга. Последствием подобной перерезки на уровне передних бугров четверохолмия бывает т. н. Децеребрационная ригидность — сильное тоническое сокращение всех разгибательных мышц и потеря животным способности активно принимать нормальную позу или поддерживать её.Ещё более ярко субординация между отделами Ц. н. с. проявляется в процессе её активной деятельности. При этом каждое вышележащее звено Ц. н. с. осуществляет более сложные по структуре и составу рефлексы, более совершенную их интеграцию с вовлечением также рефлексов, регулируемых нижележащими звеньями. Особенности рефлекторной деятельности основных «этажей» Ц. н. с. можно представить в следующем виде: рефлексы сегментов спинного мозга охватывают лишь отдельные органы (например, отдельные конечности); более совершенные рефлексы продолговатого мозга распространяются на деятельность отдельных систем органов (пищеварительной, дыхательной, сердечно-сосудистой, двигательного аппарата и т.д.); рефлексы среднего мозга охватывают всю скелетную мускулатуру тела и обеспечивают организацию таких сложных двигательных функций, как стояние и ходьба; образования промежуточного мозга рефлекторно регулируют и координируют деятельность внутренних органов всех систем организма во всевозможных сочетаниях в связи с осуществляемыми ими жизненно важными безусловными рефлексами — пищедобывательными, защитными, половыми и т.п. Большие полушария головного мозга способны не только совершенствовать все эти рефлексы и объединять их в более сложные комплексы, но и создавать качественно новые виды рефлексов — условные рефлексы. При этом чем выше уровень развития животного и уровень организации его Ц. н. с., тем сильнее «власть» высших её отделов над нижележащими, тем значительнее их участие в регулировании многообразных функций организма и управлении ими. Усиление в процессе эволюции значения высших отделов Ц. н. с. в многообразной жизнедеятельности организма определяется как церебрализация, энцефализация, или кортикализация, функций организма (см. Кортико-висцеральные отношения). Однако и нижележащие отделы Ц. н. с. влияют на вышележащие; кроме того, снизу вверх поэтапно передаётся также вся информация от наружных и внутренних органов. Потому субординацию в Ц. н. с. следует рассматривать лишь как выражение превалирующего направления в сложном и многообразном взаимодействии между нервными образованиями разных уровней. Например, ретикулярная формация оказывает сильное активирующее и тормозящее воздействие на функциональное состояние почти всех частей Ц. н. с., в том числе и на кору больших полушарий головного мозга, которая в свою очередь влияет на функциональное состояние и деятельность ретикулярной формации и др. глубинных образований мозга, в том числе и на передаточные узлы восходящих трактов, регулируя тем самым поток получаемой по ним информации. Кольцевое взаимодействие между образованиями Ц. н. с. и саморегуляция их функций подтверждают правильность положения Павлова о ведущей роли коры больших полушарий головного мозга в объединённой и целостной деятельности всей Ц. н. с.Структурные и функциональные особенности Ц. н. с. обеспечивают многообразие и совершенство её деятельности в соответствии с потребностями организма, возникновение в случае необходимости новых форм координации. Богатый фонд «запасных возможностей» и компенсаторных приспособлений Ц. н. с. имеет важное биологическое значение как в условиях нормального существования организма, так и при всякого рода повреждениях периферических воспринимающих и исполнительных органов, приводящих и отводящих нервных структур и различных органов самой Ц. н. с. О патологии Ц. н. с. см. Нервные болезни, Психические болезни, Череп.Лит.: Прохаска Г., Физиология, или наука о естестве человеческом, пер. с нем., СПБ. 1822; Орбели Л. А., Лекции по физиологии нервной системы, 3 изд., М. — Л., 1938; Декарт Р., Избр. произв., пер. с франц. и лат., М., 1950; Ухтомский А. А., Собр. соч., т. 1, Л., 1950; Павлов И. П., Полное собрание соч., 2 изд., т. 1—6, М. — Л., 1951—52; Введенский Н. Е., Полное собрание соч., т. 4, Л., 1953; Сеченов И. М., Избр. произв., т. 1—2, М., 1952—56; Ройтбак А. И., Биоэлектрические явления в коре больших полушарий, ч. 1, Тб., 1955; Магнус Р., Установка тела, пер. с нем., М. — Л., 1962; Мэгун Г., Бодрствующий мозг, пер. с англ., 2 изд., М., 1965; Беритов И. О., Общая физиология мышечной и нервной системы, 3 изд., т. 2, М., 1966; Самойлов А. Ф., Избр. труды, М., 1967; Анохин П. К., Биология и нейрофизиология условного рефлекса, М., 1968; Русинов В. С., Доминанта. Электрофизиологические исследования, М., 1969; Асратян Э. А., Очерки по физиологии условных рефлексов, М., 1970; Физиология высшей нервной деятельности, ч. 1—2, М., 1970—71; Костюк П. Г., физиология центральной нервной системы, К., 1977; Ливанов М. Н., Пространственная организация процессов головного мозга, М., 1972; Рабинович М. Я., Замыкательная функция мозга, М., 1975.Э. А. Асратян.
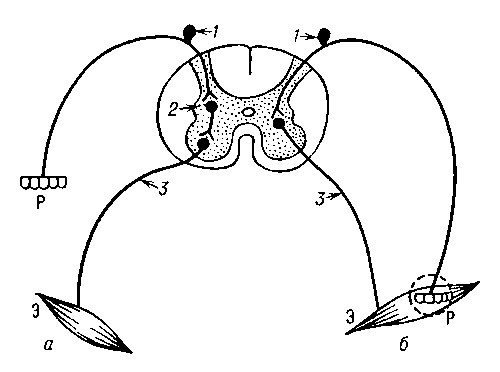 Рис. 3. Схема дивергенции (а) и конвергенции (б) нервных путей в спинном мозге: 1 — афферентный нейрон; 2 — интернейрон; 3 — эфферентный нейрон.
Рис. 3. Схема дивергенции (а) и конвергенции (б) нервных путей в спинном мозге: 1 — афферентный нейрон; 2 — интернейрон; 3 — эфферентный нейрон.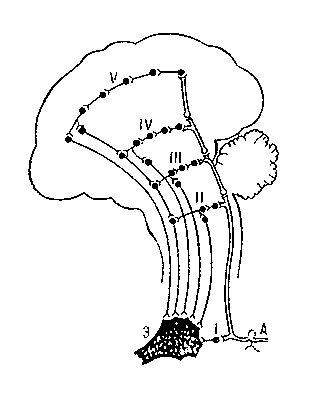 Рис. 2. Схема рефлекторной дуги с многоэтажной центральной частью: А — афферентный нейрон, Э — эфферентный нейрон; I—V уровни ветвей центральной части дуги.
Рис. 2. Схема рефлекторной дуги с многоэтажной центральной частью: А — афферентный нейрон, Э — эфферентный нейрон; I—V уровни ветвей центральной части дуги.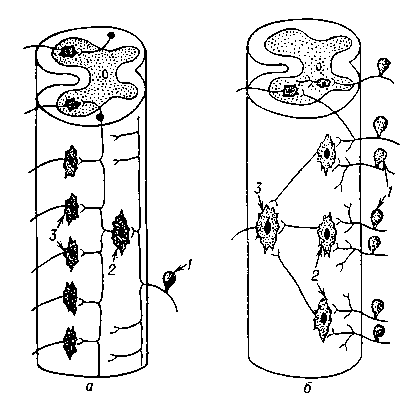 Рис. 1. Схема рефлекторной дуги спинального рефлекса: а — трёхнейронная дуга; б — двухнейронная дуга; Р — рецептор; Э — эффекторный орган; 1 — афферентный нейрон; 2 — вставочный нейрон; 3 — эфферентный нейрон.
Рис. 1. Схема рефлекторной дуги спинального рефлекса: а — трёхнейронная дуга; б — двухнейронная дуга; Р — рецептор; Э — эффекторный орган; 1 — афферентный нейрон; 2 — вставочный нейрон; 3 — эфферентный нейрон.
Большая советская энциклопедия. — М.: Советская энциклопедия. 1969—1978.